
Abstract
Transgenic animals bearing the reporter gene, LacZ, encoding the histochemical enzyme, β-galactosidase, are increasingly becoming available. Similarly, antibody conjugates consisting of specific IgGs coupled to horseradish peroxidase (HRP) are widely used for Western blotting, ELISA, and immunohistochemistry. Here we provide a detailed fixation and histochemical protocol for the simultaneous electron microscopic visualization and discrimination of β-galactosidase and peroxidase reaction products within mouse kidney. After incubation of transgenic LacZ tissues with IgG-HRP conjugates, samples were lightly fixed with 2% paraformaldehyde and 0.4% glutaraldehyde and processed for peroxidase histochemistry. Tissues underwent β-galactosidase histochemistry, were refixed with glutaraldehyde, osmicated, and embedded. In Flk1/LacZ mice, we immunolocalized anti-laminin β1 chain IgG-HRP specifically to developing glomerular basement membranes, whereas Flk1/LacZ was expressed only by glomerular endothelial cells. In Epas1/LacZ mice, we immunolocalized anti-platelet endothelial cell adhesion molecule-1 specifically to glomerular endothelial plasma membranes, whereas Epas1/LacZ was expressed by both glomerular endothelial and mesangial cells. This dual ultrastructural localization technique should be broadly applicable for immunoelectron microscopic studies in LacZ transgenic animals, particularly those where LacZ expression and antibody-HRP binding are both relatively abundant.
Keywords
D
With the advent of inducible and cell-selective transgene expression technologies, certain experiments in transgenics may require cellular identification of reporter gene expression by electron microscopy. These studies may also present opportunities for immunoelectron microscopy, and there may be instances where the dual localization of β-galactosidase expression and immunoperoxidase labeling are desirable. Here we provide a detailed protocol for the simultaneous ultrastructural localization of β-galactosidase and immunoperoxidase in kidneys of LacZ transgenic mice. In developing this combined β-galactosidase and peroxidase technique, we needed outcomes in which both the transgene expression and antibody distribution patterns were already well understood. Therefore, we carried out immunolocalization of laminin β1 chain within the developing glomerular basement membranes (GBMs) of Flk1/LacZ mice, which express LacZ specifically on endothelial cells (Robert et al. 1998). Flk1 [also referred to as vascular endothelial growth factor (VEGF) receptor 2] is a receptor tyrosine kinase that binds VEGF, is found on hemangioblasts and endothelial cells and is needed for the development of the vascular system (Shalaby et al. 1995). Laminin β1 is a member of the laminin α1β1γ1 heterotrimer (LM-111), which is present transiently within GBMs of immature glomeruli (Abrahamson and St. John 1992) and replaced by LM-521 during glomerular maturation (Miner 2005). In separate experiments, we immunolocalized platelet endothelial cell adhesion molecule-1 (PECAM-1; CD31) on plasma membranes of glomerular endothelial cells of Epas1/LacZ transgenic mice. Epas1 (also known as HIF2α), is a member of the hypoxia-inducible family of transcription factors important for the expression of gene products necessary to survive hypoxic stress (such as erythropoietin) and for vascular development (such as VEGF) (Wenger et al. 2005; Haase 2006). We recently showed in the kidney that Epas1/LacZ is expressed chiefly by endothelium and vascular smooth muscle, including glomerular mesangial cells (Steenhard et al. 2007). PECAM-1 is a plasma membrane cell adhesion molecule expressed only by platelets, monocytes, neutrophils, selected T cells, and endothelial cells (Scholz and Schaper 1997; Ilan and Madri 2003). For both experiments, we simultaneously localized β-galactosidase expression and bound antibody-HRP conjugates to the appropriate ultrastructural locations, and there was no ectopic deposition of reaction product for either enzyme.
Materials and Methods
Reagents
HRP (type VI) and DAB were purchased from Sigma Chemical Co. (St. Louis, MO), and Bluo-gal was obtained from Invitrogen Life Technologies (Carlsbad, CA). Rat anti-mouse laminin monoclonal IgG (MAb 5A2) was purified and characterized as previously described (Abrahamson et al. 1989). Rat monoclonal anti-PECAM-1 IgG was purchased from BD Pharmingen (clone MEC 13.3; BD Biosciences, San Jose, CA). Both antibodies were conjugated directly to activated HRP (Nakane and Kawaoi 1974) and dialyzed into PBS.
Transgenic Mice
Epas1tm 1 Rus mice were purchased from The Jackson Laboratory (Bar Harbor, ME). The colony was maintained by intercrossing heterozygotes (Tian et al 1998), and mice were genotyped as described previously (Steenhard et al. 2007). Flk1tm 1 Jrt mice also came from The Jackson Laboratory, and animals were genotyped as before (Robert et al. 1998).
Tissue Fixation
Animal experiments were approved by the Institutional Animal Care and Use Committee to ensure compliance with the Health Research Extension Act, Animal Welfare Act, and the Public Health Service Policy on Humane Care and Use of Laboratory Animals.
