
Abstract
The present communication reports the dose and duration dependent toxicity of a PCB, Aroclor 1254, to a few ion dependent ATPases, Acid phosphatase, Alkaline phosphatase and Glucose-6-phosphatase in the whole brain tissue of mice. Two groups of mice were subjected to two sublethal doses (0.1 and 1 mg kgbw−1 day−1) of PCB orally and exposed for 4, 8 or 12 days. A separate control group received the corn oil vehicle for the same exposure times. The observed results indicated exposure duration dependent changes in the enzymatic levels in the brain. The results suggest that the alteration in the enzymatic activity was possibly due to imposed oxidative stress generated by Aroclor 1254 on membrane-bound ion-dependent ATPases and other phosphatases in the brain tissue.
1. INTRODUCTION
PCBs are persistent in the ecosystem, and are bioconcentrated through the food chain due to their affinity for lipids and resistance to metabolism. In the environment, the behavior of PCB mixtures is directly correlated to chlorination. Aroclor is strongly absorbed into soil, remains immobile when leached and mixtures of Aroclor are reported to be highly mobile in the presence of organic solvents (USAF 1989). Oral LD50 values for Aroclor 1254 are reported to be 1295 mg/kg/day in male Osborne-Mendel rats, 1010 mg/kg/day in Sherman rats and 4000 mg/kg/day in the mink (ATSDR 1995). It was reported that rats administered with sublethal dose of Aroclor 1254 in the diet for 4 days exhibited increased relative liver weight and increased serum cholesterol (Carter 1984, 1985). Hepatic vacuolar degeneration and a 30% decrease in body weight gain were observed in rats administered sublethal doses of Aroclor 1254 in the diet for four days (Carter and Koo 1984). Price et al. (1988) reported that rats exposed to 7.5 mg/kg/day dose of Aroclor 1254 in the diet for 7 days showed increased relative liver weight, decreased liver glucose 6-phosphatase activity, and decreased serum T4 hormone levels. Whereas, in mice fed 130 mg/kg/day Aroclor 1254 in the diet for 2 weeks, a 10-fold increase in serum corticosterone and a 2-fold increase in relative adrenal weight were observed (Sanders et al. 1974).
PCBs are known to have carcinogenic, hepato- and immunotoxic activities and exhibit inhibitory effects on the enzymatic systems investigated in laboratory animals, birds and fish (Perez-Lopez et al. 2002). The neurotoxic effects of Aroclor 1254 include disturbance of intracellular signaling or second messenger homeostasis (Lynch 1998). Effects of PCBs on rats also include diarrhea, breathing difficulties, dehydration, decreased response to pain and coma. PCBs reportedly cause considerable damage to the lungs, stomach, and pancreas in mammals (Kodavanti 2005). The activity of Na+, K+-ATPase and Mg2+ ATPase from several fish tissues were reported to be inhibited by PCBs (Yap 1971; Cutkomp 1972; Desaiah 1972; Koch 1972). It was reported that the administration of commercial PCBs, as well as purified isomers to rats inhibited the total ATPase activity in the liver, kidney and brain tissues (LaRocca and Carlson 1975; Hale et al. 2003). LaRocca and Carlson (1979) have evaluated the relationship between the inhibition of Mg-ATPase and the lipophilic property of PCBs and reported a strong negative correlation between PCB-induced inhibition of ATPases and solubility of the PCBs. Nishihara and Utsumi (1985) reported that ortho-substituted PCBs alter calcium homeostasis by inducing changes in mitochondrial membrane integrity. The present communication reports the sublethal dose and duration dependent in vivo toxicity of PCB (Aroclor 1254) to some membrane-bound ion dependent ATPases and other phosphatases in the whole brain tissue of adult male Swiss albino mice.
2. MATERIALS AND METHODS
2.1 Chemicals
Aroclor 1254 was procured from Sigma-Aldrich Chemicals Ltd. (CAS No. 11097-69-1). All the other chemicals used for this study were of analytical grade.
