
Abstract
An invasion of the llanera locust Rhammatocerus schistocercoides ocurred in 1994 in the Oriental Plains of Colombia, causing damages in agricultural and cattle production of the region. Previous studies of this species demonstrated that three biotypes: green, brown and red are present on individuals of a same population. The phase polymorphism theory express that Acrididae may change of a solitarious to a gregarious phase or viceversa, expresing changes in morphometric characteristics in colorations and in behaviour. These insects are considerated are true locusts. Although R. schistocercoides have a gregarious behaviour is not recognised as a locust, therefore the objective of the work was to verify the presence of phase polymorphism in this specie. Green nymphs were placed in cages individually and grouply and the changes of the coloration was observed in order to describe the morfological and coloration characteristics in females and males adults belonging to biotypes red, brown (gregarious) and green (solitarious). The length of the elytron, hind femur and pronotum, and width of the head were also measured. With this data were made the index elytron/femur (E/F) and femur/head (F/H), recommended to determine differences between polimorphic states. The obtained results permited conclude that exist group effect in nymphs of the species, due to the coloration change toward instar. Significatives differences were also found between E/F and F/H index on green and red biotypes, and in E/F on green and brown biotypes. Moreover, these insects didn't present differences for the E/F index between red and brown biotypes, neither for the F/H index between green and brown biotypes. We could demonstrate that exists phase polimorphism in R. schistocercoides, between green (solitarious) and red (gregarious) biotypes, so this species can be considerated as a true locust.
Introducción
Los modos anormales de fertilización, poliembrionía y fases en los insectos pueden considerarse como tipos inusuales de reproducción, éstos en gran medida dependen de los factores estacionales, incluyendo la disposición del alimento (Raabe 1986). El ejemplo menos complicado de reproducción y fases lo proporcionan las langostas (acrídidos), que difieren en el color, morfología, comportamiento y caracteres fisiológicos distintivos (Joly y Joly 1953; Cassier y Papillon 1968, citados por Raabe 1986).
A pesar de la enorme destrucción causada por las poblaciones de langostas y de la confusión taxonómica entre las especies involucradas que se consideraban como distintas, fue hasta 1913 que una investigación realizada por Plotnikov, demostró que Locusta migratoria capito Saussure y Locusta danica Saussure tenían formas intercambiables dentro de una misma especie. En 1921, Uvarov hizo pública su teoría de las fases, en la que afirma que las langostas se pueden transformar en saltones solitarios y viceversa, y que pueden diferir en su morfología. Estas diferencias, en varias partes del cuerpo, han sido consideradas por Uvarov como "índices morfométricos" que pueden formar proporciones o "radios" con diferencias aún más significativas, que permiten distinguir ambas fases y ubicar una población cualquiera en la fase a la cual pertenece, teniendo en cuenta una muestra de ambos sexos.
Según Beingolea (1995), las especies más peligrosas de los acrididos son las verdaderas langostas, debido a la presencia de dos biotipos o fases; una gregaria y otra solitaria, que cambian de una a otra a través de variaciones o formas intermedias calificadas como "transiens congregans" (de solitaria a gregaria) y "transiens dissocians" (de gregaria a solitaria). Los individuos de cada fase se comportan de manera diferente, donde las gregarias tienden a la aglomeración formando masas densas de individuos de estados ninfales y de adultos que ocasionan daños grandes en áreas de producción agrícola.
En la transformación de fases, la especie de acrídido involucrada es capaz de presentar dos aspectos diferentes que se pueden encontrar en número reducido (fase solitaria), o por el contrario, en cantidades abundantes (fase gregaria). Las transformaciones de una fase a la otra siempre requieren más de una generación para poder ser completadas y son reversibles. El principal factor que provoca este mecanismo en los insectos, y poco frecuente en los acrídidos es la densidad, ya que ciertos efectos de aislamiento o agrupamiento son inmediatos y otros son transmitidos a través de generaciones y se refieren no sólo a la forma exterior, sino también al color del integumento, a ciertos caracteres anatómicos a nivel del aparato reproductor, a la fisiología, al comportamiento, al tipo de desarrollo y a las exigencias ecológicas (Barrientos 1992).
