
Abstract
The artificial diet called CENIBROCA was developed for culturing Hypothenemus hampei (Ferrari). The diet consists of casein, power yeast, industrial agar, and ground green coffee and some antimicrobial agents. It was suitable for rearing CBB through five generations in the laboratory. This generations were obtained without any significant differences in fecundity (α=0,05), but they were affected in fertility. The values of net maternity, oviposition percentage and flying activity obtained from first to fifth generation were compared with the values of CBB from the field (control), which were reared under the same laboratory condition. The fecundity was significantly affected, the highest value (35,6 ± SD=12,8) was obtained in CBB from the field, which was significantly different from the others (α < 0,05). The second highest value occurred in the first generation with 25,1 ± 12,7, followed for second, third, fourth and fifth generations with 23,6 ± SD= 14,3, 24,0 ± SD=11,3, 20,4 ± SD=15,9 and 21,2 ± SD=14,3, respectively (±=0,05). The lowest value of gross fertility was calculated for the fifth generation. The maximum value of net maternity was for CBB from the field.
Introduction
The coffee berry borer (CBB), Hypothenemus hampei (Ferrari), is the most notorious pest of coffee, feeding exclusively on coffee berries. Native to Central Africa, it has spread to most coffee producing countries in the word. Its feeding habits have caused substantial production losses, and the berries can be attacked at all stages of development.
There has been extensive knowledge on CBB life cycle under laboratory conditions, which has been an essential prerequisite to colony management. The most studies provide information on behavior and feeding, fertility, mortality and hatchability of eggs, stage development times, which have been determined under different conditions of temperature and relative humidity.
Much is already known about the culturing techniques of H. hampei (Benavides and Portilla, 1990; Barrera et al. 1991; Brun et al. 1993; Villacorta and Barrera 1993; Portilla and Bustillo 1995). In Colombia Benavides and Portilla (1991) reared H. hampei on different coffee substrates and reported for coffee berries an oviposition rate of 55,5% of berries with 74,8% contamination by fungus; for dry parchment coffee artificially moistened 78,3% oviposition and contamination of 21%; on wet parchment coffee (recently processed) 87,8% oviposition and 4,1% contamination, and finally on almond coffee (coffee without hull) 55,3% oviposition and 82,7% contamination. However, one of the problems with the methods previously mentioned is the dependence upon fruit collected from the field.
The first attempt to rear H. hampei using artificial diet was done by Bautista and Martinez in 1982, but their results were not successful (Villacorta 1989). In 1985 Villacorta developed a successful diet for rearing this insect. Villacorta and Barrera (1993) modified this diet, producing the Ecobrovill-160 artificial diet (1996). Brun et al. (1993) reported another artificial diet for H. hampei on which were reared 15 generations. In 1998 an ARS scientists at Weslaco, TX developed an artificial diet and her diet has been used to maintain a small culture of CBB for over 2 years (Rojas, unpublished). Portilla (1999a) modified and improved the Villacorta diet (Now called Cenibroca) and reduced the cost from $ 15/L to about $ 1.5 and she also develop a technique for rearing CBB in large-amount using a multi-cellular tray (Portilla 1999b).
The ingredients used for preparing Cenibroca diet, such as industrial agar, powder torula yeast and ground green coffee offer the required physical consistency of the medium for the external development of CBB stages. The amount of coffee and torula yeast increased the CBB brood size very significantly in relation to previous diets (Portilla 1999a).
Brun et al. (1993) and Rojas (unpublished) kept the borer for several generations, but they do not mention how is the CBB response to continuous rearing. Rojas (unpublished) worked with Ecobrovill-160 artificial diet and observed that the CBB reproductive potential declined through generations. Portilla (unpublished) found similar results in studies of continuous rearing of CBB using parchment coffee beans. Similar declines in fecundity occur to the CBB reared in the Cenicafe laboratories and by private producers, so they have to re-collect new stock from the field each generation, a very difficult and expensive option (Baker 1999). Thus, the normal reproduction of this beetle is affected even if it is developed on its own natural medium. However, the sex ratio (10 females: 1 male) was not affected (Portilla 1999c).
Thus, the objective of this study is to provide accurate information about the response of the CBB demographic parameters trough continuous rearing.