2.2 Animals
Adult male Swiss albino mice, around 2 months of age and weighing around 30–40g, were used for the study. The animal groups were provided ad libitum rodent diet and water, kept under prescribed standard conditions and all experiments were conducted according to the ethical norms approved by the CPCSEA, India.
2.3 Experimental design and Enzyme assays
A total of 27 adult male Swiss albino mice were used for the entire study. Different groups of mice were given an oral administration of Aroclor 1254 (0.1 and 1 mg/kg body weight /day) dissolved in corn oil for three different exposure periods of 4, 8 and 12 days. After completion of toxic exposure, the whole brain tissue was quickly dissected out and washed with ice cold distilled water to free it from blood and other tissues. The complete brain was homogenized in a Potter-Elvehjem glass homogenizer in sucrose-EDTA-imidazole buffer at 0 – 4° C to make a 10% (w/v) tissue concentration. Activities of ATPases viz. total-, Na+, K+-, Ca2+- and Mg2+- ATPases were estimated as per the method of Zaugg (1982) with appropriate modifications (Lakshmi et al. 1991). Activity of Glucose-6-phosphatase was estimated by the method of Shimeno (1982) and inorganic phosphate was measured by the method of Fiske and Subbarow (1925). Activities of alkaline phosphatase (ALP) and acid phosphatase (ACP) were measured by the method of Dasgupta and Ghosh (1993). Protein was estimated by the method of Lowry et al. (1951) using crystalline bovine serum albumin as a standard.
2.4 Statistical analyses
The obtained data were analyzed by a two-factor ANOVA, single-factor ANOVA and Student's t-test. All statistical procedures were computed as per Sokal and Rohlf (1969).
3. RESULTS
Results of the present investigation showed variations in the toxicity of Aroclor 1254 in different doses and exposure duration groups. In the 0.1 mg/kg/day dose group of PCB, the activity of total ATPase was slightly increased in all the exposure durations as compared to control but in case of the 1 mg/kg/day dose group, the enzyme activity was highly inhibited after 4 days of exposure but slightly inhibited after 8 days of exposure (Fig. 1a). Activity of Na+, K+- ATPase showed slight stimulation in all exposure durations in the 0.1 mg/kg/day dose group, whereas, in 1 mg/kg/day dose group, activity was highly inhibited after 4 days of exposure (Fig. 1b). Ca2+-ATPase activity was stimulated in the 0.1 mg/kg/day group after 4 and 8 days of exposure while, slight inhibition of activity was observed after 12 days of exposure. In the brain of the animals exposed to 1 mg/kg/day dose, the activity of Ca2+- ATPase was inhibited after 4 days of exposure, but showed slight stimulation in 8 days of exposure (Fig. 1c). Specific activity of Mg2+- ATPase showed inhibition at the lower dose after 4 days but was stimulated after 8 and 12 days of exposure. In the higher dose group, the activity of this enzyme was slightly stimulated in all exposure periods (Fig. 1d). Glucose-6-phosphatase levels showed a stimulatory effect in the lower dose group after 4 days of exposure, while inhibitory effects were detected in the higher dose group after 8 and 12 days of exposure (Fig. 2a). An inhibitory trend was observed in the higher dose group for alkaline phosphatase activity while the lower dose group showed stimulation after 4 and 12 days of exposure and inhibition after 8 days of exposure (Fig. 2b). In the lower dose group, acid phosphatase levels were slightly stimulated after 4 days of exposure and strongly stimulated after 12 days of exposure, but inhibited after 8 days of exposure. However, in higher dose group, an inhibitory trend was observed in levels of acid phosphatase.(Fig. 2c).
