
Abstract
The difficult to define the species limits of Ectemnaspis or Psilopelmia arise from two opposite points of view from Coscarón (1984, 1987 y 1990) and Crosskey (1987). To define the monophyly and the sister group of Ectemnaspis-Psilopelmia the data from Coscarón (1987) and Coscarón et al. (In press) were bootstraped. To know the subgeneric position two different information sources were used: morphology and chromosome banding. In the cladistic examination from morphological data, fourteen taxa were selected, which cover all the possibilities to be estimated as Psilopelmia or Ectemnaspis from both authors or have problems in the subgeneric position. Forty eight characters from larva, pupa, adult female and rnale were choosen. As sister group the species S. oviedoi- S. rivasi, S. pertinax and S. subpallidum were used. In the chromosome analysis seven species and 12 characters were used: principal and secondary markers from chromosome I - III. The species represent the bicoloratum, romanai and dinellii groups. As sister group it was selected the Simulium subgenus map. With the morphological and chromosomal data, separated and as a set, a parsimony cladistic analysis was conducted. The results show at least five independent units: romanai, bicoloratum, dinellii, perflavum and Psilopelmia. To be consistent with the actual nomanclature the sub-groups must get the subgeneric status.
Introducción
La cladística se ha convertido en una matriz disciplinar en sistemática, debido a que es un esquema de clasificación altamente informativo que recoge los aspectos históricos de los taxa. La taxonomía de dípteros, y en particular la de simulí-dos, no ha sido ajena a este fenómeno, de hecho, como lo resalta Craw (1992), Wygodzinsky, quien fue uno de los investigadores más importantes en simulíidos, jugó un papel preponderante en la difusión de la obra de Hennig en los Estados Unidos.
Aunque podría considerarse que todos los problemas han sido resueltos, todavía se presentan dificultades en la asignación de especies a distintas unidades supraespecíficas. Para resolver un problema particular en la delimitación de especies entre los subgéneros Simulium (Ectemnaspis) o Simulium (Psilopelmia), se plantea el análisis cladístico, a nivel morfológico y citológico, de algunas especies seleccionadas para establecer la validez de cada unidad monofilética. Debido a la estructura de la tarea, el trabajo está dividido en tres partes: en primer lugar se ubican las distintas definiciones de género, que también son aplicables a subgénero; en segundo lugar se revisa la historia de la clasificación para Ectemnaspis y para Psilopelmia, lo que da indicios de las razones de las dificultades; y por último se reconstruye la filogenia a partir de datos morfológicos y citológicos, por separado y en conjunto, además de plantear la situación nominativa propuesta por la filogenia.
Definición de términos
Cladística
Clasificar es generar hipótesis sobre distintos objetos a partir de las propiedades que estos objetos poseen (Sober 1988). Originalmente, el objetivo de la taxonomía y más tarde de la sistemática, fue el de crear clasificaciones; aunque este es un punto de inicio, a la actual sistemática le atañen otro tipo de hipótesis. La sistemática filogenética o cladística busca reconstruir la historia de los organismos basada en el principio de novedad evolutiva compartida, es decir aquella característica homóloga que poseen dos o más especies, que indica que tienen mayor afinidad filogenética entre sí que con cualquier otra especie (Hennig 1968).
Para un análisis filogenético se realiza una serie de pasos claramente definidos:
Selección de grupos. Donde se especifican las unidades de comparación, bien sea especies dentro de un género o géneros dentro de una familia; o taxa en general que en primera instancia corresponden a cualquier categoría del sistema Linneano de clasificación, los cuales se espera representen una entidad monofilética. Determinación de homologías (caracteres homólogos). La filogenia no se puede realizar con todos los caracteres, sino sólo con los caracteres homólogos. En general, se utiliza el criterio de homología por estructuraontogenia y no por función, evaluable por una prueba triple de similitud-congruencia filogenética-conjunción. Para decidir la polaridad se compara con un grupo externo o por ontogenia. Sin embargo, el tema es lo suficientemente amplio, por lo que se recomienda revisar el libro editado por Hall (1994) que pone de manifiesto las dificultades para la asignación de homología dependiendo del tipo de fuente de información, morfología, moléculas, etc. Agrupamiento por parsomonia. Una vez se han delimitado los caracteres homólogos, se agrupan las unidades según el principio de parsimonia; es decir, se prefiere la hipótesis que indica el menor número de cambios (Wiley 1981). El resultado del agrupamiento o hipótesis de parentesco es diagramado como un árbol enraizado, que se denomina cladograma. Evaluación de la calidad de la hipótesis de parentesco frente a la información original. Debido a que en general no todos los caracteres son congruentes entre sí, se presentan hipótesis con mayor o menor grado de información, dependiendo del grado de conflicto entre caracteres. Tradicionalmente se han utilizado dos índices: el de consistencia (C.I.) propuesto por Kluge y Farris (1969) y el índice de retención (R.I.) propuesto por Farris (1991). Para examinar las bondades de cada valor se sugiere revisar a Siebert (1992).
El cladograma final puede ser utilizado para contrastar distintas características de los organismos, desde su comportamiento hasta la coevolución (Fig. 1); en este trabajo se visualiza la validez de cada subgénero y la situación nominativa que implica la filogenia.
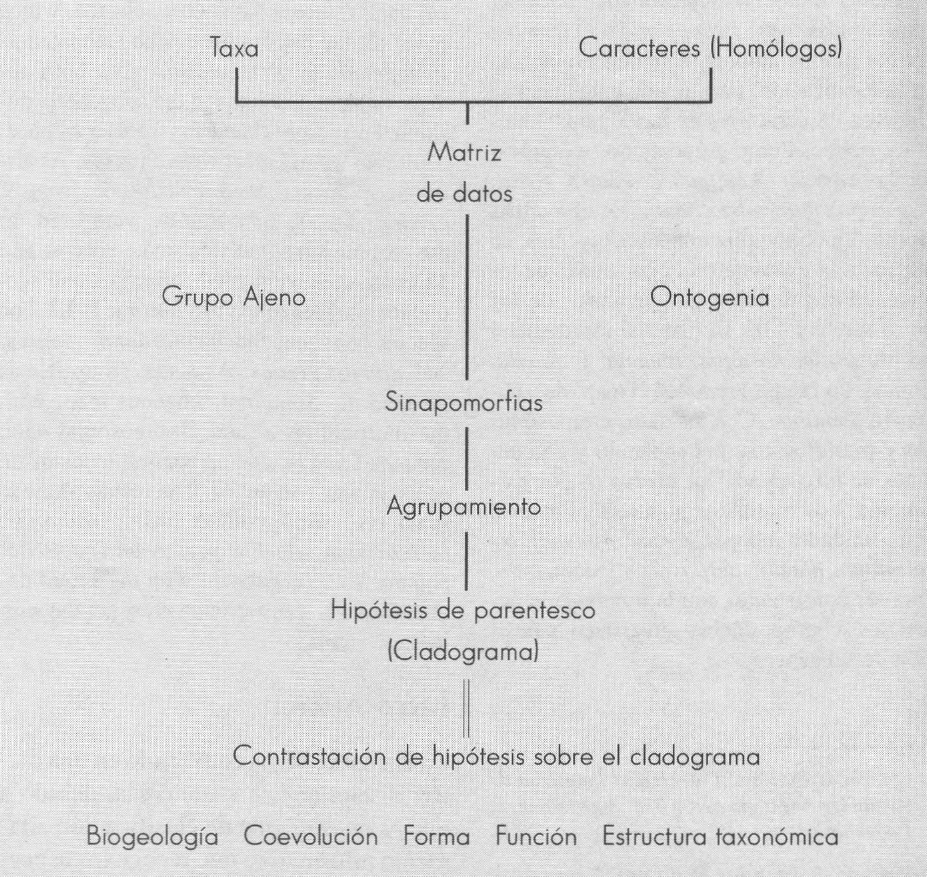
Principios generales del sistema filogenético de clasificación.