Cada especie gregaria tiene su propia sensibilidad a la presencia de sus congéneres. La densidad que induce a la transformación de fases varía entre 500 y 50.000 individuos por hectárea según la especie. Por ejemplo: las primeras manifestaciones de gregarización de Schistocerca gregaria Forskal, 1775, se observan a partir de 500 individuos por hectárea; mientras que para Locusta migratoria capito es de 20.000 individuos por hectárea.
Los saltones solitarios son de coloración uniforme y generalmente de color verde, mientras que los saltones gregarios presentan diferentes tonalidades de color sobresaliente, de acuerdo con la especie, éstos pueden tener tonalidades oscura, amarilla, rojo y naranja. Se ha investigado que los cambios asociados a la pigmentación responsable de la coloración están relacionados con diferencias bioquímicas. Los saltones gregarios se caracterizan por la presencia de insectorubin y melanina, ambos responsables del color oscuro básico, el cual está ausente en los solitarios. Los solitarios deben su coloración a una mezcla de cromoproteínas amarillas con azul mesobiliverdin ausente en las gregarias (Uvarov 1966).
En el año de 1994, en los Llanos Orientales de Colombia se presentaron densidades poblacionales altas de la especie Rhammatocerus schistocercoides Rehn 1906, lo que se constituyó en una amenaza potencial para la producción agrícola y ganadera de la región (Ebratt et al. 1998). Esta especie fue determinada en el National Resources Institute (NRI), a partir de un especimen colectado en Puerto López por J.A. Jiménez Gómez y corroborado por Lecoq (1996), acridiólogo del PRIFASCIRAD de Francia, quien concluyó, durante su misión de estudio del fenómeno en Colombia, que a diferencia del Brasil, se trató de una invasión de origen local y no de un fenómeno de poblaciones crónicas.
El género Rhammatocerus Saussure (1861) se encuentra ampliamente distribuido desde el sur de los Estados Unidos hasta la Argentina (Otte 1981, citado por Assis-Pujol 1997) y en la actualidad cuenta con 14 especies, consideradas de importancia económica por los constantes daños ocasionados en la producción agrícola de la región. Sin embargo, este género ha sido muy poco estudiado desde el punto de vista taxonómico, resultando en diversos errores de identificación y de distribución geográfica.
R. schistocercoides es gregaria y forma manchas densas de saltones y mangas de adultos. En el Brasil, las áreas de explosión incluyen el estado de Mato Grosso y la Chapada dos Parecis; también ha sido mencionada la existencia de polimorfismo de fases en esta especie sin que hasta el momento haya sido demostrado (Lecoq y Pierozzi 1994a). Esta especie presenta muchas diferencias en las formas cromáticas que tiene el estado adulto (Carbonell 1988 citado por Lecoq y Pierozzi 1994b). En observaciones obtenidas en campo, por Lecoq y Pierozzi (1996), se estableció que el estado adulto cambia progresivamente de marrón a verde como parte de la maduración sexual, lo que es un fenómeno similar al observado en S. gregaria.
Según Ebratt et al. (1998), para el caso de las poblaciones de R. schistocercoides, en los Llanos Orientales de Colombia, existen cuatro tipos fenotípicos en los individuos de una misma población gregaria según el cromatismo presente desde los estados inmaduros. Estos fueron descritos como: biotipo A, aquellos de color pardo o marrón; biotipo B, individuos con manchas color marrón o negro en los bordes laterales anteriores del pronoto y región post e infraocular; individuos biotipo C, de color verde esmeralda (encontrado en poblaciones dispersas en el campo) e individuos biotipo D, de color rojo.