Materials and Methods
This study was carried out in the entomology laboratory of CENICAFE in Colombia. It was conducted in constant temperature environmental chambers at 27°C ± 1 and 85% of relative humidity. The relative humidity was controlled by saturated solutions of salts inside containers (26,5 g of KCI/100 ml of water) (Winston and Bates 1960).
Preparation of the diet
Dried green coffee beans (12% moisture content) were ground (2-mm particle size) using a Provat Emmerich miller it was autoclaved for 20 min at 120°C and 15 lb/m of pressure. The agar and water mixed was also sterilised using an autoclave. Group II ingredients were combined and partially mixed and tipped into the blender together with molten agar. The mixture was blended for 10 min to achieve complete mixing. When it had cooled to 50°C, the group III ingredients, dissolved in 10 ml of sterilised water, were added (Table 1).
After a final mixing the completed diet was poured into rectangular plastic boxes, 150 x 60 x 15 mm to a depth of 5 mm each (10 ml of diet/box). The diet was divided into 20 small pellets using a grille squeezed into it while still hot. Each pellet was 0,5 ml.
The rearing containers of solid diets were dried at 50°C for 24 hours in a heater until the diet attained a leathery texture (55% diet humidity). Afterwards, these containers of solid diet were infested with CBB.
Fecundity
Recently emerged adult females of CBB were used for infesting Cenibroca diet. These adults were collected from berries naturally infested in the field and kept for some weeks in the laboratory to allow pupae and larvae to become adults. The most active borers were selected and disinfected for infesting diet and were put into the box individually for each pellet. These plastic boxes with infested diet were placed inside the environmental chambers and kept for 30 days under darkness. The brood obtained from these initial CBB was considered as control generation.
Composition of the CENIBROCA, artificial diet for rearing coffee berry borer
Formaldehyde only is used when the diet was infested with CBB from the field.
At 30 days the most active females obtained from the control generation were transferred to fresh diet to begin a new cycle. Thus, the broods obtained in each generation were transferred to fresh diet every 30 days to get a new one for a total of 6 months. So, five generations were obtained from the control generation. From each generation 50 diet pellets randomly selected were picked from the rearing container every 5 days for 30 days. They were dissected and the contents recorded.
Activity of CBB
1800 CBB (10 groups of 30 CBB per generation) were randomly selected. These groups of CBB were evaluated one at a time in a lighted room 1 m wide x 2,5 m length x 2,5 m high at 27°C, 85% RH. Each group was placed into a transparent plastic box 35 x 25 x 12 cm, which was located in the center of the room. The length of the first flight was measure in minutes, the fraction of walking and non - walking borers were calculated by dividing the number of borers that walked and the number of non-walking borers by the total of sample size used per treatment (30 CBB per group).
Reproductive potential and rates of increase
Basics population parameters were calculated. From the number of eggs counted per sample, of total of eggs laid was calculated. Developmental stage mortality found in studies carried out by Portilla (1999c) (Temperature dependent development of CBB using Cenibroca artificial diet) was taken for estimating the survival at age x. From the numbers of eggs counted in the second sample (5 or 10 days after infestation) were subtracted the number of eggs counted in the previous sample, the difference was assumed as m. This parameter was estimated assuming a sex ratio of borer of 10 females: 1 male (Baker et al. 1994; Portilla 1999c).