Los conceptos de género
La palabra género y lo que ella define es un aspecto controversial desde la categoría propuesta por Linneo y por ende mandatoria en los códigos de nomenclatura, hasta la condición de inexistencia que mantiene la escuela tradicional, hasta llegar a la delimitación filogenética de los taxas supraespecíficos. Sobre el esquema filogenético contemporáneo se pueden proponer las definiciones de Papavero y Llorente (1992), quienes estiman que existen dos percepciones de género como unidad supraespecífica:
El C-género (género de catálogo) como resultado de los postulados aristotélicos sobre especies ínfimas (basadas en individuos "tipo"). Debido a que estos grupos son abstracciones o géneros formados a partir de especies, que a su vez son abstracciones, serían en última instancia abstracciones de abstracciones, o meta-abstracciones, y por lo tanto no poseerían existencia real en el sentido espacio-tiempo. Según esta definición son clases y no individuos (sensu Hull 1976). Las especies que lo conforman son grupos terminales a la filogenia. El F-género (género filogenético) es el resultado de la adopción de la teoría de la evolución, definido como el conjunto de tres o más especies que componen un sistema monofilético (sensu Hennig 1968); es por lo tanto el conjunto de todas las especies de una filogenia.
A partir del F-género, la pertenencia a un grupo (histórico) puede ser corroborada acorde con la distribución de caracteres homólogos dentro y entre los grupos involucrados, para obtener como resultado grupos monofiléticos nomenclados según la localización de la especie tipo (Miranda y Muñoz de Hoyos 1996).
El estado actual de la determinación Ectemnaspis / Psilopelmia
Los obstáculos para decidir la posición taxonómica de un grupo y su nombre dentro de la familia Simuliidae (Diptera) se remontan a principios de este siglo y continúan hasta el presente. Ampliar en tal discusión está fuera de los alcances de este trabajo. Para una revisión se sugiere consultar a Miranda y Muñoz de Hoyos (1996). La dificultad actual en la asignación de límites se presenta a partir de dos propuestas:
Coscarón (1984, 1987, 1990) quien incluye en Ectemnaspis 39 especies asignadas a cuatro subgrupos: bicoloratum, romanai, dinellii y perflavum.
Crosskey (1987, 1990), quien en su lista mundial considera al subgénero Ectemnaspis conformado por 14 especies, casi las mismas del subgrupo bicoloratum sensu Coscarón (1990), y al subgénero Psilopelmia conformado por 50 especies, las cuales son casi las mismas que considera Coscarón en los subgrupos romanai, perflavum y dinellii.
De hecho, la dificultad estriba en la asignación de los grupos dinellii, perflavum y romanai, ya que en opinión de Crosskey son más cercanos a S. escomeli, especie tipo del subgénero Psilopelmia, que a S. bicoloratum, especie tipo del subgénero Ectemnaspis.
Reconstrucción de la Filogenia: Materiales y Métodos
Datos preliminares como aproximación a la monofilia
Como labor preliminar, y para evaluar la existencia de los subgéneros como grupos monofiléticos y establecer la posición de los distintos taxa, se reanalizaron los datos de Coscarón (1987). A partir del cladograma se seleccionaron dos series de datos: la primera compuesta por los subgéneros Psilopelmia, Ectemnaspis, Simulium y Chirostilbia, y la segunda excluyendo el grupo Simulium. Una tercera serie estuvo conformada por los datos para la filogenia del subgénero Psilopelmia, donde se incluyen los grupos bicoloratum, romanai, perflavum y dinellii (Coscarón et al. En prensa).
Con el primero y segundo conjuntos se buscó, así mismo, una aproximación al grupo hermano de Psilopelmia - Ectemnaspis. Coscarón (1987) considera a oviedoi como tal, pero en los análisis de Psilopelmia - Ectemnaspis utiliza a Chirostilbia como grupo ajeno.
El subgénero Simulium se involucra, ya que en el primer análisis este grupo se situaba en algunos casos entre Chirostilbia y el grupo oviedoi.
Elección de taxa
Para el estudio de la filogenia establecida por caracteres morfológicos se escogieron 14 taxa por la disponibilidad de material para examen y por representar la variabilidad posible de selección sensu Coscarón (1990) o Crosskey (1987) (Tabla 1).
Listado de especies seleccionadas para la reconstrucción de la filogenia a partir de caracteres morfológicos, fuente de información utilizada y posición subgenérica
Aunque la especie fue descrita con posterioridad a la lista, no se alberga duda sobre su inclusión en Ectemnaspis.
La mayor parte del material se revisó directamente con apoyo de bibliografía, la lista de especímenes trabajados se encuentra en la Tabla 1. Para la especie S. bipunctatum se utilizó fundamentalmente la redescripción de Shelley et al. (1989), que contempla la mayoría de los caracteres trabajados (Tabla 2). Las especies recogen todas las posibilidades, desde especies que son consideradas como Psilopelmia o Ectemnaspis por ambos autores, hasta especies que presentan problemas en la localización.
Las especies S. pertinax, S. subpallidum del subgénero Chirostilbia y S. rivasi - S. oviedoi del grupo oviedoi se involucraron como grupo ajeno.
Para el análisis cromosómico se utilizaron siete especies: S. bicornutum, S. furcillatum, S. tunja, S. ignescens, S. ochraceum, S. lutzianum-lewisi y S. romanai. El material representa a los grupos bicoloratum, romanai y dinellii. Como grupo ajeno se emplearon los mapas del subgénero Simulium, ya que no existen estudios citológicos para S. oviedoi - S. rivasi o para especies del subgénero Chirostilbia. El menor número de especies para el trabajo cromosómico obedece a la dificultad de obtener mapas de todas ellas.
Aunque no se obtuvo material que represente a S. perflavum y a Psilopelmia sensu stricto, la hipótesis propuesta se convierte en un punto de comparación que permitirá relacionar e integrar la información adicional que se obtenga en el futuro,
Definición de carácter
La definición de carácter en este trabajo sigue los lineamentos de Inglis (1991) у Nelson (1994), quienes plantean la existencia de caracteres como componentes homólogos derivados, y también se sigue la formalización como elemento de lógica de conjuntos de Papavero et al. (1993).
Caracteres morfológicos
Para los distintos taxa se eligieron 48 caracteres: de larva (caracteres 1 al 7), pupa (caracteres 8 al 20), coloración de adulto (caracteres 21 a 25), hembra (caracteres 26 a 36) y macho (caracteres 37 a 48) (Tabla 2). De los 132 caracteres considerados inicialmente, se descartaron algunos que no presentaban variación que delimitara grupos, tal es el caso de la onchotaxia (distribución de pelos en la pupa), que es similar entre S. bicoloratum y S. escomeli. Igualmente se descartaron caracteres como la coloración de la larva o el patrón del apotoma cefálico de la misma, ya que en todos los análisis preliminares el Indice de consistencia reescalonado fue cero.
Algunos de los caracteres son multiestado y otros son datos continuos. Aunque existe controversia sobre el uso de los datos multiestado, es de anotar que la división en datos binarios sobre la misma estructura no es coincidente con el espíritu de este trabajo, ya que la fragmentación a caracteres doble estado sería un poco irreal y haría necesario considerar un gran número de caracteres como ausentes o no comparables.
Los estados de carácter que se utilizaron son aquellos que permiten evaluar la homología como hipótesis dentro de un marco de sinapomorfia (Inglis 1991; Nelson 1994). El patrón de color no posee un trabajo filogenético preliminar y su codificación es bastante compleja debido a todas las posibilidades de variación en tono y amplitud de las marcas, por lo tanto se contemplaron dos posibilidades: negro o en general oscuro y con color, que para las especies evaluadas puede ser amarillo, naranja y las variaciones entre ellos.
