
Abstract
Mahanarva andigena was detected in Colombia for the first time in 1999, in the municipality of Tumaco (Nariño), on the south Pacific coast, associated with the hosts Sorghum halepense and Saccharum officinarum. In order to complement studies aimed at an understanding of the biology of other species of spittlebugs associated with cultivated grasses in Colombia, this research was designed to establish an initial base on the morphological differentiation, duration of the developmental stages, and certain aspects of the reproductive biology. The morphological studies permitted the differentiation and characterization of the egg, nymph and adult, at the same time determining the duration of the life cycle under semi-controlled conditions. For the eggs (27°C, 100% R. H.), the mean time to eclosion was 16.4 days. For the nymphs (mean temperature min/max 19.5/29.9°C, 95% R. H.) the mean development time was 48.4 days, divided in 6.4, 8.6, 8.2, 10.1 and 15.1 days for the five instars, respectively. For the adults (mean temperature min/max 19.5/29.9°C, 95% R. H.), the mean longevity was 21.4 days, 25.6 days for the females and 18.3 days for the males. Under the conditions of the study, the total duration of the life cycle of M. andigena was 75.5 days. M. andigena exhibited a flexibility for oviposition substrate, with a preference for uncovered soil with 51.2% of the recovered eggs; 16.4, 32.4 and 0% were recovered from soil under litter, surface of the stem, and the litter, respectively. The information provided strengthens certain patterns established for the spittlebug complex in grasses and broadens our understanding of the biological variation. As the first study on a spittlebug species associated with sugar cane in Colombia, a base line is established for biological studies of spittlebugs as emerging pests of the sugar cane industry.
Introducción
El salivazo de los pastos (Hemiptera: Cercopidae) es una plaga limitante en la producción ganadera de América tropical (Lapointe y Miles 1992; Holmann y Peck 2002). Ataca a casi todos los géneros importantes de gramíneas forrajeras (p. е. Bothriochloa, Brachiaria, Cynodon, Panicum, Pennisetum) (Peck 2001), al igual que la caña de azúcar (Fewkes 1969; Guagliumi 1972; Dinardo-Miranda et al. 2004) y bajo ciertas circunstancias arroz y césped (Nilakhe 1985; Shortman et al. 2002). A pesar del amplio rango de hospederos reportados para el salivazo de los pastos y del impacto económico que genera en la ganadería, todavía no se tiene una estrategia efectiva de manejo integrado de plagas (MIP). La diversidad taxonómica del grupo implica que la biología y las tácticas de manejo apropiadas varían de una región a otra, entonces el manejo de la plaga es afectado por una simplificación del problema dado el rango de variación en las asociaciones salivazo/forraje/hábitat.
En 1999 se reportó por primera vez para Colombia la especie Mahanarva andigena (Jacobi), de la cual solo se tenía referencia de su ubicación en la costa y amazonía de Ecuador (Peck 2001). En Colombia, M. andigena se reporta hasta la fecha solamente en la costa Pacífica, específicamente en Tumaco (Departamento de Nariño) en el Centro de Investigación El Mira de Corpoica (1° 33′ 101” N, 78° 42' 849″ W, 50 msnm) (Peck 2001). Además, se pudo establecer como hospederos de M. andigena a Sorghum halepense (L.) Pers. (pasto Johnson) y Saccharum officinarum L. (caña de azúcar). En el Ecuador, se reporta como hospedero de M. andigena a la caña de azúcar, donde los primeros reportes se remontan a 1986 en el Ingenio San Carlos en el Valle de Guayas (Mendoza 1999). Adicionalmente, Fors (2000) reporta para el ingenio San Carlos un área de 100 ha afectada por el insecto, al igual que una nueva zona con incidencia del insecto en inmediaciones del ingenio Valdéz cerca de Guayaquil (Ecuador).
En Ecuador las variedades de caña con mayor incidencia del insecto son aquellas que exhiben dificultad para el deshoje (Mendoza 1999). Para el caso específico de Colombia, M. andigena se encontró asociada con caña panelera, logrando una alta incidencia hasta 25-35 ninfas por cogollo (Rodríguez observación personal). Al igual que en el Ecuador, esta especie de salivazo en Colombia exhibe un comportamiento aéreo siendo común poder observar los diferentes instares ninfales en una sola masa de espuma localizada dentro del cogollo de la planta. Este comportamiento permite diferenciar a M. andigena de las demás especies reportadas para Colombia, y la compara con Mahanarva bipars (Walker) (Peck et al. 2004).