Gross maternity (Mx), defined as the average number of female offspring in the interval x; survival (lx), designated as the proportion of a cohort surviving from birth to exact age x; fecundity (mx) represents the number of female eggs laid by a female at age x. The net maternity function, is the product of fecundity and survivorship (Carey 1993). That is:

The sum of lxmx over all age classes is defined as the net reproductive rate (Ro). It is the average number of female offspring that would be born to a birth cohort of females during their lifetime if they experience a fixed pattern of age-specific birth and death rates (Pressat 1985). This rate represents the factor by which a population will increase each generation. The formula for net reproductive rate is:
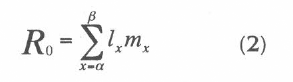
The factor by which a population will increase each day defined as finite rate of increase (λ), is related to the life table schedule in the equation (Krebs 1972):

The intrinsic rate of increase, rm is the rate of natural increase in a closed population that has been subject to constant age-specific schedule of fertility and mortality for a long period and has converged to be a stable population (Keyfitz 1964; Pressat 1985). This rate is basically a special case of a crude growth rate and it was determined by analytical approximation using the Lotka (1925) equation quoted by Carey (1993):
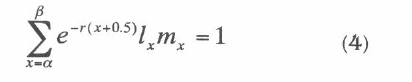
The mean generation time (T), is the mean age of reproduction, which characterises T as the mean interval separating the births of one generation from those of the next (Pressat 1985). The formula is:
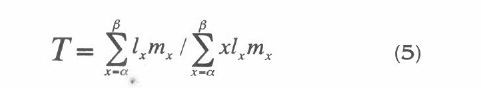
The doubling time (DT) was calculated using the following formula (Krebs 1972):

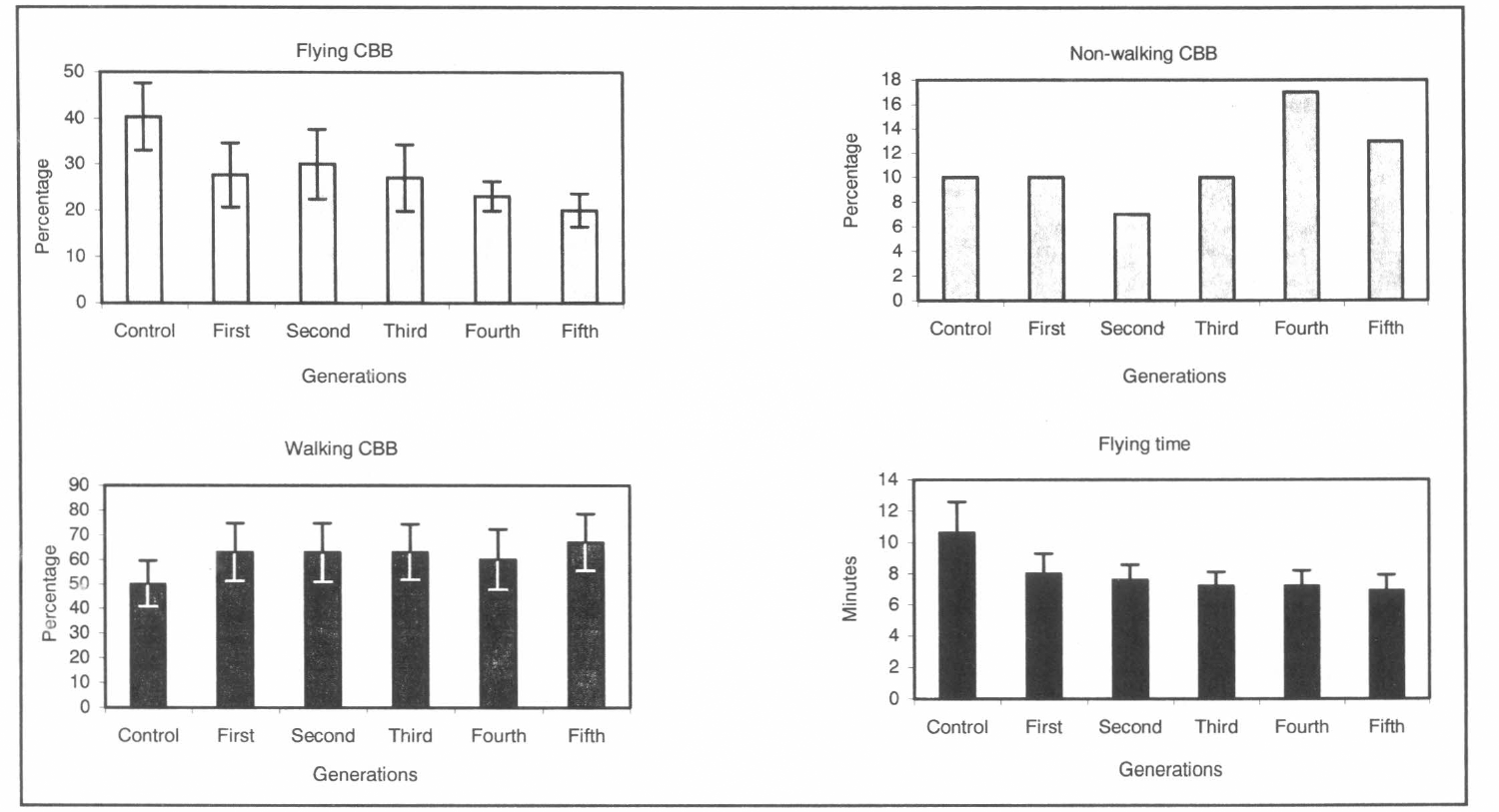
The parameter Mx is the average number of offspring produced by female age x, then the sum of this schedule over all age classes gives the lifetime production of offspring by an average female that lives to the last day of possible life in the cohort. The parameter hx is the average number of the hatched eggs produced by female age x. The counterpart of gross fecundity rate is gross fertility rate. Given by the formula