Los cocientes y los valores numéricos se consideraron como ordenados y la codificación se hizo por segmento, pero los tamaños de intervalo no son siempre iguales, debido a que se buscó recoger al máximo la variabilidad, pero a su vez reducir al mínimo las sobreposiciones (Chappill 1989); los caracteres multiestado fueron codificados como no ordenados (Hauser y Presch 1991).
Matriz de carácteres morfológicos y su codificación.

Caracteres cromosómicos
Para el análisis a partir de datos cromosómicos se utilizaron 12 caracteres de los cromosomas I - II - III (Tabla 3). Los mapas cromosómicos provienen de distintas fuentes: para las especies S. bicornutum, S. furcillatum, S. tunja, S. ignescens, S. romanai los mapas han sido fruto de los trabajos del grupo de investigación en simulíidos en el Instituto de Ciencias Naturales Universidad Nacional de Colombia (Campos 1989; Campos y Muñoz de Hoyos 1990; Duque 1980; Duque et al. 1988; Miranda 1991; Miranda y Muñoz de Hoyos 1993). Los otros mapas se obtuvieron de algunas tesis de doctorado: el citotipo A de S. ochraceum (Millest 1989; 1992) y la especie A de S. lutzianum-lewisi (Sawyer 1991); esta última referencia de un conjunto, obedece a la imposibilidad de Sawyer de separar las dos especies a nivel morfológico, de tal forma que se pueda soportar una diferencia cromosómica a nivel de larva; es necesario recalcar que la diferencia entre las especies está dada por la coloración en los segmentos abdominales en el estado de adulto. A partir de esta semejanza cromosómica y morfológica Shelley (En prensa) plantea que los dos epítetos son sinónimos.
Matriz de carácteres cromosómicos y su codificación

Pruebas preliminares
Con los datos de Coscarón (1987) y Coscarón et al. (En prensa) se realizó una serie de pruebas de permutación con el programa Bootstrap del paquete PHYLIP, versión 3.1 (Felsenstein 1985, 1988), para datos binarios o para datos multiestado no ordenados, previamente convertidos a binarios.
En el análisis de «bootstrap» se establece un número denominado P para la aceptación de la pertenencia al grupo; este límite ha recibido críticas al considerarlo una cifra estática que no puede ser siempre la misma para todas las posibilidades de número de taxa y caracteres (Sanderson 1989; Hillis y Bull 1993). Esta cifra puede ser recalibrada para reducir la probabilidad de rechazar la monofilia cuando ésta es cierta (Rodrigo 1993), a lo que se le puede llamar una evaluación dentro del contexto de los datos existentes. Como lo recalcan Felsenstein y Kishino (1993), es posible juzgar los valores altos de P como indicadores de confianza de la exactitud de obtener el grupo considerado en el árbol verdadero [c.f. la 'modesta propuesta' de Felsenstein y Kishino (1993)].
Reconstrucción de la filogenia
Para el análisis filogenético a partir de los datos morfológicos y cromosómicos, tanto en forma separada como en conjunto, se realizó un análisis por parsimonia usando el programa Hennig86 (Farris 1988).
Para delimitar las homologías dentro de los datos morfológicos y cromosómicos se utilizaron varios criterios: en primer lugar se rechazaron caracteres para los cuales no se poseían datos de la mayoría de grupos, se aplicó como regla a priori el descartar datos en los cuales se desconociese el estado de más de tres taxa. Los caracteres no polarizables con el grupo ajeno usado también fueron descartados, ya que sólo se consideraron las sinapomorfias.
En el análisis conjunto de los datos se le dió peso diferencial a los caracteres cromosómicos, ya que su menor número indicaba que prevalecería la historia evidenciable por morfología. Para la asignación de peso se planteó la ecuación:
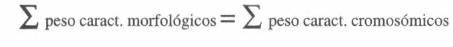
la cual iguala el peso de las dos historias.
Estructuración de los datos morfológicos y cromosómicos
Con los datos morfológicos y cromosómicos por separado se ejecutó una serie de pruebas de permutación PTP (permutation tail probability), para establecer el grado de estructuración de los datos (Faith 1991; Faith y Cranston 1991, 1992). Para esta prueba se usó el paquete Random cladistics 2.1.1 (Siddall 1995), que permuta los caracteres y crea como salida un archivo para ser ejecutado en Hennig86 (Farris 1988), donde se emplea un enfoque heurístico: mhennig, a partir del cual se evaluó la longitud de los distintos árboles obtenidos.
Hipótesis de clasificación por similitud
Con el ánimo de evaluar las posibles hipótesis de clasificación por similitud fenética se construyó una matriz de distancia de Gower (1971) con todos los caracteres, sobre el principio que para tal tipo de agrupamiento se debe utilizar el mayor número de caracteres. Los datos fueron agrupados por UPGMA. Para el análisis y la generación del fenograma se utilizó el programa NTSYS-pc versión 1.70 (Rohlf 1992).
Como base de datos para el manejo de la información de caracteres se utilizó el programa DELTA (Dallwitz 1980; Dallwitz et al. 1993; Partridge et al. 1993).
Reconstrucción de la Filogenia: Resultados y Discusión
Evaluación de los grupos monofiléticos
Al plantear el criterio de permutación por <<bootstrap>>, para el conjunto de datos de Coscarón (1987), se empleó una estrategia de reparametrización del valor P. Al seguir el protocolo de Rodrigo (1993) y usar como grupo de prueba (G) Psilopelmia - Ectemnaspis se obtuvo un valor de 80% para aceptar la monofilia, con 10 ejecuciones de 50 seudorréplicas cada una.
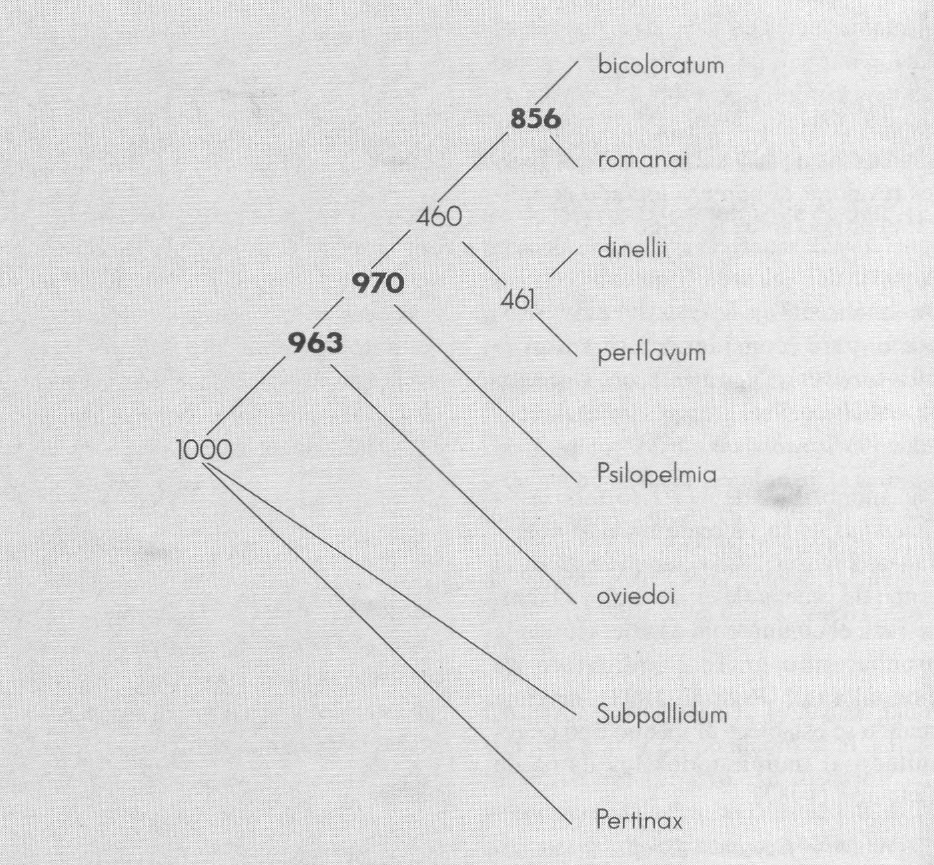
Arbol de bootstrap con 1.000 seudorréplicas a partir de los datos de Coscarón (1987).