En este estudio se describe la biología y comportamiento de M. andigena bajo condiciones de casa de malla. A pesar de su importancia en Ecuador, no se cuenta con estudios detallados sobre la biología de este insecto. Este conocimiento es necesario para implementar tácticas de manejo tendientes a disminuir la presión que M. andigena pueda llegar a ejercer sobre la caña de azúcar, y como punto de partida para el desarrollo de estudios biológicos por parte de otras instituciones, por ejemplo en el caso de M. bipars asociada a caña en el Departamento de Risaralda (Peck et al. 2004).
Materiales y Métodos
Con el propósito de contar con todos los estados de desarrollo del insecto, se implementó una colonia en las instalaciones del Centro Internacional de Agricultura Tropical (CIAT), Palmira, Valle del Cauca, localizado a 3.517°N y 76.367°W, 965 msnm, precipitación anual 1.000 mm, y temperatura promedia de 24°C. Para el caso especifico de M. andigena, la metodología de cría implementada en otras especies de salivazo (Rodríguez et al. 2002) fue modificada en algunos aspectos según el comportamiento del insecto en campo y las condiciones climáticas del lugar de origen. Primero, se usaron nebulizadores en la casa de malla para facilitar el normal desarrollo de las ninfas, teniendo en cuenta que en el lugar de origen la humedad relativa (H. R.) es del 88% (Fig. 1). Segundo, se usaron jaulas de multiplicación acordes con el comportamiento aéreo de las ninfas. En cada jaula se dispusieron 20 materas plantadas con Brachiaria ruziziensis Germain y Everard (CIAT 654) como sustrato de alimentación para los adultos y posteriormente las ninfas. Cada jaula se infestó con un número adecuado de adultos asegurando no deteriorar la calidad del alimento y poder obtener así un número considerable de ninfas para el mantenimiento de la colonia y para los ensayos planteados durante la investigación (Fig. 1).

Montaje para el establecimiento de la colonia de M. andigena en el CIAT. A. Nebulizadores para asegurar humedad adecuada para desarrollo de las ninfas. B. Jaula de multiplicación para posterior cosecha de las ninfas para desarrollo de los ensayos.
Se realizaron estudios de caracterización biológica de M. andigena empleando metodologías establecidas y evaluadas anteriormente en otras especies de salivazo presentes en Colombia por el CIAT (Peck et al. 2002; Rodríguez et al. 2002; Rodríguez et al. 2003). Esto incluyó tres aspectos fundamentales: reconocimiento de los estados de vida mediante caracterización morfológica, cuantificación del ciclo de vida, y descripción de los sitios de oviposición como un aspecto de la biología reproductiva. Para reconocer los estados de vida, se caracterizaron ciertos aspectos morfológicos externos de las cuatro fases de desarrollo de los huevos, cinco instares ninfales, y ambos sexos en los adultos. Eso se realizó con la ayuda de un microscopio estereoscopio acondicionado con una reglilla ocular. Los adultos se coleccionaron en campo, las ninfas se obtuvieron en la colonia, y los huevos se obtuvieron de hembras colectadas en la colonia.
Para establecer el ciclo de vida, se cuantificó la duración de cada uno de los estados de desarrollo. Para la fase de huevo, las observaciones se llevaron acabo bajo condiciones controladas de incubación (27°C, 100% H. R. y oscuridad total). Los huevos (< 24 horas de edad) fueron mantenidos sobre papel filtro húmedo y mediante observación diaria se estableció el paso de una fase a otra. La duración de cada una de las cuatro fases generalizadas de desarrollo se determinó basado en 100 individuos. Para los estados ninfal y adulto, las actividades se realizaron en condiciones de casa de malla. Se emplearon ninfas de primer instar recién eclosionadas (< 12 horas de edad) que fueron trasladadas a potes individuales con B. ruziziensis como sustrato de alimentación. Cada pote fue infestado con una sola ninfa, facilitando de esta manera determinar el paso de un instar a otro por observación directa o por presencia de la exuvia. Para el caso de los adultos, se emplearon tenerales (< 12 horas de edad) obtenidos en la colonia, confinados en potes plantados con B. ruziziensis y cubiertos con un cilindro de acetato. Cada pote fue infestado con cinco adultos no sexados. Mediante observación diaria se estableció la longevidad media para los adultos y se determinó el sexo.