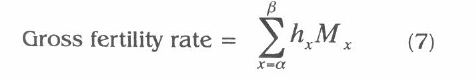
Which gives the total number of viable eggs an hypothetical cohort of females will produce if all live to the last age of reproduction. The ratio of gross fertility to gross fecundity gives the gross hatch rate (Carey 1993):
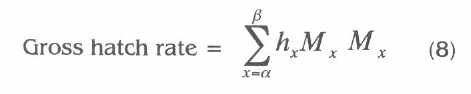
The general linear model (GLM) procedure of SAS (SAS institute 1988) was used to analyze differences in the mean number of stages per CBB found in 0.5 cc of diet at various times (5, 10, 15, 20, 25 and 30 days) after infestation between generations. The activity and percentage of oviposition were analyzed using the same statistical program.
Results
Reproductive potential
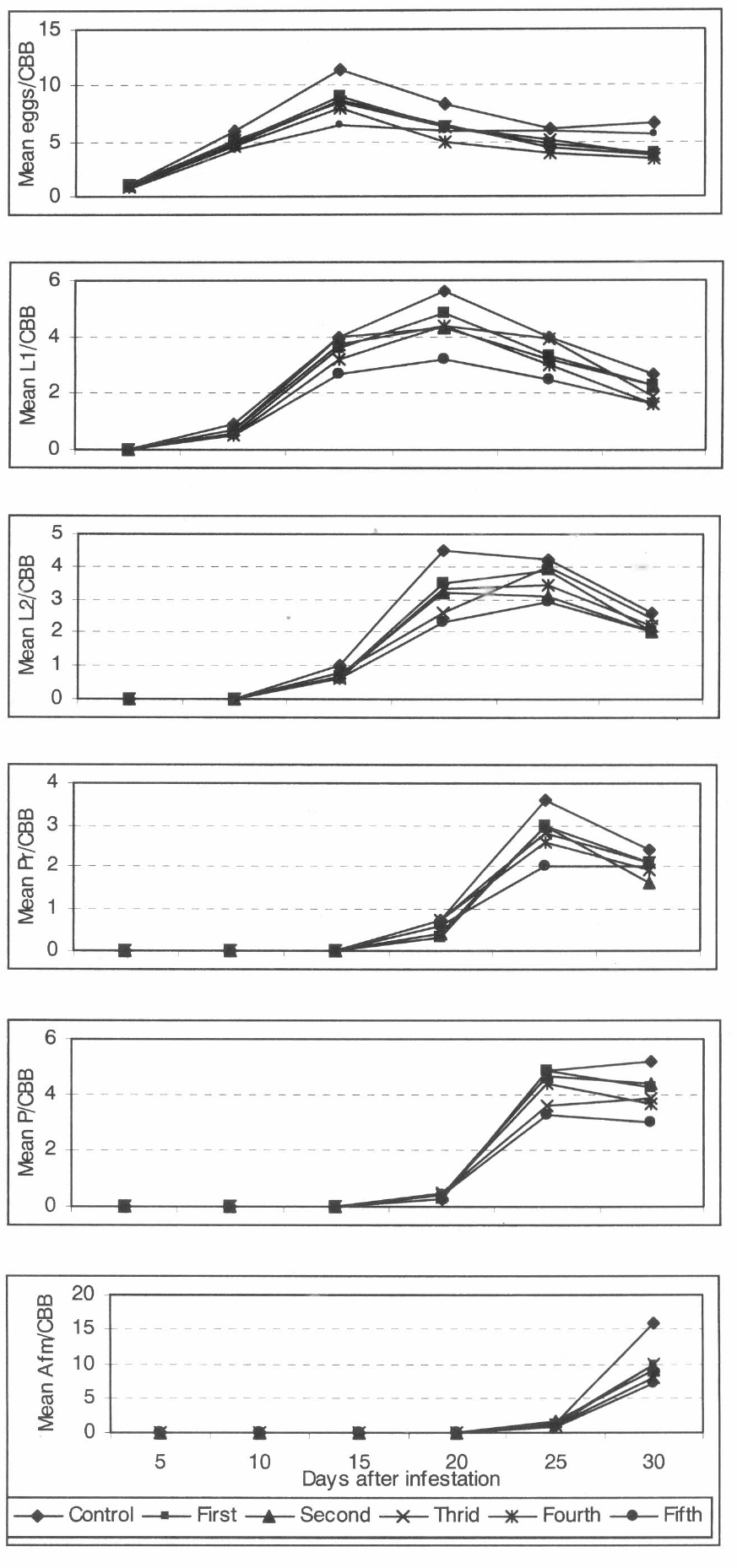
Table 2 gives the productivity of CCB in terms of the number of stages produced per beetle in 0,5 ml of diet obtained through five generations. GLM analysis showed that there were no significant differences among treatments at day 5. At day 30 the fecundity of CBB from the field was significantly different from all other treatments (α = 0,05) (F = 8,05, DF = 5, 294, p = 0,0001). These results clearly indicate that the fecundity was affected by continuous rearing.
Mean number of stage progeny produced per female per 0.5 cc in Cenibroca artificial diet through five generations
Means followed by the same letter in each row are not significantly different (α = 0,05 Tukey test). DAI = Days after infestation
Table 3 gives the values of 7 demographic parameters that were estimated based on various experiments. The growth rate of CBB from the field was 19,3 females per newborn female in a mean time of 45,4 days. Its rate decreased to 13 females per females in a mean time of 43 days for the first to the third generations. The growth rates for CBB fourth and fifth generations were 11 and 12 daughter females per female in a mean period of 43 and 44 days respectively. The population of CBB from the field could double in 10 days, while the generation from first to fourth generation could double in 11 days. The fifth generation could double its population in 12 days.
Life table statistic for coffee berry borer on Cenibroca artificial diet calculated for five generations