Two 5-day-old Flk1/LacZ heterozygous mice received intraperitoneal injections of 75 μl rat anti-mouse laminin MAb 5A2 IgG-HRP (1.1 mg/ml in PBS). Eighteen hours later, mice were killed by halothane inhalation and cervical dislocation, and the left kidneys were fixed in situ by the injection of a mixture of 2% paraformaldehyde and 0.4% glutaraldehyde in PBS into the cortex (0.05 ml of fixative was gently injected using a 1.0-ml syringe fitted with a 30-gauge needle). Fixative was injected while simultaneously clamping the renal artery, vein, and ureter at the hilus with a hemostat. Kidneys were removed 5 min later, and cortical tissue was trimmed into 2-mm cubes and placed into vials of fixative for 90 min on ice. Samples were washed with PBS (three exchanges for 5 min each) and stored in the same buffer at 4C. Similarly, two 4-week-old Epas1/LacZ heterozygous mice received intraperitoneal injections of 100 μl rat anti-PECAM-1 IgG-HRP (0.35 mg/ml). Four hours after injection, kidneys were fixed exactly as described above, except 0.1 ml of fixative was injected into kidneys of these older mice. Kidney tissues from uninjected, wildtype mice served as controls.
Histochemistry
Fixed kidney tissues from each animal were sliced into sections, 50 μm thick, using a Vibratome (Series 1000; Technical Products International, St. Louis, MO), and placed into vials of PBS. Sections were first incubated with 0.05% DAB in 0.1 M phosphate buffer, pH 6.0, for 15 min at room temperature, and then with 0.05% DAB plus 0.01% hydrogen peroxide (Graham and Karnovsky 1966) in the same buffer for 45 min at room temperature with gentle mixing on a rotating platform. After DAB incubation, samples were washed with three exchanges of 0.1 M phosphate buffer, pH 7.3 (5 min each). Next, samples were color developed for 4–6 hr at 37C with gentle mixing on a rotator in 20 mM potassium ferricyanide, 20 mM potassium ferrocyanide, 2 mM magnesium chloride, and 2 mM Bluo-gal (Weis et al. 1991). These color development reagents were prepared in 20 mM Tris, pH 7.6 (which drops to pH 7.3 at 37C), to minimize endogenous, mammalian β-galactosidase activity (Weiss et al. 1999). Tissues were washed three times with 0.1 M phosphate buffer, pH 7.3, and were refixed with 2% glutaraldehyde in 0.1 M phosphate buffer, pH 7.3, for 30 min on ice. Samples were washed with three 5-min exchanges of the same buffer and postfixed with 2% OsO4 in 0.1 M phosphate buffer, pH 7.3, for 90 min on ice. Next, samples were dehydrated through graded ethanol and propylene oxide and infiltrated overnight in 50:50 propylene oxide/Polybed 812 (Polysciences; Warrington, PA). Tissue slices were flat-embedded in the caps of inverted BEEM capsules containing fresh Polybed 812 and polymerized overnight at 60C. Care was taken to section the outermost surface of samples. Ultrathin sections were stained for 2 min with Reynolds lead citrate (Reynolds 1963) only and examined at 60 kV. Staining with uranyl acetate was omitted because it obscured the HRP reaction product.
Results
In this study, we developed a detailed protocol to carry out immunoelectron microscopic localization of IgG-HRP conjugates while simultaneously visualizing the expression of the LacZ gene product, β-galactosidase.
In infant Flk1/LacZ mice that received injections of anti-laminin β1 IgG-HRP, the β-galactosidase and peroxidase reaction products were distinctly different qualitatively and localized to discrete structures (Figure 1). Specifically, β-galactosidase produced a crystalline, extremely electron dense reaction product on intracellular and plasma membranes of kidney glomerular endothelial cells, confirming the endothelial expression of LacZ seen previously in tissue sections processed for light microscopy with X-gal (Robert et al. 1998). Occasionally, the β-galactosidase reaction product appeared to contain small cracks or tears (Figure 1) that were probably caused by generation of abundant reaction product at these sites and/or heat created during ultrathin sectioning. Importantly, the β-galactosidase reaction product was not observed on podocytes, which is also consistent with an absence of podocyte expression of Flk1/LacZ as seen by light microscopy (Robert et al. 1998). Additionally, LacZ expression was absent from glomerular mesangial cells, indicating that Flk1 is expressed exclusively by endothelial cells in kidney. In contrast, anti-laminin IgG-HRP localized only to the GBMs and mesangial matrices of developing glomeruli. This peroxidase-antibody conjugate produced a diffuse reaction product that appeared less electron dense and more uniform than the heavy, crystalline reaction product observed for β-galactosidase (Figure 1). The HRP reaction product was absent in endothelial cells, mesangial cells, and podocytes. Identical dual labeling results were obtained for both Flk1/LacZ mice examined. No β-galactosidase or peroxidase reaction products were observed in uninjected, wildtype mice that served as controls.