Para establecer preferencias por sitios de oviposición, se replicaron condiciones de campo en casa de malla, teniendo en cuenta cuatro sustratos de oviposición: suelo descubierto, suelo cubierto con hojarasca, hojarasca, y la superficie de la planta (Rodríguez et al. 2002). Para esto se emplearon diez potes cada uno con dos hembras y dos machos tenerales de la colonia. Diez días después de su montaje se procedió a recuperar los huevos de cada uno de los sustratos. Para la recuperación de los huevos del suelo descubierto y cubierto por hojarasca se empleó el método de Lapointe et al. (1989). Para el caso de la hojarasca y la superficie de la planta la observación fue directamente sobre el sustrato (Rodríguez 2001).
La información registrada fue agrupada en valores mínimos, máximos y promedios acompañados del error estándar como medida de variación. Los análisis estadísticos se realizaron con el programa SAS 8.1 y la prueba de comparación múltiple Tukey-Kramer HSD al P > 0,05. En el caso de la longevidad de los adultos, se empleó la distribución de Weibull (Pinder et al. 1978).
Resultados
Los huevos de M. andigena pasaron por cuatro fases de desarrollo plenamente diferenciables por características morfológicas externas que las acompañan. En la fase S1, los huevos presentaron una tonalidad crema, sin ninguna característica externa. La fase S2 se caracterizó por la aparición de una mancha roja ubicada debajo de la sutura de eclosión. Para la fase S3 se estableció la apertura de la sutura de eclosión como característica externa. La fase S4 exhibió como característica externa adicional la presencia de las manchas oculares y abdominales. Durante su desarrollo, los huevos de M. andigena presentaron un incremento en el largo y ancho a medida que pasaban de una fase a otra, con un 6,2% de incremento en tamaño entre la fase S1 y S4 para largo y ancho (Tabla 1). Los huevos de M. andigena exhibieron una duración total de 16,4 días con una duración mayor para la fase S4 con el 39,3% de la duración total (Tabla 2).
Largo y ancho (mm) para las fases de desarrollo de los huevos de M. andigena (promedio ± E.E., rango, n = 93-100).
En cada fila, promedios seguidos de letras diferentes son estadísticamente diferentes (P < 0,05).
Duración (días) de las fases de desarrollo de los huevos de M. andigena (n = 96-105).
En cada fila, promedios seguidos de letras diferentes son estadísticamente diferentes (P < 0,05).
Las ninfas de M. andigena presentaron un incremento para cada uno de los parámetros morfológicos evaluados al pasar de un instar a otro (Tabla 3). Se pudo establecer además una disminución para ambos sexos del largo del estilete al pasar del instar Vb (próximo a transformarse en adulto, evidenciado por la pesencia de espinas en la metatibia) al estado adulto (P < 0,05). De los parámetros morfológicos evaluados, el ancho de la cápsula cefálica exhibió la mayor diferencia entre instares (no había traslape), siendo posible la utilización de este carácter como herramienta para diferenciar entre instares (Fig. 2). La duración del estado ninfal fue de 48,4 días, con un 32,4% representado por la duración del V instar (Tabla 4). Las ninfas de M. andigena exhibieron una tonalidad amarillenta característica durante el primer instar; a medida que se desarrollaron adquirieron una tonalidad verdusca en el abdomen.
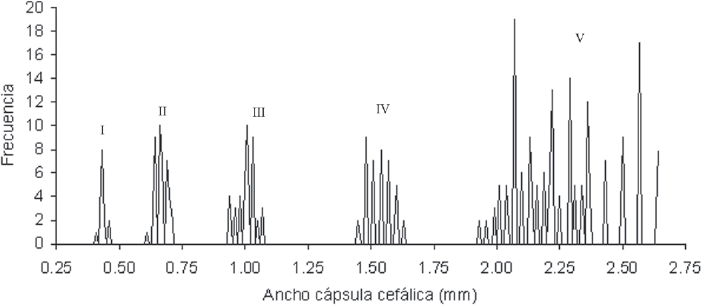
Frecuencia del ancho cápsula cefálica para las ninfas de M. andigena.