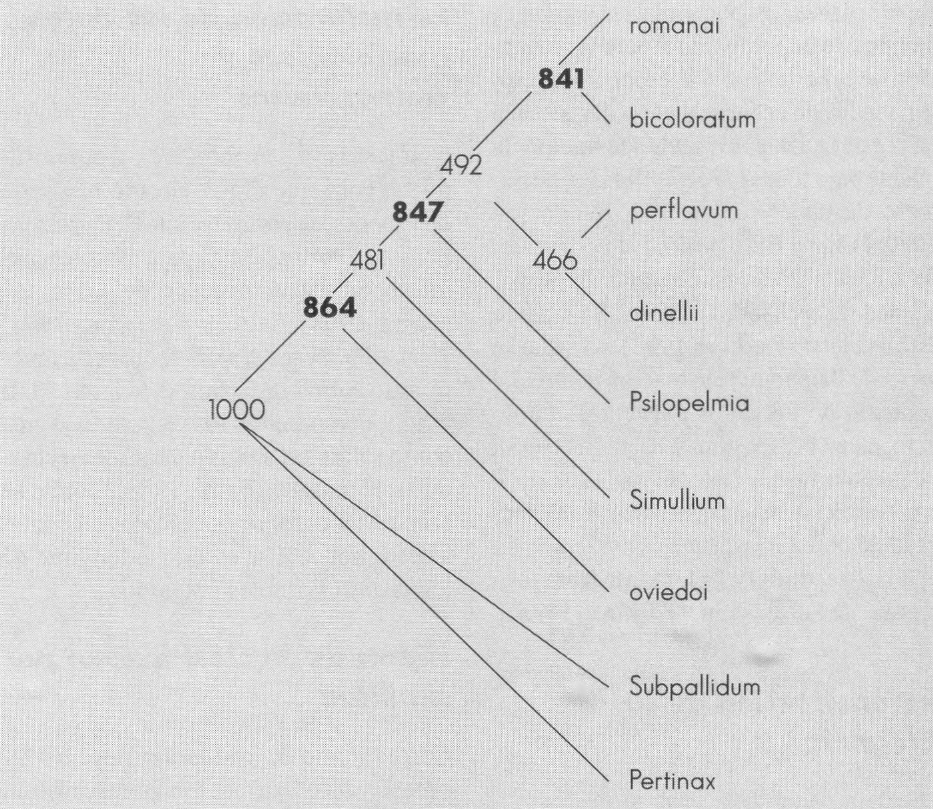
Arbol de bootstrap con 1.000 seudorréplicas a partir de los datos de Coscarón (1987), sin incluir el subgénero Simulium.
La prueba final se hizo con una ejecución independiente de 1.000 seudorréplicas (Fig. 2). La prueba se repitió excluyendo el subgénero Simulium (Fig. 3); se aprecia que se modifica levemente la topología, pero los valores de cada clado se ven fuertemente afectados, sobre todo aquellos de dudosa monofilia.
Con el tercer juego de datos y siguiendo el mismo protocolo se alcanzó un valor de 85%. La prueba final se realizó en la misma forma anterior (Fig. 4).
Aunque existe amplia discusión sobre la verdadera significancia del valor obtenido por <bootstrap> como indicativo de la exactitud de la filogenia (Hillis y Bull 1993; Felsenstein y Kishino 1993), la revaluación del valor de P, como prueba dentro de contexto, permite la obtención de un valor más realista, que no es estático y que depende de los datos.
Evidentemente es necesario hacer pruebas de evaluación de «bootstrap>> con datos no decisivos (sensu Goloboff 1991) y totalmente decisivos, que posean el mismo número de taxa y caracteres que los datos originales, sobre los cuales se pueden determinar los valores mínimos y máximos de P, y sobre estos dos límites revalorar el número logrado al aplicar el protocolo de Rodrigo (1993).
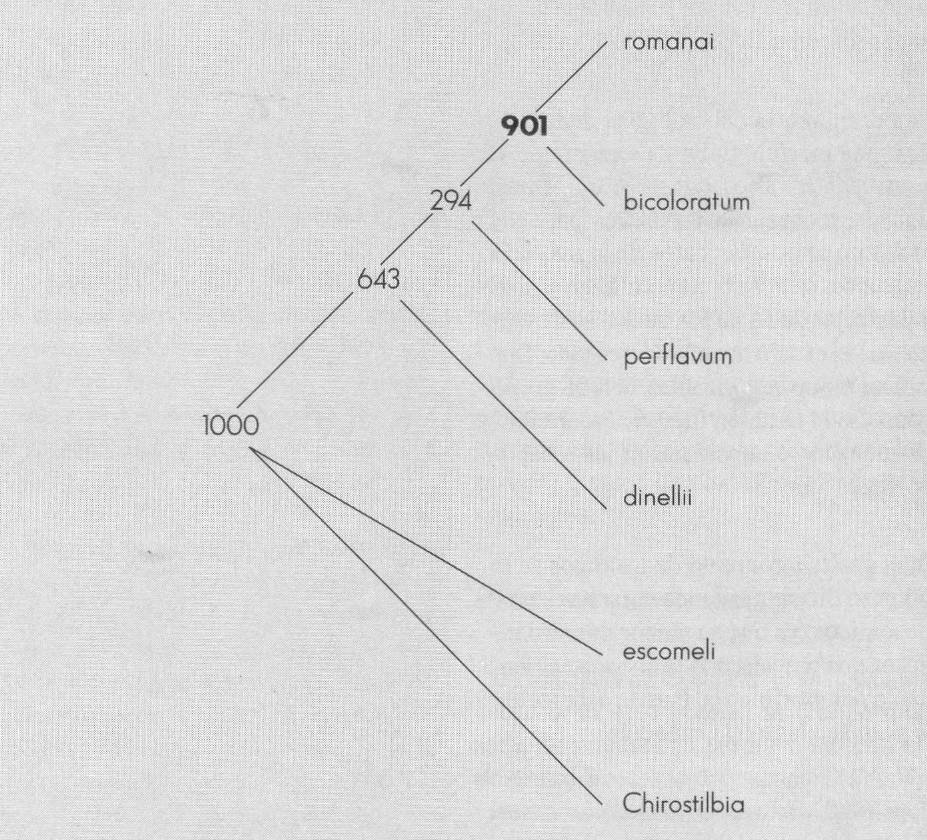
Arbol de bootstrap con 1.000 seudorréplicas a partir de los datos de Coscarón et al. (En prensa).
A partir del valor de P obtenido en los tres análisis (Fig. 2-4) existe mayor soporte para considerar romanai - bicoloratum como un grupo monofilético. Este grupo es independiente de perflavum - dinellii.
La monofilia de Psilopelmia - Ectemnaspis se ve reafirmada al observar las Figuras 2-4, y aunque este es el grupo de prueba G, el alto valor obtenido para el conjunto no es artefacto de la prueba, sino un real indicativo de monofilia (c.f. Rodrigo 1993), máxime cuando se establece el mismo tipo de resultado al incluir todos los datos de Coscarón (1987).