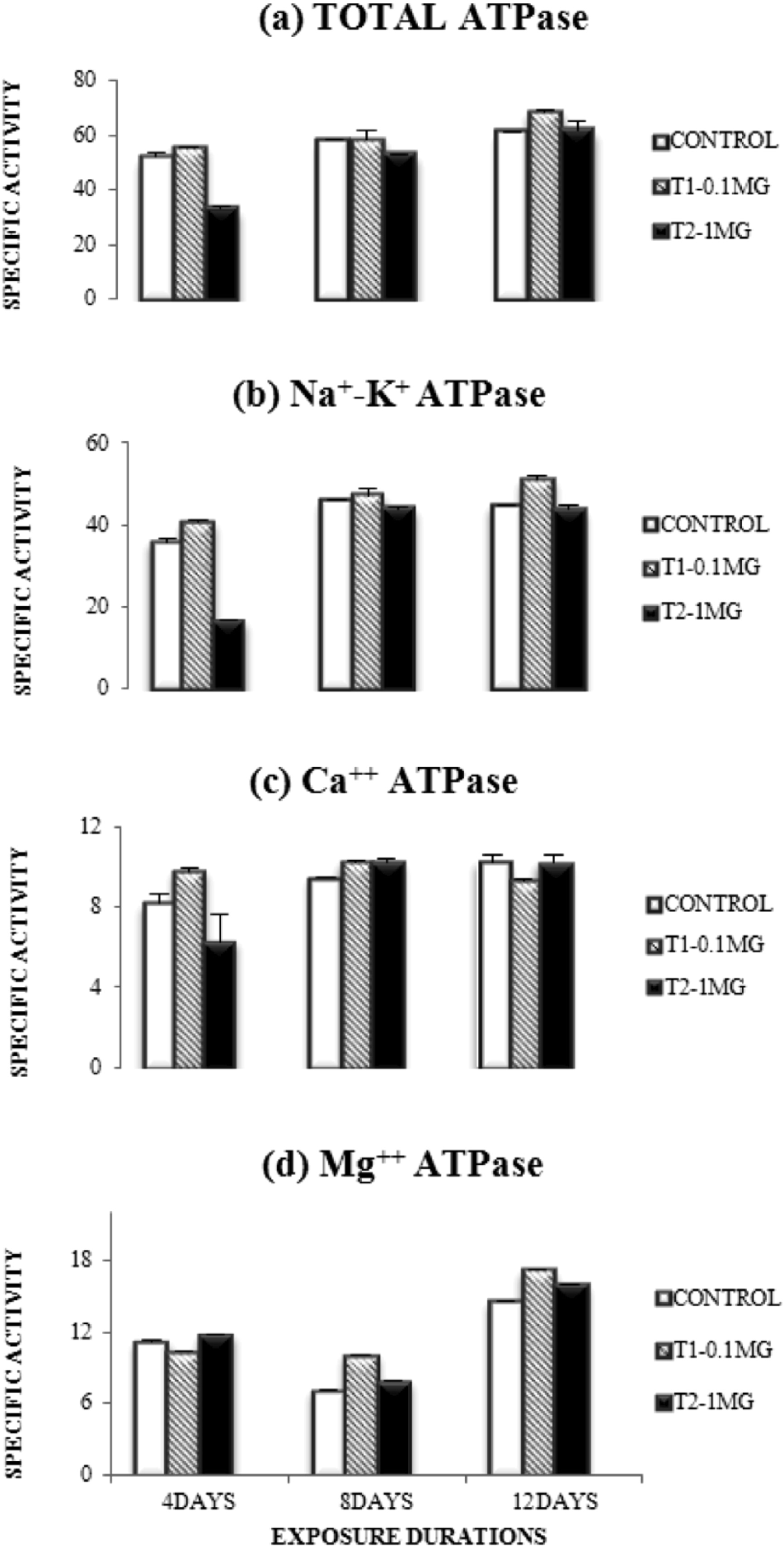
Dose and duration dependent changes in the specific activities of different ATPases after Aroclor 1254 intoxication.

Dose and duration dependent changes in the specific activities of different phosphatases after Aroclor 1254 intoxication.