Barrientos (1992) expresó que en las especies polimórficas centroamericanas existen modificaciones cromáticas y morfológicas desde su estado original solitario hasta su completa evolución de fases; es decir, hasta adquirir su forma gregaria y en el cual se consideran individuos solitarios los de color verde, individuos en transición los de color amarillo claro y gregarios los de color rosado con manchas negras.
Según lo descrito por Chauvin (1967), en los animales se ha observado que éstos reaccionan a la proximidad de sus congéneres, no sólo por un fenómeno de comportamiento, sino también por modificaciones fisiológicas (apetito, celo etc.) denominados "efectos de grupo". Este fenómeno se aplica a los efectos de una densidad de población débil sobre los individuos que la componen. Por lo general, resulta en aceleración del desarrollo en comparación con el de los individuos aislados. El efecto de grupo se demuestra cuando dos o más individuos se agrupan en un espacio bastante restringido siendo el criterio de dos individuos una de las mejores pruebas de dicho efecto. Es decir, si no se produce ningún efecto agrupando solamente dos individuos, no podría esperarse efecto de grupo, aumentando la densidad. El efecto de grupo culmina con el fenómeno de las fases, descubierto por Uvarov en las langostas en 1924; debido a que los cambios cromáticos producidos por la agrupación de los acrídidos se consideran como el primer paso para llegar a formar grandes poblaciones con un comportamiento gregario.
Con el propósito de realizar estudios morfométricos en diferentes especies de acrídidos, investigadores como Zolotarevsky (1930), Uvarov (1923), Maxwell-Darling (1934), Rao (1942) y Roonwal (1946) tomaron las medidas de diferentes partes del cuerpo del insecto (longitud del fémur, del pronoto y del élitro, así como el ancho de la cabeza y otras). Posteriormente, debido a que las mediciones directas fueron inadecuadas para el fin práctico de la diferenciación de fases por los métodos estadísticos simples, fue necesario hallar los cocientes entre las medidas que fueran más apropiadas para el caso. Los cocientes son el resultado de la división entre el promedio de una medida y la otra. Así, los cocientes o índices polimórficos más prácticos y comunmente utilizados para distinguir el polimorfismo de fases son el élitro/fémur (E/F) y el fémur/cabeza (F/C) (Dirsh 1953).
Dado que R. schistocercoides no es reconocida como una verdadera langosta, a pesar de su conducta gregaria, el objetivo del presente trabajo fue el de probar la existencia de polimorfismo de fases en R. schistocercoides, considerando la presencia o ausencia del efecto de grupo y las diferencias morfométricas entre los biotipos existentes.
Materiales y Métodos
El presente trabajo fue realizado en el Centro de Investigaciones (C.I) Carimagua, entre los límites de los departamentos de El Meta y El Vichada; municipio de Puerto Gaitán, a los 4°34′29″ latitud norte y 71°20'7" longitud oeste, a una altitud promedio de 150 m.s.n.m.
Determinación del efecto de grupo
En el Hato El Tranquero, ubicado a 10 km al occidente de Carimagua, se realizó un muestreo de individuos teniendo en cuenta la presencia de estadios ninfales dispersos de coloración verde (a partir del sexto instar). Posteriormente, se llevaron a jaulas de 0.5 mβ ubicadas dentro de una casa de malla. Se consideraron cuatro tratamientos: jaulas con un individuo confinado, jaulas con tres individuos confinados, jaulas con cuatro individuos y jaulas con diez individuos. El diseño experimental fue completamente al azar y cada tratamiento contó con tres repeticiones. La fuente alimenticia para los insectos fue con base en cespedones de la gramínea Axonopus purpussi, considerada como una de las especies nativas de la sabana más preferida por el insecto (Ebratt et al. 1998).
Se realizó una observación diaria de los individuos en cada una de las jaulas, teniendo en cuenta los posibles cambios cromáticos en los diferentes estadios hasta el estado adulto. A partir de estas observaciones se realizó una descripción morfológica y de coloración de los individuos y su tipo de confinamiento a que hubieran sido sometidos.