El grupo perflavum - dinellii no es soportado por los datos utilizados como grupo monofilético, ni como conjunto independiente formado por los dos subgrupos, ni como parte de Ectemnaspis. Sin embargo, esto no apunta a que sea parte de Psilopelmia, más bien propone que puede constituir un clado separado de Psilopelmia o Ectemnaspis.
La localización de Simulium entre Psilopelmia - Ectemnaspis y oviedoi no permite delimitar con claridad el grupo hermano de Psilopelmia - Ectemnaspis, por lo que es necesario realizar una prueba más exhaustiva, incluyendo los grupos en cuestión: Psilopelmia, oviedoi, Chirostilbia y Simulium e involucrando información adicional de tipo morfológico, cromosómico o molecular para S. oviedoi - S. rivasi.
Al no incluir a Simulium se reducen los valores para la aceptación de Ectemnaspis como un subgénero de cuatro grupos, pero en ninguno de los dos cladogramas se presentan valores altos para el grupo conformado por romanai - bicoloratum - perflavum - dinellii.
La propuesta que plantean los tres árboles es la existencia de al menos tres grupos: el primero bicoloratum - romanai, el segundo perflavum - dinellii, los cuales no parecen ser un grupo monofilético, y por último Psilopelmia.
Filogenia morfológica
Para los 17 taxa estudiados se presenta el consenso (Fig. 5) a partir de los siete árboles generados con un enfoque heurístico: mh* y bb (Farris 1988). Para efectos del cálculo de estadísticas se redujo el <<outgroup>> a un solo nodo.

Cladograma (datos morfológicos) para los grupos considerados. Consenso estricto de 7 árboles. longitud=245 CI=0,49 RI=0,52. (ptp<0,05).
Se observan tres clados bien determinados: [Chrirostilbia-oviedoi], [Psilopelmia] y [bicoloratum]. Dinellii no es un clado monofilético, ya que sus distintos miembros no se localizan como única rama. Al no poseer réplica del subgrupo perflavum, su monofilia es incierta y para tal caso deben seguirse considerando como válidos los límites dados por Coscarón (1990). La politomía que se observa imposibilita definir totalmente las relaciones entre los subgrupos bicoloratum, romanai y perflavum, pero el agrupamiento ((bicoloratum, romanai) perflavum)) sigue siendo una hipótesis aceptable, ya que tal clado es una de las posibles descomposiciones de la trifurcación. La inclusión de S. ochraceum dentro de la politomía y no junto a S. dinellii, señala dos posibles situaciones: o bien, tal especie no pertenece a dinellii sino más bien a (bicoloratum, romanai) o los caracteres considerados no disciernen a dinellii como grupo.
Caracteres
No existe claridad en cuanto a la delimitación de las homologías para cada taxón y todos los caracteres que eventualmente pueden definir grupos se presentan en otros items. Así, los grupos están definidos por combinaciones de caracteres.
Al comparar los caracteres considerados por Coscarón (1984, 1987, 1990), frente a los presentados en este trabajo, y dentro de un contexto totalmente distinto, se presenta una diferencia de caracteres. Este estudio se apoya tanto en datos morfométricos como en cualidades, mientras que Coscarón se apoya más en los últimos. La controversia es el resultado de incluir una nueva serie de datos y reconsiderar los datos anteriores. Puede y de hecho se da un cambio en la estructura de las sinapomorfias que en vez de rechazar la hipótesis original de monofilia de Ectemnaspis - Psilopelmia, refina los límites de definición de los grupos planteados.
Coscarón (1984, 1987, 1990) ha dado siempre mayor preponderancia al carácter base del cibario, su división en subgrupos se da por las distintas modificaciones que presenta el área basal, y contempla al carácter como una serie binaria de caracteres, mientras que en este trabajo se considera como un solo carácter multiestado.
La fragmentación o no fragmentación de un carácter multiestado en varios caracteres binarios excluyentes entre sí, modifica el contexto. Según el tipo de partición se puede generar un cierto peso sobre una estructura que, al no fragmentarse, no lo tendría; además, son dos marcos de referencia totalmente opuestos, pues la partición binaria excluyente obliga a ver la naturaleza como estructuras binarias, mientras que la posición en este trabajo es que se pueden dar múltiples casos de estructuras multiestado.
Obviamente, la ordenación de estos caracteres multiestado es y deber seguir siendo no aditiva, y evitar en lo posible la optimización, que indica más un problema de reconsideración frente al árbol que una realidad de la naturaleza (ver Hauser y Presch (1991) y Hauser (1992) a favor de los caracteres multiestado como no ordenados; en contra Wilkinson (1992)).
La prueba de permutación por colas (ptp<0,05) indica que para el árbol obtenido aunque se presentan valores bajos de Indice de consistencia e Indice de retención, este es el árbol más adecuado para los datos morfológicos contemplados.
No existen árboles más cortos logrados al azar. Como lo muestra la forma de la curva obtenida, cualquier modificación de los datos genera árboles más largos.
La prueba de ptp indica que los caracteres a pesar de mostrar gran cantidad de ruido y no delimitar con claridad los grupos, son de hecho informativos, pero con una definición diferente que la expuesta por Goloboff (1991). En lo que puede ser contemplado como una paradoja, el árbol exhibe estructuración cladística, pero dentro del cual existe gran nivel de ruido.
Taxa a partir de datos morfológicos
La agrupación expresada a partir de los datos morfológicos refuerza los trabajos anteriores (Coscarón 1990; Miranda 1991; Miranda y Muñoz de Hoyos 1993). El cladograma muestra, en primer lugar, que los taxa romanai y bicoloratum, percibidos como grupos, presentan una apropiada definición. Los resultados del análisis de filogenia a partir de datos morfológicos apuntan a su inclusión en la misma unidad jerárquica, aspecto que no está acorde con la evidencia cromosómica.
Los grupos perflavum y dinellii deben ser considerados como no Ectemnaspis. Tal posición es preferida a la expuesta por Coscarón (1990), porque aunque son taxa monofiléticos se maximizan las hipótesis de grupo al considerar en forma independiente a romanai - bicoloratum y no a los cuatro elementos en conjunto. Este aspecto también es notorio al observar los valores del grupo romanai - bicoloratum, frente a los de los cuatro grupos (Fig. 2-4).
Fenograma de distancia para los grupos considerados.
Dinellii, representada en este caso por S. dinellii - S. ochraceum - S. bipunctatum, no forma una unidad monofilética de por sí, como lo es S. lewisi - S. romanai; S. bipunctatum, según los datos, es colocado en un grupo monofilético con S. escomeli - S. quadrivittatum, lo que indica su pertenencia a Psilopelmia y no a Ectemnaspis.
Debido a que se presentan algunos vacíos en la información de S. quadrivittatum, su posición puede verse alterada al reemplazar los faltantes por su respectivo valor (Platnick et al. 1991).
Perflavum, que en este trabajo está caracterizada por una sola especie, se presenta como grupo separado y basal a romanai - bicoloratum. Esta ramificación de perflavum es sustentada igualmente por la evidencia biogeográfica, ya que es un grupo guyano - brasilero, mientras que los otros tres grupos son andinos (Coscarón 1990). Esta disyunción, que puede ser tipo trans-/cis- andina, es coincidente con la presentada por algunas especies de mosquitos (Conn et al. En prensa) y de aves (Haffer 1969; Simpson y Haffer 1978; Cracraft y Prum 1988).
Las afinidades que originalmente planteaba Crosskey (1987) no se deben ni a similitud ni a relación de parentesco, por lo tanto su clasificación no sería acertada ni desde una base filogenética ni desde una fenética.
