
Abstract
This study quantified the spore production of 12 isolates M. anisopliae and 10 of B. bassiana, obtained from the orders Coleoptera, Homoptera, Lepidoptera and Orthoptera, these isolates are known to cause pathogenicity levels greater or equal to 80 % in the coffe berry borer. They were cultivated on a rice substrate and coffee berry borers. For the rice substrate cultivation, two bottles per isolate were used, each of them containing 50 grams of rice. The bottles were incubated for 30 days, after which five sporulated rice samples per bottle were taken. The spore count was made from these by using succesive dilutions of the original sample. With regard to the coffee berry borer substrate, 30 individuals per isolate were inoculated and these were incubated in a humid chamber for a 10 day period and afterwards each fungus colonized in the borer was transfererd to a test tube containing 4,5 ml of distilled water, for counting purposes. In the rice substrate, isolates Bb9207, Bb 9301, Ma 9236 presented the greatest number of spores: respectively, 3.57 x 10¹⁰, 1. 53 x 10¹⁰, 1.31 x 10¹⁰ spores/gram. These were statistically different from the rest of isolates; the isolate with the least number of spores was Ma 9303 with 1.65 x 10⁸ spores/gram. In the coffee berry borer substrate, isolate Bb 9218 presented the greatest sporulation with a total of 1.36 x 10⁷ spores/borer, those with the lowest sporulation were eight of M. anisopliae and one of B. bassiana with an average of 5.86 x 10⁵ spores/borer. At the same time pathogenicity and sporulation were observed as independent characteristics. These results demonstrate that spores production depends on the isolate and interaction with the culture media in which develops.
Introducción
Entre los enemigos naturales de la broca, los hongos entomopatógenos han marcado patrones constantes de mortalidad (Villanueva 1990). En el campo, el insecto es atacado naturalmente por diferentes hongos, entre ellos Beauveria bassiana (Vélez y Benavides 1990), Hirsutella eleutheratorum (Posada et al. 1993), Metarhizium anisopliae (Bustillo et al. 1993) y recientemente Fusarium sp. en varias regiones de la zona cafetera (Pérez et al. 1996); en condiciones de laboratorio se ha inducido el ataque de B. brongniartii (Niño et al. 1993).
Beauveria bassiana y Metarhizium anisopliae han mostrado su capacidad para controlar poblaciones de broca; sin embargo, diversos aislamientos de estos hongos presentan variaciones en su eficiencia para atacar dicho insecto (González et al. 1996). Por tal razón, la importancia de esta investigación radica en caracterizar los aislamientos de B. bassiana y M. anisopliae más patogénicos a la broca del café y su producción de esporas tanto en sustratos naturales como en el insecto plaga. La selección de aislamientos con base en la esporulación sobre estos sustratos permitirá diseñar una formulación en la cual se logren niveles óptimos de esporulación (Morales 1991; Olarte 1993) y un control más eficaz en el campo.
González et al. (1993b) estimaron la producción de esporas promedio de algunos aislamientos de B. bassiana, obteniendo entre 2,5 y 8,8 x 10⁹ esporas/broca. Pelaez et al. (1994) estudiaron el desarrollo de dos aislamientos de B. bassiana y M. anisopliae, en sustratos naturales de arroz, cebada, frijol y mucílago bajo condiciones controladas de tiempo, luz y temperatura. La máxima concentracion de esporas se obtuvo de arroz, 2,2 x 10⁹ esporas/g para B. bassiana, y de 5,6 x 10⁸ esporas/g para M. anisopliae.
El objetivo de este trabajo fue cuantificar la producción promedio de esporas de 10 aislamientos de B. bassiana y 12 de M. anisopliae que registran patogenicidad igual o mayor al 80% sobre la broca del café, provenientes de diferentes órdenes de insectos, cultivados en sustrato arroz por 30 días y en el hospedero, broca del café por 10 días.