The distribution of developmental time of CBB is presented in figure 1. These distributions showed no variability in development time among generation, which were held under identical condition. Therefore, the presence of the first larval stage appeared at day 9, the second larval stage at 12 days, prepupae at day 16 and adult females and males at day 22. The distribution obtained in this study presents a noteworthy feature about the presence of each life stage of CBB, which appeared at the same time in every generation. This information can be incorporated into the harvest rate process in the rearing of H. hampei.
Mean number of stages per borer per 0.5 ml of Cenibroca artificial diet found at various times after infestation for five generations comparing with CBB brood from the field.
Fertility
The number of infested diet pellets with presence of unhatched eggs was considered for calculating gross hatch rate, and the ratio of total infested diet pellets and the number of diet pellets with progeny found at end of evaluation was taken to calculate the gross fertility. Figure 2b shows the percentage of each variable (gross fertility and gross hatch rate) obtained in this investigation and the mean numbers of stages per CBB per 0,5 cc of diet found at day 30 after infestation is give in the figure 2a.
Gross fecundity, gross fertility rate and gross hatch rate of CBB calculated for five generations using Cenibroca artificial diet (27°C, 85%RH, 55%DH). 95% confidence limits of the mean (sample size n = 50 CBB/treatment.
No berries with unhatched eggs were found in the CBB field treatment and during its first three generations. However, in the fourth and fifth generations 3 and 4 berries with unfertilized eggs were found respectively. The dividend of these diet pellets by the total sample size gave the fraction of unfertilized eggs (0,06 and 0,08, respectively), so the fraction hatch was 0,94 for fourth generation and 0,92 for fifth generation. The highest value of gross fertility rate was obtained in CBB from the field. The first, second and third generation had very little differences between their values, while the last two generations had the lowest percentages.