4. DISCUSSION
The result of the present study revealed an exposure duration dependent effects of the Aroclor 1254 on the enzymes studied. This is possibly due to the high lipid solubility of PCB by which it is readily diffused through the membrane (Kodavanti 2005). The predominant trend of enzymes stimulation in most of the cases was possibly due to exposure duration dependent effects. Disruption of membrane structures can cause changes in ion permeability through voltage or ligand gated channels as well as changes in activity of membrane bound enzymes (Tan et al. 2004). The disruption of both plasma membrane integrity and intracellular organelles suggested that PCB may be altering the membrane structure and calcium homeostasis by inducing the changes in mitochondrial membrane structure (Nishihara and Utsumi 1985). ATPases are known to regulate different ions and metabolites across the membrane, in order to maintain the physiological requirements of the cell. It may be possible that the observed disturbance of Na+, K+- ATPase affected the Na+, K+ pump, causing uncontrollable entry of Na+ into the cell along the concentration gradient. As a result of this, the water molecules followed along the osmotic gradient (Thaker et al. 1996) caused swelling of the cell and membrane rupture (Kundu et al. 1992). It is also possible that the enzyme might have accumulated in the intracellular space due to altered membrane permeability or leakage in the affected membrane (Suzuki 1988). Alkaline phosphatase is a membrane bound enzyme responsible for the movement of metabolites across the membrane. Thus, the disturbance of the activity of this enzyme indicates impaired nutrient assimilation and absorption (Hossain and Dutta 1986; O'Brien et al. 1989). Several studies showed that the alterations in the membrane transport mechanisms and neurotransmitters appear sensitive to PCBs possibly caused by the formation of reactive oxygen species (Kodavanti 2005). These results suggest that sublethal PCB concentrations may be activating an oxidative stress-related pathway, which potentially disrupts neuron function. The results of two-factor ANOVA showed exposure duration dependent effects of PCBs in the brain tissue (Table 1), however, the t-test performed between the control and treatments showed variations in different enzymes studied (Table 2). For total ATPase, significant differences were observed in 4 days (2.68, p = 0.05) and 12 days (6.19, p = 0.05) exposure treatment groups between control and lowest dose. However, high statistical significance was observed in the highest dose group in all other enzymes studied except Na+, K+-ATPase and Mg2+-ATPase (Table 2). In case of the Na+, K+-ATPase, the results of the t-test conducted between control and treatments showed significant differences between control and most of the doses exposed for different exposure durations (6.67 in 0.1 mg, 4 days; 14.30 in .01 mg, 12 days; 36.02 in 1 mg, 4 days and 5.30 in 1 mg, 8 days; p = 0.05) (Table 2). The activity of Ca2+-ATPase, Mg2+- ATPase, alkaline phosphatase and glucose-6-phosphatase in the lower dose group showed less variation from the controls in most of the exposure treatment groups. (Table 2). This trend is indicative of marginally significant (2.97, p = 0.05) dose dependent effects on the enzymes in brain tissue. The result of single-factor ANOVA which indicates the variations between exposure treatment groups within each dose indicated highly significant variations (76.19, 46.03; p = 0.05) between the exposure durations in most of the enzymes studied except acid phosphatase (Table 3).
Results of Two-factor ANOVA between control and toxicated groups.

Significant at P = 0.05 (F crit (df=3, 8) = 3.01)
Significant at P = 0.05 (F crit (df=8, 35) =2.36)
Results of Student's ‘t’-test between control and individual exposure durations within each dose.

Significance at P = 0.05 (T crit. = 2.447).
Results of single-factor ANOVA between individual exposure durations within each group.

Significance at P = 0.05 (F crit. = 4.26).
In conclusion, it can be stated that the result of the present study revealed a predominantly exposure duration dependent effects of the Aroclor 1254 on the enzymes studied. At this point of the study, the reasons for lesser magnitude of dose dependent effects of the toxicant is not clear, however, the damage caused by exposure durations is clearly indicative of indirect effect of PCB to these enzyme systems in in vivo conditions.
Footnotes
5. ACKNOWLEDGMENTS
Authors are thankful to UGC, New Delhi and Govt. of Gujarat (Research Oriented Programme in Marine Biology) for financial support. UGC, Govt. of India is also thankfully acknowledged for a Meritorious Fellowship awarded to the senior author.