Caracterización morfológica de ninfas de M. andigena (promedio ± E.E., rango, n = 15-40).
En cada columna, promedios seguidos de letras diferentes son estadísticamente diferentes (P < 0,05).
Se evidencian espinas en metatibia del adulto. Todas las medidas en mm.
Duración en días de los estados ninfales de M. andigena (n = 40).
Promedios seguidos de letras diferentes son estadísticamente diferentes (P < 0,05).
Los adultos de M. andigena exhibieron un dimorfismo sexual expresado por el mayor tamaño de las hembras en cada uno de los caracteres morfológicos evaluados (Tabla 5). La relación macho-hembra fue de 1:1. El parámetro morfológico que exhibió mayor diferencia entre los sexos fue el largo del cuerpo siendo 10,9% más grande en las hembras. Los adultos de M. andigena exhibieron una longevidad media de 21,4 días, con una longevidad de 25,6 y 18,3 días para hembras y machos, respectivamente. Para las condiciones del estudio, se estableció una duración del ciclo de vida de M. andigena de 75,5 días aproximadamente (16,4+48,4+10,7, huevo+ninfa+1/2 adulto).
Caracterización morfológica (mm) de los adultos de M. andigena por sexo (promedio ± E.E., rango, n = 40).
En cada columna, promedios seguidos de letras diferentes son estadísticamente diferentes (P < 0,05).
Con respecto a la preferencia por sustrato de oviposición, M. andigena exhibió una flexibilidad por el sustrato con una preferencia por el suelo descubierto donde el 51,2% de los huevos fueron recuperados. El 16,4 y 32,4% de los huevos fueron recuperados en el suelo bajo hojarasca y superficie de la planta, respectivamente. No se recuperaron huevos de la hojarasca. Adicionalmente, se determinó una fertilidad del 93,3% para las hembras de M. andigena bajo las condiciones del estudio, con una mortalidad del 6,7% de huevo-ninfa (Fig. 3).
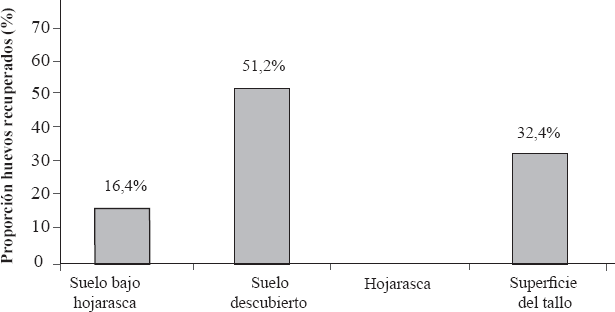
Porcentaje de huevos recuperados para M. andigena en cada uno de los sustratos evaluados en condiciones de casa de malla.
Dentro del comportamiento observado para M. andigena durante el ensayo, se resalta la preferencia de las ninfas por la parte aérea (axilas de las hojas y/o cogollos) para establecerse y continuar su desarrollo. Además, hubo un comportamiento agregado expresado en la facilidad de encontrar dentro de una masa de espuma diferentes instares ninfales.
Discusión
Se pudo establecer la eficacia de la metodología usada para el estudio de M. andigena, teniendo en cuenta algunas modificaciones producto del comportamiento diferencial de esta especie con respecto a otras especies estudiadas anteriormente por el CIAT. Al igual que otras especies de salivazo, M. andigena presenta dimorfismo sexual expresado por el mayor tamaño de las hembras, los instares ninfales pueden diferenciarse por ciertas características morfológicas externas, y los huevos incrementan su tamaño con el desarrollo. El ancho de la cápsula cefálica, permitió identificar los cincos instares observados para M. andigena al ajustarse a la ley de Dyar, presentando una progresión geométrica regular de 1,5 veces en cada muda (Cole 1980; Magalhães et al. 1987; Francisco y Prado 2001; Hernández et al. 2005).