The results of these variables may have influenced in the decline of the reproductive potential of CBB (Fig. 2a) and hence in the values of all population parameters calculated previously.
Activity of CBB
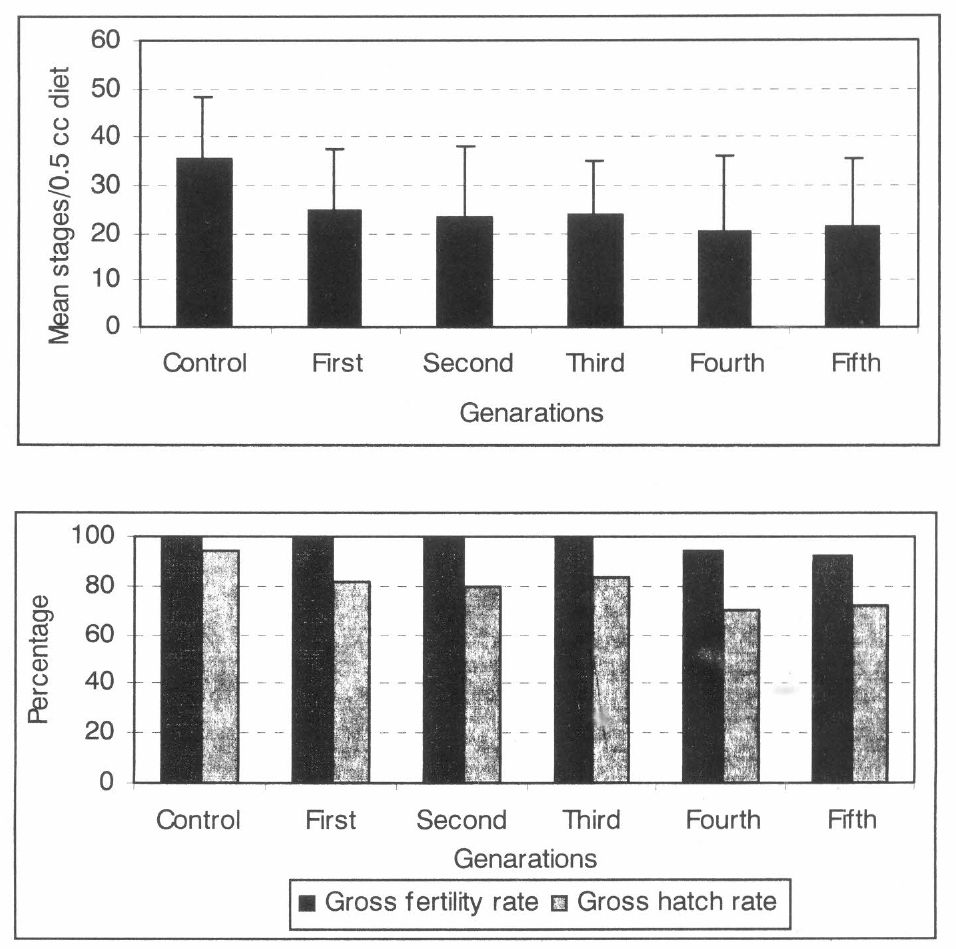
GLM analysis showed that there were no significant differences in average flight time, minimum flight time, and fraction walking However, there were significant differences in maximum flight time and the fraction of flying CBB. Due to the high variability within and between samples and the great number of zeros in the fraction of non-walking CBB, its statistical analysis was not carried out (Fig. 3).
Flight time and activity percentage of CBB evaluated for five generations. 95% confidence limits of the mean (sample size n=10 cohorts/treatment (30 CBB/cohort).
Flight of CBB females for periods up to 22 minutes, and in a succession of flights a total of three hours under laboratory condition has been recorded (Baker 1984). Although CBB were observed in a succession of flights and some times the subsequent flight was longer than the previous. In this experiment only the first continuous flight was considered. Thus, only 11 min was registered as average maximum flight time, which was observed for CBB from the field, and this was significantly different from the others treatments (α = 0,05) (F=3,92, DF=5, 54; p = 0,0001). Seven minutes was the average common time observed for all treatments.