Materiales Y Metodos
Reactivación de los aislamientos
Los aislamientos a utilizar durante el experimento fueron reactivados sobre broca del café, con el fin de potencializar su patogenicidad (Samsinakova y Kalalova 1982; González et al. 1993b). Las brocas recien emergidas del grano, obtenidas de la unidad de cría de Cenicafé, fueron desinfestadas en hipoclorito de sodio al 0,5% por 10 minutos, tiempo durante el cual cada individuo es liberado de contaminantes externos de fácil remoción. Luego de este tiempo se lavaron por tres veces consecutivas con agua destilada estéril (ADE) y finalmente se colocaron sobre una toalla estéril para su secado.
De cada aislamiento se tomó, con ayuda de un asa, una muestra que se suspendió en 4 ml de ADE obteniendo asi el inóculo. Las brocas fueron expuestas a éste por dos minutos, tiempo suficiente para que el hongo se adhiera al cuerpo del insecto. Posteriormente las brocas se llevaron a cámaras humedas, consistentes en cajas petri con toalla estéril humedecida, cada caja se selló y se llevó a incubación por 10 días a 25 ± 2°C; luego de este tiempo las brocas ya esporuladas se llevaron a una solución de hipoclorito de sodio al 5,25% por 3 minutos y se lavaron una vez conADE; seguidamente, fueron colocadas sobre papel toalla para retirar el exceso de agua y se inocularon individualmente en un tubo inclinado con agar saboraud dextrosa (SDA). Los tubos se llevaron a una temperatura de 25°C hasta el desarrollo del hongo.
Obtención del inóculo
Los aislamientos reactivados de cada uno de los hongos se suspendieron en 9 ml de una solución de ADE + Tween 80 al 0,1% obteniendo de esta forma la solución madre. A partir de ésta se realizaron tres o cuatro diluciones consecutivas en base 10; la estimación de esporas presente se realizó en cámara de Neubauer a partir de la ultima dilución. El promedio de seis recuentos se multiplicó por el recíproco de la dilución en la cual se hizo el recuento y por el factor de corrección de la cámara (10⁴). Para llegar a la concentración deseada, (1 x 10⁷ esporas/ml) se utilizó la fórmula:
Sustrato arroz
Se utilizaron botellas planas de 375 cc donde se prepararon los medios de cultivo asi: se colocaron 50 g de arroz comercial y 80 ml de agua destilada, siguiendo la metodología descrita por Antia et al. (1992). Cada botella se inoculó con 1 ml de la suspensión del hongo en concentración 1 x 10⁷ esporas/ml. Posteriormente, se incubaron a una temperatura de 25 ± 2°C, HR de 70 ± 5% y luz constante por 30 días; las botellas se colocaron en forma horizontal para mejorar la dispersión del hongo en el medio; después de este tiempo de incubación, se estimó la tasa de esporulación presentada por los hongos removiendo el medio de cultivo con ayuda de una espátula, se tomaron 5 gramos del medio ya esporulado de cada botella (Fig. 1) para repartir cada gramo en un tubo con 9 ml de ADE + tween 80 al 0,1%; de esta solución madre se realizaron cuatro diluciones consecutivas en potencia 10 para efectuar la cuantificación de las esporas producidas por aislamiento; se tomaron dos alicuotas de 10 ml con las cuales se llenó la cámara de Neubauer, se realizó el conteo de éstas en los 25 cuadrantes del centro, informando al final el número de esporas presentes en cada retículo; este mismo procedimiento se repitió dos veces más, para obtener 6 recuentos por tubo. La producción de esporas / aislamiento se estimó tomando dos botellas por tratamiento. Para el análisis de los resultados no se tuvo en cuenta el aislamiento Ma 9201 por presentar problemas de contaminación, en el sustrato arroz.
Esporulación de los hongos B. bassiana y M. anisopliae en sustrato arroz a los 30 días de incubación. A, aislamiento Bb 9218; B, aislamientos Ma 9107 y Ma 9232.
Esporulación de B. bassiana y M. anisoploiae sobre la broca del café a los 10 días de incubación. A, aislamiento Bb 9218. B, aislamiento Ma 9211 (Estereoscopio de luz, 40X).