Determinación de índices polimórficos en R. schistocercoides
Con el propósito de determinar los índices polimórficos se realizaron capturas sobre los enjambres de R. schistocercoides existentes en el área de estudio, correspondientes a 60 individuos de biotipo marrón y 60 individuos de biotipo rojo de ambos sexos.
Para la obtención de los 60 individuos de biotipo verde, se realizó una captura de ninfas de este biotipo y se confinaron de forma individual en jaulas de muselina ubicadas en el campo. Mediante un calibrador pie de rey (precisión de 0.02 mm) se llevaron a cabo las mediciones de los diferentes biotipos. Los parámetros medidos fueron el ancho de la cabeza (C) (tomando la longitud entre gena y gena sobre el vértice de ésta), la longitud del élitro (E) (tomando los puntos extremos; es decir, la unión de las venas mediastina y humeral), la longitud del fémur de la pata posterior (F) (tomando los cóndilos externos) y la longitud dorsal total del pronoto (P) (tomando el límite superior de la prosona hasta el límite inferior del metasona) (Dirsh 1953). El tamaño de la muestra estuvo representado por 360 individuos, para un total de 1200 medidas morfológicas.
Los datos morfométricos obtenidos se sometieron a diferentes pruebas para corroborar que cumplieran con los supuestos estadísticos. Éstos fueron, la prueba de normalidad, la prueba de homogeneidad de varianzas y la prueba de independencia de errores, y con todo el universo de muestra se realizó una eliminación de datos atípicos. Posteriormente, cada parámetro a medir de los diferentes biotipos se sometió a un análisis de varianza y éstos se sometieron a intervalos de Tukey y al análisis multivariado de varianzas.
Resultados y Discusión
A partir del confinamiento individual y en grupo de las ninfas de biotipo verde, y de las observaciones a los diferentes estadios, los individuos solitarios mantenidos en grupos de tres a diez por jaula presentaron transformaciones cromáticas después de cada muda, hacia individuos gregarios (color amarillo-naranja). Dicha coloración se inició a partir de la región inferior del dorso del cuerpo hasta abarcar toda su área. La cara interna de los fémures posteriores presentaron un cambio de color del verde esmeralda hacia el azul oscuro, característico de los individuos gregarios. Los ejemplares solitarios, que se mantuvieron aislados individualmente en cada jaula, no presentaron cambio alguno en su coloración.
Al evidenciarse los cambios cromáticos, de los individuos solitarios que se mantuvieron agrupados, hacia el color gregario característico, se constató que existe el efecto de grupo en esta especie. Este hecho podría mostrar la presencia de un fenómeno de polimorfismo de fases en R. schistocercoides en los Llanos Orientales de Colombia, ya que el efecto de grupo se considera como un paso obligado en la ocurrencia de dicho fenómeno de fases en los acrídidos.
A su vez, la presencia de cambios en la coloración de la cara interna del fémur posterior debido al efecto de grupo, posibilitaría revaluar lo expresado por Lecoq y Pierozzi (1996), quienes afirman que la coloración azul en la cara interna del fémur es natural de esta especie; y confirmar lo expresado por Carbonell (1988), quien advierte que en los ejemplares de R. schistocercoides de la serie H.H. Smith colectados en el primer decenio del presente siglo, esa parte es de color marfil. Debido a esto, es posible que la coloración azul no existiera originalmente y, aún más, Rehn (1906) no lo menciona en su descripción original con base en ejemplares recién capturados.