GLM showed significant differences in the fraction of flying CBB. Field treatment was significantly different than first, second, third and fourth generations (α = 0,05) (F = 4,38, DF = 5, 54, P = 0,998) and high significantly different from the fifth generation. CBB from the field had the highest fraction of flying CBB (0,43) and the second lowest fraction of non-walking CBB (0,09), while the highest fraction of non-walking CBB (0,12) occurred in fourth generation. The results clearly indicate that in all generation the number of walking CBB was always highest than the number of flying CBB. Number of non-walking CBB (immobile) was very low in all cases.
Discussion
Control of relative humidity, diet moisture content and also texture of the diet are essential factors for improving the oviposition behaviour of the CBB and to reach a maximum reproductive potential of H. hampei (Portilla 1999c). Highest mean of adults produced per female depended largely on diets that kept mean diet moisture content not less than 40%. Initial diet moisture content of 55 to 60% exposed at 85% RH kept this percentage far longer. Microbial contamination and lowest fecundity occurred in diet with high moisture content at high relative humidities, while high mortality occurred in diets with low moisture content at low relative humidity (Brun et al. 1993; Portilla 1999c). Temperature is another factor that affects the amount of CBB progeny with low mean number of stages per CBB at low temperature. Portilla (1999c) found that at 23°C occurred the largest preoviposition period and the lowest daily mean of eggs deposited. At 29°, though the mean of eggs daily deposited was higher than at 23°C, 25°C and 27°C the diet at this temperature dried faster and did not allow CBB to keep ovipositing. Developmental time was also affected by temperature, and it was evident the shortest developmental time in each immature stage was at 29°C and the longest at 23°C. The curves of the stages obtained at various times after infestation showed the earlier presence of the stages of CBB at 29°C.
Potential reproduction of CBB on the Cenibroca artificial diets with 50% MC at 85% HR found in this study were observed by Benavides and Portilla (1990) and Portilla and Bustillo (1995) in studies about mass rearing CBB using parchment coffee bean with 45% coffee humidity at 85% RH. They showed up 30 stages per parchment coffee bean per female, and total offspring at 23 ± 4 days was obtained.
Productions reported from artificial diets have demonstrated that the borer can be bred on this substrate for many generations. Brun et al. (1993) obtained ten generations without any significant change in fecundity. One female reared on this diet lived for 380 days (Brun et al. 1993). Villacorta (1989) reported more than 15 generations of borer on his first artificial diet and on the ECOBROVILL-160 diet reported at 85-90 days after the diet has been infested the population reaches between 36,250 to 43,000 individuals per litre of diet.
Although these authors obtained various generation of CBB, no one has reported studies in quality control through generations (Brun et al. 1993, Rojas unpublished). In this study the CBB life table parameters started to deteriorate, including a significant (a=0,05) decline in gross fertility, as well as non significant reduction in fecundity (mean stages produced) and gross hatch rate. Flight capability also declined with significant reduction in maximum flight time and percentage flying being significantly high the fourth and fifth generation in relation to the control.
According to the results these values declined through generations even providing good conditions for its rearing. In studies carried out using parchment coffee bean Portilla, (unpublished) showed that these values had higher variations than using artificial diet. This indicates that under laboratory condition the normal reproduction of this scolytid is affected even if it develops on its own plant host. Similar declines in fecundity occur to the CBB reared on parchment coffee in Cenicafe and the private producer, and so the CBB colonies using a parchment coffee for rearing are renovated every month (Bustillo et al. 1996). All this suggests that there is still a fundamental problem with CBB rearing in the laboratory and can be solved by further researchers on CBB nutrition. However, although problems with the diet remain, this work show that rearing CBB can be make cheaply, thus could pass easily to the next stage which will be the production of the parasitoids Cephalonomia stephanoderis, Prorops nasuta and Phymastichus coffea.
Footnotes
Acknowledgements
This work is an output of the IIBC-CENICAFE collaborative project "Colombia: Integrated Pest Management for Coffee" funded by the Department for International Development (Overseas Development Administration) of the UK and the Federation Nacional de Cafeteros de Colombia. The authors thank all staff at CENICAFE and CABI for their help and unlimited use of the facilities at the headquarters.