Sustrato broca
Las brocas fueron desinfestadas con hipoclorito de sodio al 0,5% por 10 minutos y lavadas con ADE por tres veces consecutivas; inicialmente se empleó una población de 40 a 50 brocas por aislamiento, las cuales fueron suspendidas en el inóculo por un tiempo de dos minutos y luego se llevaron a platos de petri en cámaras húmedas. Posteriormente, se incubaron a 25 ± 2°C, HR de 70 ± 5% y luz constante durante 10 días. Después de transcurrido el tiempo de incubación se tomaron solo 30 de las brocas inoculadas que presentaron una buena esporulación (Fig. 2). Cada broca se suspendió en un tubo de ensayo con 4,5 ml de ADE + Tween 80 al 0,1% y a partir de esta solución, se realizó una dilución en potencia 10 y se registró el número de esporas por broca utilizando la metodología descrita anteriormente.
Resultados y Discusión
Sustrato arroz
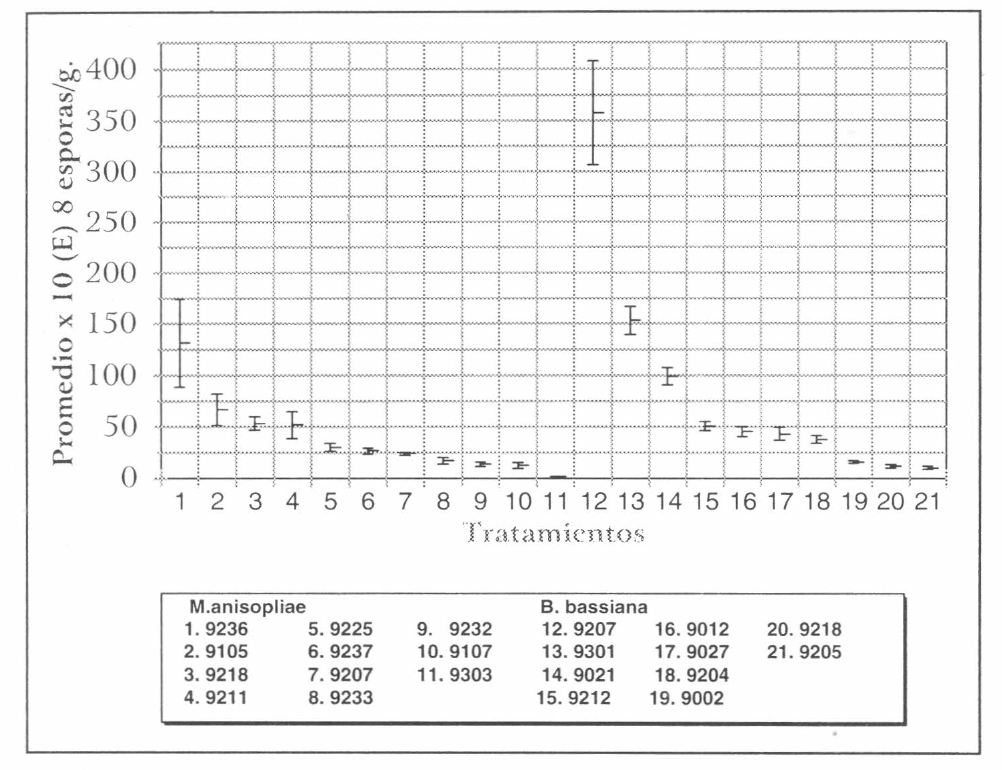
El promedio mayor de esporas para el hongo B. bassiana se obtuvo del aislamiento Bb 9207 con 3,57 x 10¹⁰ esporas/ gramo (esporas/g) obtenido del orden Coleoptera (H. hampei) y el promedio menor para el aislamiento Bb 9205 con 1,03 x 10⁹ esporas/g de Lepidoptera (Diatraea saccharalis). En el caso del hongo M. anisopliae la producción mayor de esporas la presentó Ma 9236, con 1,31 x 10¹⁰ esporas/g (hospedero desconocido) y la menor esporulación el aislamiento Ma 9303 con un promedio de 1,65 x 10⁸ esporas/g fue registrado de Coleoptera H. hampei (Tabla 1). No se observó relación entre la esporulación obtenida de los aislamientos y el hospedero original.
El promedio máximo de esporas se obtuvo del aislamiento Bb 9207 siendo estadísticamente diferente de los demás tratamientos, seguido del aislamiento Ma 9236 el cual no muestra diferencias significativas con los aislamientos Bb 9301 y Bb 9021. Se observa que el tratamiento que tuvo la esporulación más baja, con respecto a los demás, fue Ma 9303 (Fig. 3).