De otra parte, con este ensayo se pudieron diferenciar morfológicamente dos tipos de individuos, describiendo las características básicas con que se identifica la especie, considerando las hembras y los machos:
Determinación del polimorfismo de fases en Rhammatocerus schistocercoides
A partir de los imagos verdes emergidos, que se mantuvieron individualizados en cautiverio, se realizó la descripción morfológica. Únicamente se describieron las hembras verdes, debido a que no hubo emergencia de machos en las jaulas:
A partir de los individuos marrones y rojos capturados en campo abierto que formaban enjambres se realizó la siguiente descripción morfológica:
Estudio morfométrico
Al comparar las medidas de los machos marrones con respecto a las de las hembras, éstas mostraron mayor variabilidad (Tabla 1). Este aspecto también se cumplió para el biotipo rojo; esta diferencia no se pudo establecer para el biotipo verde, debido a que en confinamiento sólo emergieron hembras, sin poder establecer diferencias con respecto a los machos. Debido a la menor variabilidad en los parámetros morfométricos de las hembras y a la falta de adultos machos en el biotipo verde, todo este estudio morfométrico se realizó con hembras.
Varianza morfométrica de las muestras en enjambres de R. schistocercoides
C = Cabeza; E = Élitro; F = Fémur; P = Pronoto; n = tamaño de la muestra
La ausencia de adultos machos en la población de individuos verdes, podría evidenciar la existencia de un inusual tipo de reproducción denominado partenogénesis telitokia, en el cual a partir de huevos no fecundados se desarrollan únicamente individuos hembras (Uvarov 1966).
De la totalidad de datos morfométricos se excluyeron 13 atípicos que sobrepasaron el valor de 2.5 al ser normalizados para una C = 0 y σβ = 1. A partir de las medidas obtenidas en élitros, fémures, pronotos y cabezas, se observaron diferencias claras entre las poblaciones evaluadas en los individuos de coloración marrón, rojo y verde (Figs. 1, 2, 3, y 4).
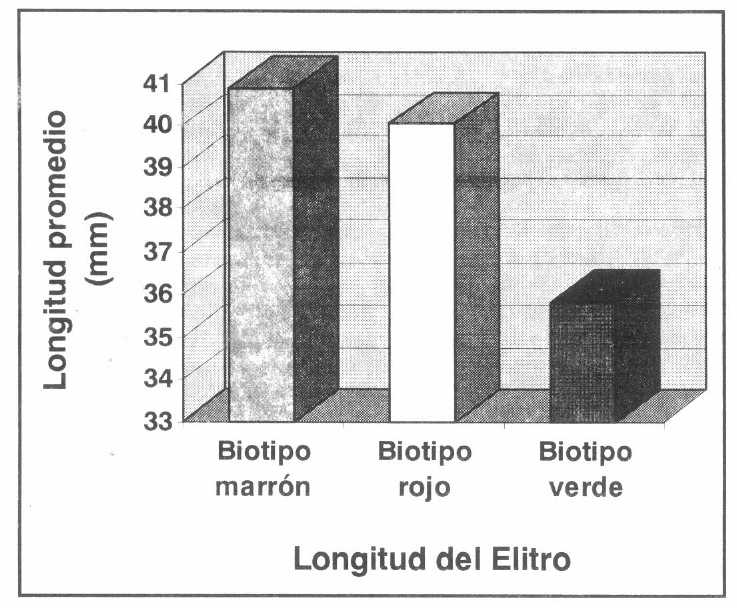
Medidas del élitro en los biotipos de R. schistocercoides.
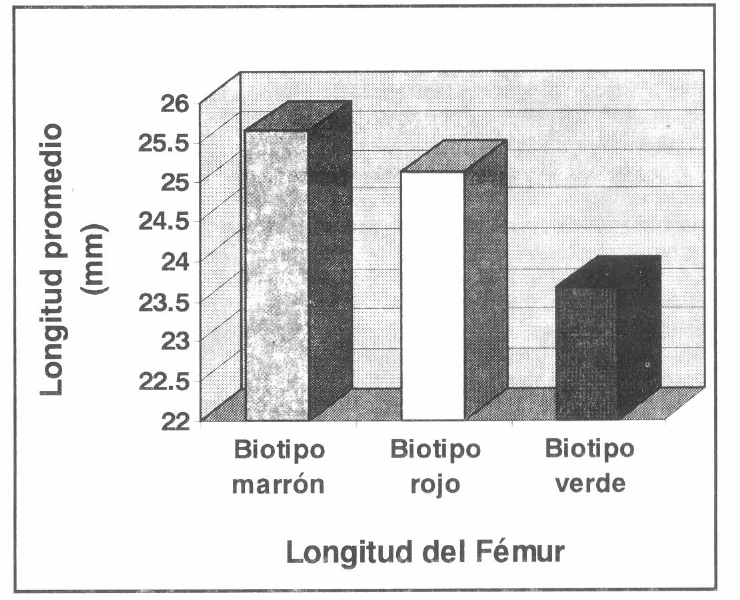
Medidas del fémur en los biotipos de R. schistocercoides.