El comportamiento de las ninfas en las condiciones de cría especificadas difiere del observado en otras especies, pero no de las observaciones realizadas en campo donde éstas se encuentran en la parte aérea de la planta de los hospederos reportados (Rodríguez observación personal). El color característico exhibido por las ninfas durante su desarrollo facilita diferenciarlas de las ninfas de otras especies estudiadas para Colombia (Peck et al. 2002; Rodríguez et al. 2002; Rodríguez et al. 2003).
La duración del ciclo de vida de M. andigena (75,5 días) es superior al de las otras ocho especies estudiadas en Colombia de los géneros Aeneolamia (45,3-52,6 días), Mahanarva (64,6 días), Prosapia (72,5 días) y Zulia (61,5-69,6 días) (CIAT 1998; Ballesteros y Gallego 1999; Peck et al. 2002; Rodríguez et al. 2002; Rodríguez et al. 2003). M. andigena es la especie colombiana que hasta la fecha exhibe la mayor duración para el estado ninfal, superando 1,9 veces la duración de Aeneolamia reducta (Lallemand) (Peck et al. 2002). De igual forma, el estado adulto de M. andigena es 3,0 veces más prolongado que lo establecido para Aeneolamia varia (Fabricius). M. andigena al igual que P. simulans, son las dos únicas especies estudiadas hasta ahora en Colombia cuyo ciclo de vida supera los 70 días (Rodríguez et al. 2002; Rodríguez et al. 2003). La duración del ciclo de vida de M. andigena difiere de lo reportado para otras especies del género en la literatura, M. posticita y M. fimbriolata con 79,1 y 60,4 días (García 2002; García et al. 2006). La preferencia por el suelo descubierto (51,2%) como sustrato de oviposición es similar al comportamiento de otras especies estudiadas (A. lepidior, A. reducta, A. varia, Mahanarva trifissa (Walker), Zulia carbonaria (Lallemand) y Zulia sp. nov.) (CIAT 1998; Ballesteros y Gallego 1999; Peck et al. 2002; Rodríguez et al. 2002; Rodríguez et al. 2003). La flexibilidad de las hembras de M. andigena de ovipositar sobre la superficie de la planta (32,4%) es comparable con lo establecido para Zulia pubescens (Fabricius) y P. simulans con el 59,2 y 82,6% de los huevos recuperados en este sustrato, respectivamente (Rodríguez et al. 2002; Rodríguez et al. 2003).
La fertilidad de las hembras de M. andigena (94,2%) es comparable con la establecida bajo las mismas condiciones para P. simulans y Z. carbonaria 95,1 y 97,9%, respectivamente (CIAT 2002). La mortalidad de huevo-ninfa exhibida por M. andigena es igual a la establecida para P. simulans, pero 1,5 veces menor a la de Z. carbonaria bajo las mismas condiciones de estudio (CIAT 2002).
Conclusiones
El comportamiento aéreo registrado por las ninfas de M. andigena en campo y bajo las condiciones del estudio, las diferencia de las demás especies de salivazo hasta la fecha estudiadas en Colombia; convirtiéndose en una característica propia del género, aspecto que tiene implicaciones en las tácticas de manejo establecidas hasta el momento, tendientes a disminuir el impacto de esta plaga en campo.
Los resultados obtenidos amplían la variación biológica conocida de este grupo de insectos plaga de gramíneas en Colombia. Destacándose la flexibilidad que exhibe M. andigena por el sustrato de oviposición, lo que implica un manejo adecuado de los materiales de propagación vegetal para evitar la dispersión de esta especie en zonas del país donde aun no ha sido reportada.
Los estudios biológicos de M. andigena, constituyen los primeros avances en el conocimiento de esta especie de salivazo asociada a caña en Colombia. Esto permite establecer protocolos para estudios biológicos posteriores de otras especies de salivazo, que en su momento se puedan constituir en plagas potenciales de la caña en Colombia.
Footnotes
Agradecimientos
Se agradece al grupo de Bioecología y MIP del Salivazo del CIAT por la colaboración prestada para el desarrollo de este trabajo, de igual forma al personal del C. I. el MIRA del ICA, Tumaco, por facilitar los muestreos en esta zona del país. Este trabajo se realizó gracias al apoyo prestado por el Proyecto IP-5 Gramíneas y Leguminosas Tropicales, CIAT y ayuda financiera parcial del Programa Nacional de Transferencia de Tecnología Agropecuaria (PRONATTA).