El análisis estadístico de la asociación esporulación - patogenicidad entre los aislamientos de B. bassiana y M. anisopliae no fue significativa en este sustrato. Se observó que el aislamiento Bb 9021, que produjo un promedio de 9,93 x 10⁹ esporas/g, tiene una patogenicidad de 91,7%, mientras el aislamiento Bb 9205, que produjo un promedio de 1,03 x 10⁹ esporas/g, tiene un porcentaje de patogenicidad de 90%. Asi mismo, para M. anisopliae se encontró que el aislamiento Ma 9105 con un promedio de 6,6 x10⁹ esporas/g tiene una patogenicidad del 95% y el aislamiento Ma 9303 con 1,6 x 10⁸ esporas/g una patogenicidad del 92,5%. Lo anterior puede implicar que no hay una asociación entre la esporulación y la patogenicidad.
Con el fin de preseleccionar aislamientos tanto de B. bassiana como de M. anisopliae que permitan obtener buenos promedios de esporulación en el sustrato arroz, se agruparon bajo los siguientes criterios: en el grupo 1, los aislamientos que presentaron un promedio mayor de 100 x 10⁸ esporas/g; grupo 2 entre 50 - 99 x 10⁸ esporas/g; grupo 3 entre 20 - 49 x 10⁸ esporas/g; grupo 4 entre 10 - 19 x 10⁸ esporas/g y en el grupo 5 se incluyeron los que presentaron menos de 10 x 10⁸ esporas/g. El análisis de varianza mostró efecto de tratamientos, y al aplicar la prueba de contrastes ortogonales, bajo el mismo análisis de varianza, se observó que todos los grupos descritos, son diferentes estadísticamente, según prueba f al 5% (Tabla 2).
Se observa que los tratamientos: Bb 9207, Bb 9301 y Ma 9236 (grupo 1) fueron obtenidos del orden Coleoptera (H. hampei y Rhynchophorus palmarum) y hospedero desconocido, respectivamente. Sin embargo, el aislamiento que presentó la esporulación más baja en este mismo sustrato Ma 9303 (grupo 5), fue obtenido sobre Hypotenemus hampei. Contrariamente a lo registrado por Prior (1992), quien menciona que el sitio más probable para encontrar aislamientos que combinen características de virulencia con especificidad de hospedero, es aquel en el cual se presentan éstos en forma constante.
Promedio de producción de esporas/gramo de arroz de los aislamientos de B. bassiana y M. anisopliae evaluados
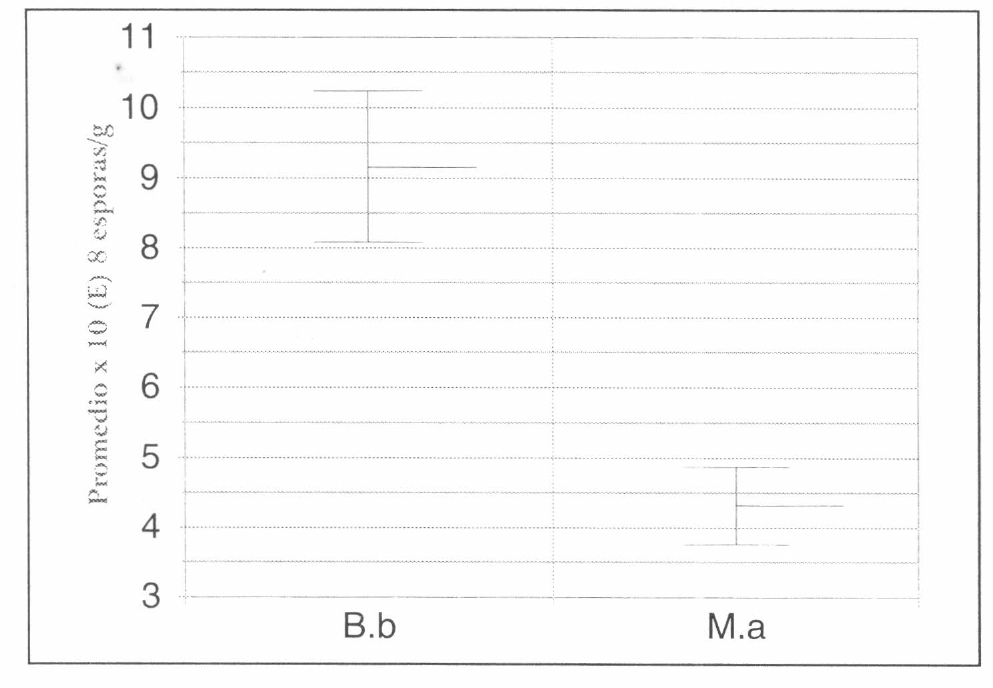
En general B. bassiana obtuvo los máximos promedios de esporulación y al mismo tiempo una mayor variabilidad, mientras que M. anisopliae presentó promedios de esporulación más bajos y una variabilidad menor, como se observa en la figura 4.