Los resultados estadísticos del análisis de varianza mostraron que, en el biotipo verde, el ancho de la cabeza fue significativamente menor al de los biotipos marrones y rojos (α=0.01; diferencia mínima significativa (DMS) = 0.0863; MSE= 0.0770; valor crítico de student (VCS) = 4.154), mientras que no existieron diferencias en los biotipos rojos y en los marrones.
Este mismo comportamiento se presentó para las medidas longitud del élitro (α=0.01; MSE=2.7376; VCS= 4.154), longitud del fémur (α=0.01; MSE=1.0394; VCS= 4.154) y longitud dorsal del pronoto (α=0.01; MSE=0.2020; VCS= 4.154); lo que indica que los individuos del biotipo verde son significativamente de menor talla que los individuos de los biotipos marrón y rojo.
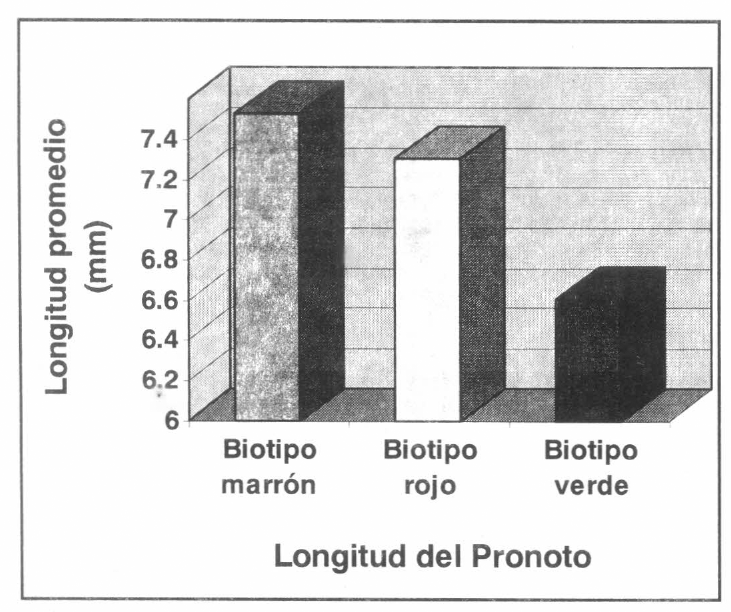
Medidas del pronoto en los biotipos de R. schistocercoides.
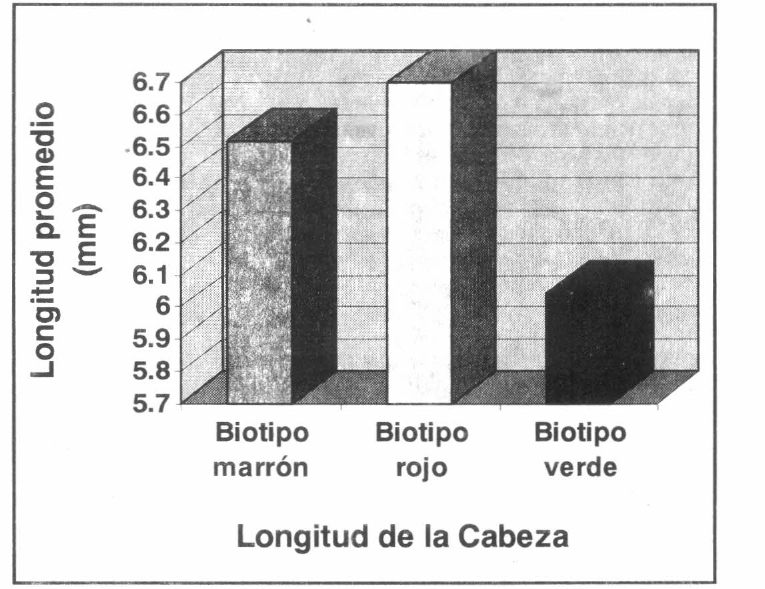
Medidas de la cabeza en los biotipos de R. schistocercoides.