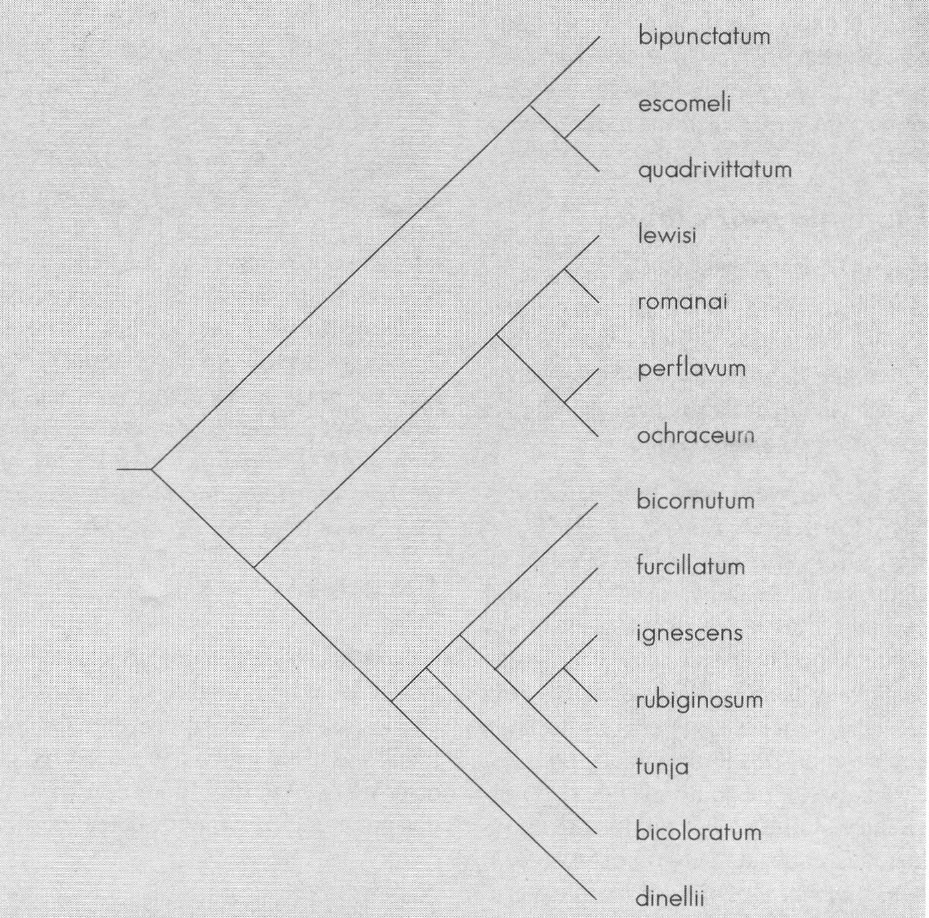
Como lo muestra la Figura 6, Psilopelmia estaría conformada por S. bipunctatum, S. escomeli y S. quadrivittatum, y a Ectemnaspis lo conformarían las especies S. bicornutum, S. furcillatum, S. ignescens, S. rubiginosum, S. tunja, S. bicoloratum y S. dinellii. Las otras especies: S. lewisi, S. romanai, S. perflavum y S. ochraceum pertenecerían a otra unidad no Ectemnaspis ni Psilopelmia.
De otro lado, ya que Crosskey sustenta la idea que perflavum y dinellii están más relacionados a S. escomeli que a S. bicoloratum, queda la discusión de medir la mayor o menor relación entre especies y Crosskey no ha propuesto ningún parámetro o valor para tal vínculo.
Cualquiera que sea la lectura de la figura, no es posible soportar los límites entre Ectemnaspis y Psilopelmia de Crosskey (1987). Este aspecto es más fácilmente evidenciable al advertir que dos especies tan ligadas como S.lewisi y S. romanai son colocadas en grupos diferentes. Así se muestra el riesgo eventual de asimilar los grupos según el criterio del investigador, sin validar los caracteres definitorios y la posible pertenencia a otras unidades.
La mejor interpretación, a la luz de los resultados, es circunscribir el subgénero Ectemnaspis al grupo bicoloratum, lo cual presenta mayor soporte no sólo con esta serie de datos sino también a partir de los obtenidos por Coscarón (1987) y Coscarón et al. (En prensa). Este taxa es claramente diferenciable de romanai, de Psilopelmia (escomeli - quadrivittatum - bipunctatum) y de dinellii y perflavum.
Descripción cromosómica
A partir de las fotografías de los complementos cromosómicos se ubicaron los marcadores primarios y secundarios, y posteriormente se comparó la posición de los marcadores. Tal cotejo se reproduce en forma de idiograma comparativo, donde se muestran las secuencias compartidas (Fig. 7-9).

Idiograma comparativo del cromosoma I para las especies involucradas en el análisis cromosómico. BIC: S. bicornutum; FUR: S. furcillatum; TUN: S. tunja; IGN: S. ignescens; ROM: S. romanai; L-L: S. lewisi-lutzianum; OCH: S. ochraceum.
Las comparaciones entre las especies anteriormente citadas y las fotografías de los complementos de S. lewisi-lutzianum están dadas al nivel de resolución más bajo, debido a la calidad de las fotografías, pero que puede ser considerado como verdadero. Con respecto a S. ochraceum, aunque existe una buena fotografía de un complemento, fue necesario localizar en primera instancia los marcadores, para realizar la posterior comparación.
Filogenia cromosómica
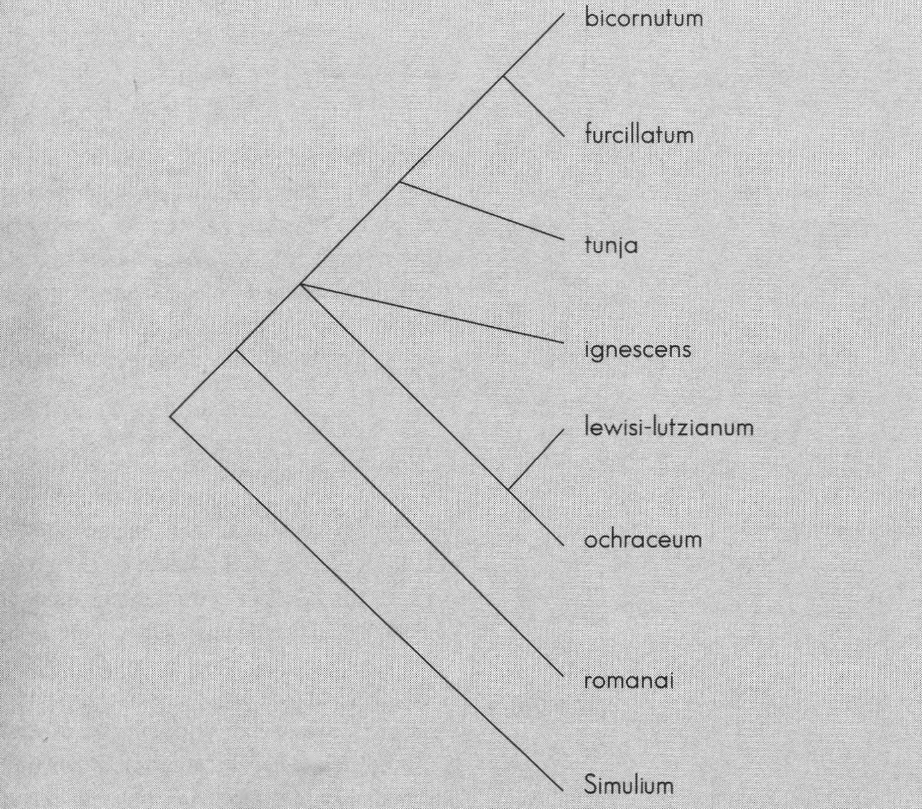
Dados los caracteres de los cromosomas (Fig. 7-9 y Tabla 3), se encuentran seis árboles posibles, cuyo consenso estricto se muestra en la Figura 10. Se muestran dos grupos claramente definidos:((S. bicornutum, S. furcillatum), S. tunja) y (S. ochraceum, S. lewisi-lutzianum); formando una politomía se encuentra S. ignescens y basal a este subclado se ubica S. romanai.