Es posible que la esporulación de los aislamientos del hongo B. bassiana en el arroz haya sido favorecida por las condiciones nutricionales que el sustrato ofrece, pues de acuerdo con Olarte (1993), el arroz como cereal recubierto representa una composición de 0,7% de grasa, 1,5% de fibra, 1,2% de minerales y 74,4% de carbohidratos y según Rivera (1993) el efecto de la esporulación se observa cuando el contenido de carbohidratos es mayor que el contenido de proteínas en el medio de cultivo. Antia (1992) y Auld (1992), citan que los valores nutricionales contenidos en el arroz proporcionan los ingredientes básicos que permiten el desarrollo y la propagación del hongo B. bassiana.
Agrupación de los tratamientos de acuerdo con el número medio de esporas producidas en el sustrato arroz
CVT = Coeficiente de variación transformado.
Promedio de producción de esporas/broca de los aislamientos de B. bassiana y M. anisopliae evaluados
Sustrato broca
No se observó relación entre el origen de los aislamientos y la capacidad de esporulación de los entomopatógenos B. bassiana y M. anisopliae; es asi como el aislamiento Bb 9218 obtenido de Coleoptera (Cosmopolites sordidus) y Ma 9211 proveniente del orden Homoptera (Zulia colombiana), presentaron las esporulaciones mayores con un promedio de 1,36 x 10⁷ y 3,92 x 10⁶ esporas/broca, respectivamente. Mientras Bb 9205 obtenido de Lepidoptera (Diatraea saccharalis) y Ma 9303 proveniente de Coleoptera (H. hampei) presentaron los promedios más bajos de esporas, con 8,72 x 10⁵ y 0,72 x 10⁵ esp/broca, respectivamente (Tabla 3). Sin embargo, Paccola y Azevedo (1990) afirman que los aislamientos provenientes de otros insectos pueden ser potencialmente más activos contra insectos de otros órdenes.
El tratamiento mejor fue Bb 9218 siendo estadísticamente diferente de los demás y seguido por Bb 9012, el cual presentó diferencias estadísticas con relación a los otros tratamientos; el aislamiento que produjo la esporulación mínima fue Ma 9303 estadísticamente diferente de los demás (Fig. 5).
El análisis estadístico, agrupando los aislamientos de un mismo hongo por patogenicidad vs esporulación en sustrato broca, mostró que dicha asociación no fue significativa. Se observa que el aislamiento Bb 9012 con 9,66 x 10⁶ esp /broca tiene una patogenicidad de 91,7%; mientras el aislamiento Bb 9205 con 8,72 x 10⁵ esp/broca tiene un porcentaje de patogenicidad de 90%. Igualmente, el aislamiento Ma 9237 con 3,3 x 10⁶ csp/broca tiene una patogenicidad de 92,5% y el aislamiento Ma 9303, que produjo 0,7 x 10⁵ esp /broca, de 92,5%. Lo anterior permite inferir, que la patogenicidad es independiente de la esporulación.
Los aislamientos de B. bassiana y de M. anisopliae se agruparon con base en la producción de esporas en el sustrato broca, en cinco grupos (Tabla 4). El análisis de varianza mostró efecto de tratamientos y al aplicar la prueba de contrastes ortogonales, bajo el mismo análisis, se observó que todos los grupos descritos son diferentes estadísticamente según prueba f al 5%. Cabe anotar que de los aislamientos que conformaron el grupo número 5 se destacó el aislamiento Ma 9303 por el promedio bajo de esporas.