De igual forma, los parámetros evaluados mostraron que existieron diferencias altamente significativas entre las hembras y los machos de los biotipos evaluados; es decir, se reconoció claramente un dimorfismo sexual en la especie.
Estas mediciones indicaron que existe un tamaño mayor en todos los parámetros para las hembras gregarias marrones y rojas en comparación con las hembras solitarias verdes. Además, se encontró que los machos gregarios tienden a mostrar las mismas dimensiones que las hembras solitarias, lo cual es un caso semejante al fenómeno de fases presente en Dociostaurus sp. y Locustana sp. en el Africa (Uvarov 1966).
En ensayos preliminares de estandarización de los datos para el análisis estadístico, se obtuvo que el índice élitro/fémur (E/F) fue significativamente menor en las hembras verdes (1.5157) (Li=1.50; Ls=1.531; α=0.05; Ds=0.0604) que en las hembras marrones (1.5955) (Li=1.576; Ls=1.614; α=0.05; Ds=0.0717) y rojas (1.5921) (Li=1.5785; Ls=1.6057; α=0.05; Ds=0.0522); lo cual está en concordancia con la conducta de los individuos verdes, en comparación con los individuos rojos y marrones; ya que los primeros poseen los fémures más largos en comparación con sus élitros. El índice E/F podría servir como un descriptivo matemático del comportamiento locomotor de cada fase polimórfica en esta especie (Fig. 5).
Así mismo, se obtuvo que el índice fémur/cabeza (F/C) fue menor en las hembras verdes (3.9188) (Li=3.863; Ls=3.973; α=0.05; Ds=0.2112) con respecto a las hembras marrones (3.9441) (Li=3.888; Ls=3.999; α=0.05; Ds=0.2128). Pero estos índices fueron significativamente menores para los individuos rojos (3.7503) (Li=3.7201; Ls=3.7806; α=0.05; Ds= 0.1160). Estos resultados permitirían sugerir que el biotipo marrón es un estado intermedio entre el biotipo verde, de acuerdo con el índice E/F y el biotipo rojo, de acuerdo con el índice F/C (Fig.6).
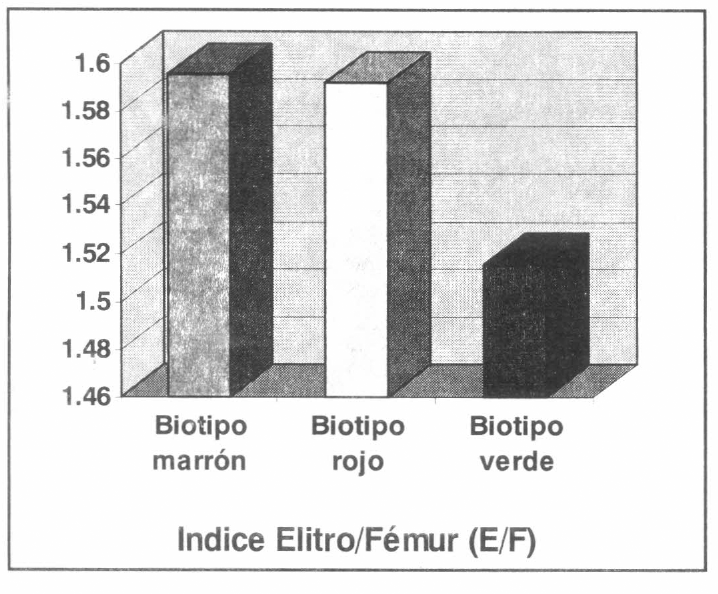
Indice polimórfico Elitro/Fémur en los biotipos de R. schistocercoides.