El grafo señala que los datos cromosómicos no soportan la agrupación morfológica, ya que es necesario incluir dentro de bicoloratum a los grupos dinellii y romanai.
A nivel cromosómico no existe evidencia para todos los subgrupos. Según trabajos anteriores, bicoloratum es definido por la homosecuencialidad en el cromosoma I y III, con rearreglos en el cromosoma II (Miranda 1991; Miranda y Muñoz de Hoyos 1993).
La inexistencia de duplicados para el grupo dinellii y las dificultades con la determinación del material de S. lewisi-lutzianum imposibilitan establecer las sinapomorfias cromosómicas para tales subgrupos y clarificar la trifurcación.

Idiograma comparativo del cromosoma II para las especies involucradas en el análisis cromosómico. BIC: S. bicornutum; FUR:S. furcillatum; TUN: S. tunja; IGN: S. ignescens; ROM: S. romanai; L-L: S. lewisi-lutzianum; OCH: S. ochraceum.
Al existir cierto grado de homosecuencialidad en los cromosomas I y III, se puede proponer la hipótesis de que los miembros de perflavum o de Psilopelmia sensu lato presentan tal carácter; lo que no es predecible con los datos actuales es la extensión de la semejanza, aunque se supone que debe ser mayor entre los miembros de Ectemnaspis - Psilopelmia que con otros grupos.
Como lo indica el valor de ptp (<0,1), los datos cromosómicos están menos estructurados que los morfológicos, ya que se obtienen varios árboles (28 = 5,6%) por permutación al azar con igual longitud que el árbol de parsimonia (ptp = 0,056); el menor nivel de estructuración puede obedecer a que existe mayor nivel de homoplasia, por lo que se tienen permutaciones con igual longitud.
Otra posible explicación a las diferencias de estructuración entre los dos tipos de datos se da por el número de repeticiones. Los datos morfológicos poseen más items y caracteres, por lo que puede requerirse mayor número de permutaciones que para los datos cromosómicos con menor número de taxa y caracteres. Sin embargo, tal problema es difícil de abordar por simulación, debido al tiempo requerido para las pruebas de PTP, ya que el tiempo de ejecución se incrementa exponencialmente al aumentar el número de items y por lo tanto de caracteres.
Es posible que una comparación banda a banda de las especies involucradas genere otros resultados. Es obvio que poseer todo el material en vivo a la vez es difícil, por simples razones logísticas; además, tal tipo de examen requiere mapas cromosómicos de mayor definición que la obtenida por tinciones tradicionales como orceina o feulgen.
Evidencia total: datos morfológicos y cromosómicos en conjunto
Ya que los datos cromosómicos contradicen los agrupamientos basados en morfología, se unieron los dos tipos de datos para obtener un grafo conjunto.
El árbol de la Figura 11 indica que existe consistencia para la definición del subgrupo bicoloratum, el cuál es claramente delimitable. No existen límites definidos para romanai, y S. ochraceum forma una unidad con S. lewisi-lutzianum, la cual no es monofilética con S. romanai.
Debido a la contraposición entre los dos tipos de datos, se puede concebir que se presenta una taza diferencial de ruido sobre cada tipo de carácter, que resulta en distintos niveles de homoplasia en los caracteres, pero no es evidenciable cuál es la fuente que posee menos ruido o cuál es la fuente más exacta y precisa para la reconstrucción de la filogenia.
Dentro de la literatura reciente se pueden encontrar casos de total incongruencia entre dos tipos de caracteres (Shaffer et al. 1991), hasta casi una total congruencia (Vane-Wright et al. 1992; Brown et al. 1994). En ninguno de los dos casos anteriores se puede hablar de congruencia total como indicativo de mejor y la incongruencia como indicativo de peor. Distintas fuentes de datos relatan distintas historias, y éstas a su vez pueden ser o no congruentes entre sí. Es evidente que no existe la fuente ideal de información, o que de existir no se poseen las herramientas para decidir cuál de ellas es, de hecho se duda de la existencia de tal mecanismo de decisión (Kluge y Wolf 1993).
Como lo indica el valor de ptp (<0,05), el nivel de estructuración de las dos fuentes de datos es mayor que el de los datos cromosómicos e igual al de los datos morfológicos. Este fenómeno es esperable, ya que es un número relativamente pequeño de items y grande de caracteres, donde se espera que exista menos distorsión por cualquier cambio introducido.
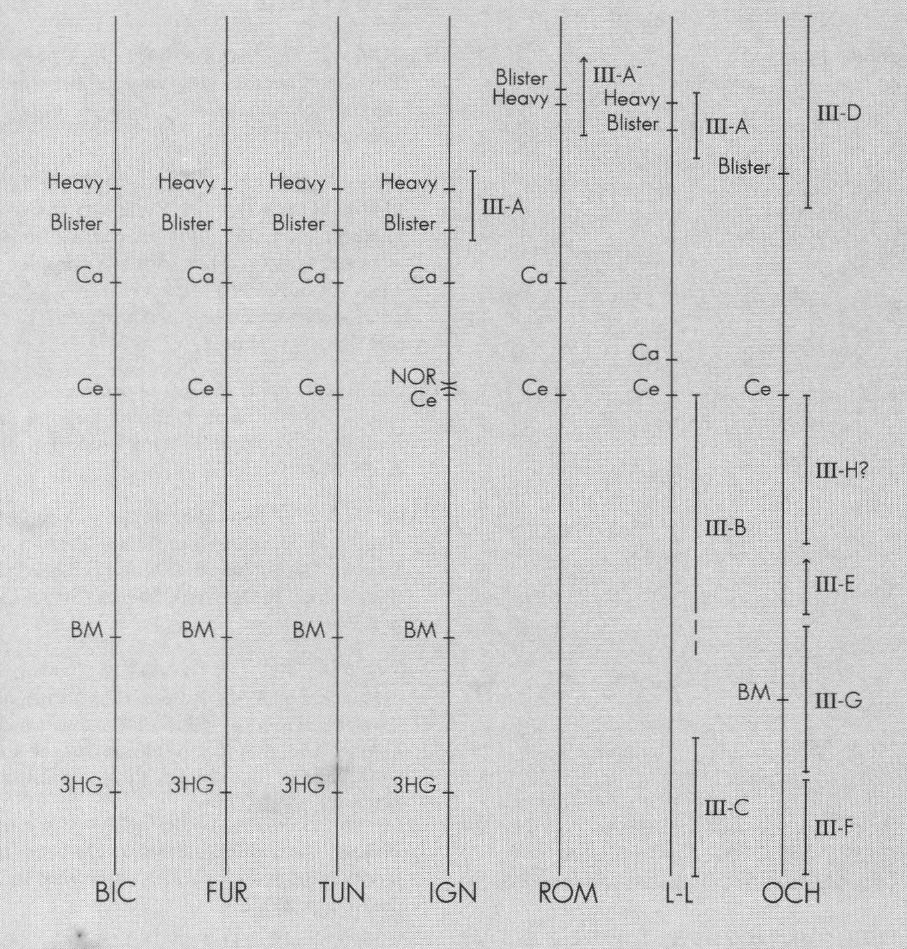
Idiograma comparativo del cromosoma III para las especies involucradas en el análisis cromosómico. BIC: S. bicornutum; FUR: S. furcillatum; TUN: S. tunja; IGN: S. ignescens; ROM: S. romanai; L-L: S. lewisi-lutzianum; OCH: S. ochraceum.
Nombres
El reto de este trabajo es determinar no sólo la pertenencia de las distintas especies a un subgénero dado sino también proponer la situación denominativa en que queda el grupo.