Comparación entre la producción promedio de esporas de cada uno de los aislamientos de B. bassiana y M. anisopliae.
Comparación entre la producción promedio de esporas de cada uno de los aislamientos de B. bassiana y M. anisopliae en sustrato arroz.
Nuevamente se observó que la esporulación expresada por los hongos es independiente del hospedero original. Aún cuando algunos autores afirman que la búsqueda de antagonistas debe iniciarse en el sitio en el cual se desarrolla el insecto o el patógeno en estudio, deben buscarse microorganismos de otros ambientes ya que pueden tener actividad potencial en el control de la plaga (Prior 1992).
Se presentaron diferencias estadísticas significativas entre B. bassiana y M. anisopliae, observando para B. bassiana, los promedios más altos de esporulación y la mayor variabilidad (Fig. 6).
Solo los aislamientos Bb 9021 y Bb 9212 presentaron tanto en el sustrato arroz como sobre la broca del café, un buen promedio de esporas, con 9,93 x 10⁹ y 5,07 x 10⁹ esporas/ g en el sustrato arroz y en el sustrato broca de 6,57x10⁶ y 5,49x10⁶ esporas/broca, respectivamente. Al igual que los aislamientos Bb 9205 y Ma 9303 que produjeron los promedios inferiores de esporulación en ambos sustratos. Estos resultados indican que el grado de esporulación depende tanto de la interacción entre el aislamiento y los componentes del medio de cultivo como de la naturaleza del aislamiento; tal como lo afirman Latge, y Sanglier, citados por Khachatourians (1991), los nutrientes como las vitaminas estimulan la esporulación, otros como los aminoácidos favorecen tanto el crecimiento como la esporulación. Al mismo tiempo afirman que la epicutícula del integumento consta de estructuras que contienen lipoproteínas, grasas y capas cerosas; algunos de estos materiales cerosos poseen un número de sustancias antifúngicas capaces de interferir y prevenir el crecimiento de los hongos entomopatógenos. Griffin (1994) sostiene que la esporulación es generalmente favorecida por condiciones nutricionales que restringen el crecimiento, otras esporulaciónes ocurren por falta de alimento o por agotamiento de nutrientes o por otros estímulos, es decir, la esporulación es una característica continua de los aislamientos que se interrumpe por períodos de dormancia y está directamente relacionada con la acumulacion endógena de reservas apropiadas en los procesos de formación de la espora e influye de manera significativa en la tolerancia de estas a la desecación, en la estabilidad de la formulación y en la eficacia en el control (Jackson 1997).
Monteiro (1988) analizó el comportamiento de aislamientos de B. bassiana, M. anisopliae y Paecilomyces marquandii producido en diferentes medios de cultivo observando diferencias en crecimiento y esporulación entre especies, entre aislamientos de la misma especie y entre los medios evaluados. Las diferencias encontradas en un mismo aislamiento cuando es cultivado en diferentes sustratos, se debe a la diversidad de nutrientes existentes en los medios y pueden ser consideradas como consecuencia de la variabilidad genética particular de cada aislamiento (Paccola y Azevedo 1990).
Conclusiones
La esporulación de Beauveria. bassiana y Metarhizium. anisopliae depende del aislamiento y la interacción entre éste y la composición del medio de cultivo.
Agrupacion de los tratamientos de acuerdo con el número medio de esporas producidas en el sustrato broca.