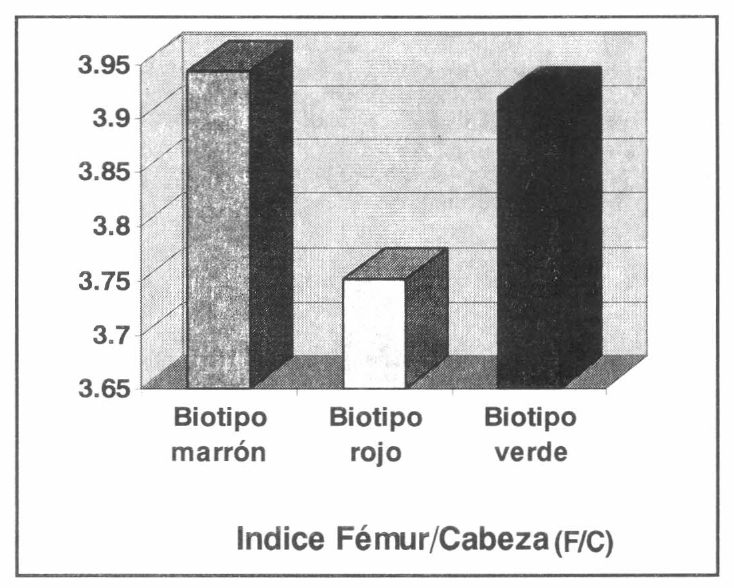
Indice polimórfico Fémur/Cabeza en los biotipos de R. schistocercoides.
El comportamiento de los índices encontrados en el presente trabajo mostró la misma tendencia que para el caso de Schistocerca piceifrons piceifrons (Walker 1870) en Nicaragua (realizado por Astacio 1966; citado por Barrientos 1992), registrada como la única especie con polimorfismo de fases en Centro América.
De acuerdo con lo anterior se podría plantear la existencia, en R. schistocercoides, de un fenómeno de polimorfismo de fases para los Llanos Orientales de Colombia, en la que el biotipo verde correspondería a la fase solitaria, el biotipo rojo a la fase gregaria extrema y el biotipo marrón como una fase gregaria intermedia. Con base en lo anterior, R. schistocercoides se convierte en una especie de gran importancia económica en el futuro desarrollo agrícola y pecuario de los Llanos Orientales de Colombia, debido a que presenta comportamiento de una verdadera langosta y podría esperarse invasiones nuevas por parte de este insecto.
Teniendo en cuenta que este insecto también existe en Venezuela (en el estado de Apure) y en el Brasil (estados de Mato Grosso y Rondonia), sería necesario realizar proyectos de investigación conjuntos con el propósito del manejo integrado de las poblaciones de este insecto.
Conclusiones
Se demostró la existencia del efecto de grupo en la langosta llanera R. schistocercoides.
Existe polimorfismo de fases entre los biotipos verde y rojo de R. schistocercoides, siendo el primero, el estado solitario y el segundo, el estado gregario extremo.
Debido a la existencia de polimorfismo de fases en la especie, R. schistocercoides se puede postular como una langosta verdadera.
Footnotes
Agradecimientos
Los autores expresan su agradecimiento a CORPOICA, ICA Y PRONATTA, por el apoyo financiero para la elaboración del trabajo. A su vez, agradecen a Jaime Albarracín y a Fabio Gaitán, auxiliares del C.I. Carimagua por su invaluable apoyo en la recolección de muestras y toma de medidas; y a Helena Molano I.A., por su constante apoyo en el envío de material bibliográfico.