Como lo indican los resultados (Fig. 2-11), se plantean al menos cinco unidades: romanai, bicoloratum, dinellii, perflavum y Psilopelmia.
Para hacer coherente la estructura nominativa con la filogenia, el antes denominado subgrupo bicoloratum conformaría el subgénero Ectemnaspis, este grupo es definible tanto morfológica como citológicamente (Coscarón 1984, 1990; Miranda 1991; Miranda y Muñoz de Hoyos 1993).
El grupo hermano de bicoloratum es romanai, que debe ser reanalizado, ya que la evidencia cromosómica indica que quizá debe contener algunos elementos de dinellii. Este item podría ser incluido en Ectemnaspis según los datos morfológicos, pero no existe respaldo para tal idea al involucrar los datos cromosómicos solos o junto a los datos morfológicos.
Al convertir a bicoloratum en el subgénero Ectemnaspis, los subgrupos romanai, dinellii y perflavum adquieren rango subgenérico, y Psilopelmia se conserva como subgénero.
La designación de grupo hermano para Ectemnaspis - Psilopelmia sensu lato es materia de discusión, la falta de material de S. oviedoi - S. rivasi obliga en primera instancia a mantener a oviedoi como tal. No se descarta que en el futuro las dos especies sean sinonimizadas e incluidas en Chirostilbia o quizá Simulium.
El nombre de oviedoi puede mantenerse si esto es necesario, pero no se recomienda su uso por dos razones, la primera de tipo nomenclatural, ya que no es clara la definición de grupo y la segunda que se supone una incierta afinidad entre oviedoi y Ectemnaspis - Psilopelmia sensu lato.
La propuesta de cambio de categoría puede ser contemplada como un aumento en las dificultades de la familia, pero a la luz de la evidencia acumulada, es dar coherencia filogenética y nomenclatural a un hecho que se ponía en evidencia desde los trabajos de Coscarón y Wygodzinsky (1972). Esta reorganización de la nomenclatura, basada en la filogenia, se advierte igualmente en el trabajo de Py-Daniel (1990), quien propone, entre otras cosas, elevar a género a Simulium sensu stricto que automáticamente elevaría a género a todos los actuales subgéneros y que además implicaría la misma posición dentro de la jerarquía linneana para los grupos que plantea Coscarón (1987).
Perspectivas
Aunque el presente trabajo clarifica muchos de los problemas de nominación a nivel de Ectemnaspis/Psilopelmia, quedan muchos dilemas por resolver. Existe un dominio específico con respecto a S. romanai - S. lewisi y S. lutzianum que debe ser esclarecido desde la doble perspectiva morfología y cromosomas, cubriendo todo el rango de distribución de las especies para evaluar no sólo los extremos, sino también las áreas de simpatría. De otro lado, está por evaluar dentro de S. oviedoi- S. rivasi la validez de estas dos especies y su posición supraespecífica.
Debido a los planteamientos de Py-Daniel (1990), es posible que se haga necesario reconsiderar las jerarquías de subfamilia y tribu dentro de la familia, además de establecer si es necesario elevar a género los actuales subgéneros.
Cladograma (datos cromosómicos) para los grupos considerados. Consenso estricto de 6 árboles. longitud=28. Indice de consistencia= 0,82. Indice de retención= 0,64 (ptp<0,1).
Se han sugerido, igualmente, relaciones entre subgéneros neotropicales y africanos (Crosskey 1990); tal escenario se debe evaluar sobre la filogenia entre subgéneros que clarifique la posición de los grupos en el neotrópico frente a Australia y Africa.
Ante lo anterior es obvio que aunque se resuelva un problema quedan a su vez muchos interrogantes por contestar. La sistemática de simulíidos es un campo sugestivo como modelo de investigación, que transciende los linderos de lo nomenclativo y se adentra en lo biogeográfico, ecológico y de asociación. Es claro que más adelante las hipótesis de parentesco permitirán realizar estudios de biodiversidad, al contraponer al cladograma las características ecológicas de estos organismos dentro de la reciente visión de la ecología histórica (Brooks y McLennan 1991), además de permitir análisis coevolutivos, cuando se conozca la biología de los patógenos y la sistemática de esta asociación.
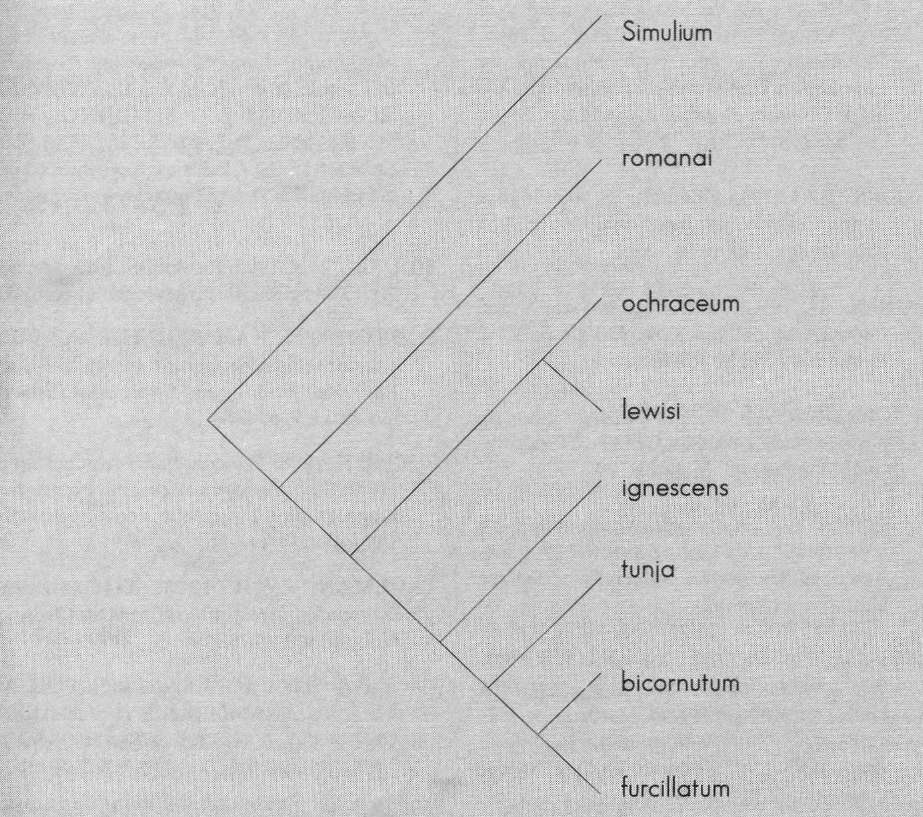
Cladograma (datos cromosómicos morfológicos) para los grupos considerados. longitud=204. Indice de consistencia= 0,76. Indice de retención= 0,54 (ptp<0,05).
Footnotes
Agradecimientos
Los autores desean expresar sus sentimientos de gratitud al Dr. Sixto Coscarón, quien permitió el acceso a la colección del Museo de La Plata durante la pasantía del primer autor, y además accedió al uso de información aún no publicada. Al Dr. A.J. Shelley, quien facilitó copias de los mapas cromosómicos de S. lutzinanum-lewisi, así como una versión de su trabajo sobre simulíidos del Ecuador. El Dr. M. Dallwitz facilitó una copia del programa DELTA. La bióloga Libia Mireya Hernández contribuyó con sus comentarios en la elaboración del manuscrito, en el acceso al programa RANDOM CLADISTICS y en la conversión a formato digital de las gráficas originales realizadas por Freddy Castellanos.
Este trabajo ha sido financiado por COLCIENCIAS dentro del proyecto «Estudio sistemático de los simúlidos de Colombia: transecto Bogotá - Honda.» (Proyecto 1101-05-037-92).