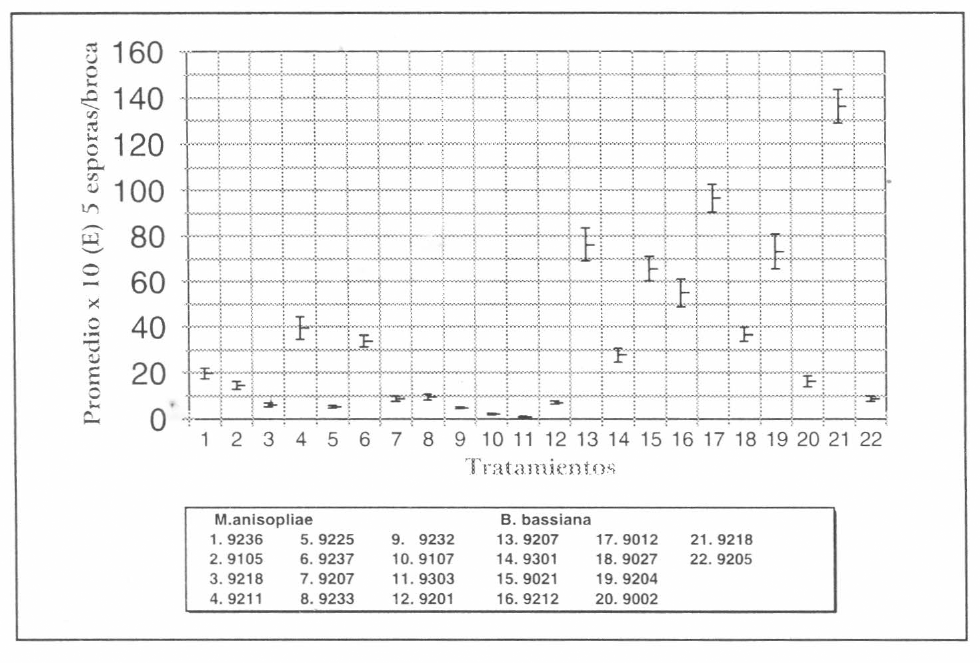
Comparacioón entre la producción promedio de esporas de cada uno de los aislamientos de B. bassiana y M. anisopliae en sustrato broca.
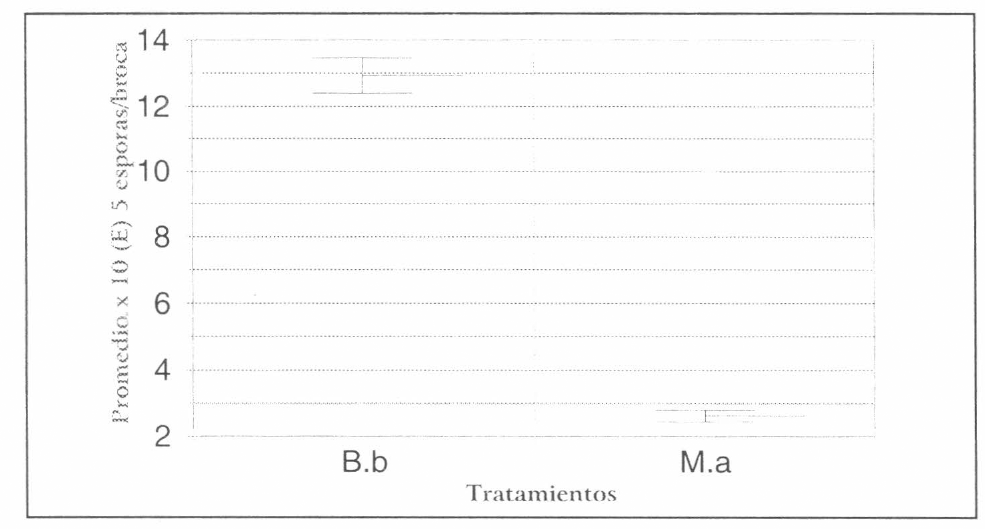
Comparacion entre la producción promedio de esporas de todos los aislamientos de B. bassiana y M. anisopliae en sustrato broca.
Las diferencias presentadas por un mismo aislamiento, cultivados tanto en sustrato arroz como en la broca del café, se deben al aporte nutricional del sustrato y a la capacidad del hongo para utilizar dichos compuestos.
Los aislamientos Bb 9303 y Ma 9205, presentaron los promedios más bajos de esporulación en ambos sustratos.
Algunos aislamientos producen una mayor esporulación cuando se desarrollan sobre el hospedero original, otros, desarrollan mejor esta característica cuando atacan hospederos diferentes a éste.
No se observa relación directa entre la patogenicidad y la esporulación de los aislamientos de B. bassiana y M. anisopliae evaluados en este experimento.