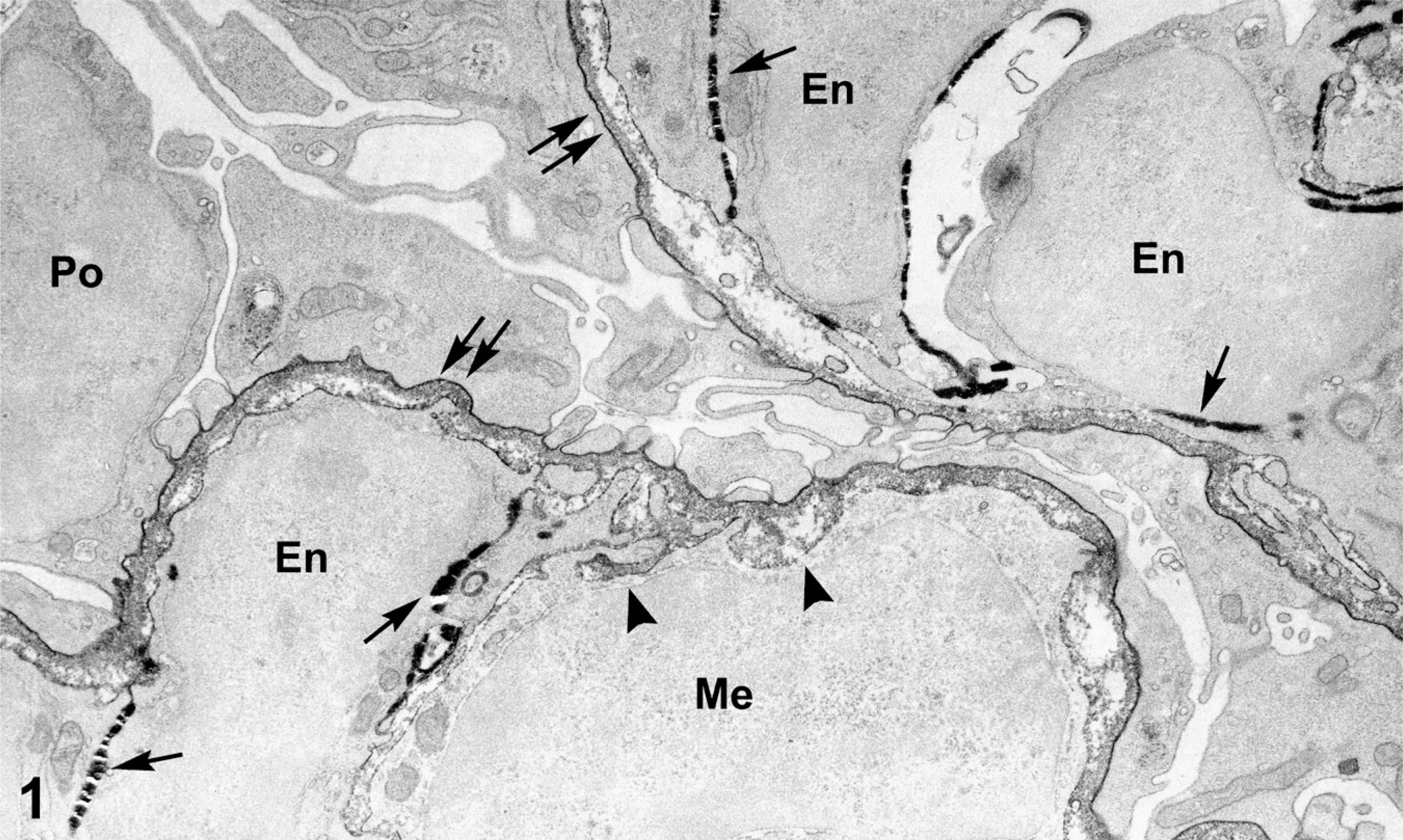
Electron micrograph showing portion of kidney glomerulus from a 5-day-old Flk1/LacZ heterozygous mouse that had received an intraperitoneal injection of anti-laminin β1 chain IgG-horseradish peroxidase (HRP). LacZ expression, reflected by the β-galactosidase reaction product (arrows), is seen as extremely electron dense deposits on plasma and intracellular membranes of endothelial cells only (En). Mesangial cells (Me) and podocytes (Po) do not express LacZ. Anti-laminin IgG-HRP generates a peroxidase reaction product within the glomerular basement membrane (GBM; double arrows) and mesangial matrices (arrowheads) of developing glomeruli. Note distinctly different appearances between the β-galactosidase (arrows) and peroxidase (double arrows) reaction products, respectively.
In 4-week-old Epas1/LacZ mice that received injections of anti-PECAM-1-HRP, the reaction products for β-galactosidase and peroxidase also differed (Figure 2). In this case, a heavy β-galactosidase reaction product occurred on membranous structures of both glomerular endothelial and mesangial cells, confirming expression of Epas/LacZ by these cells as observed before (Steenhard et al 2007). On the other hand, anti-PECAM IgG-HRP generated a less dense but more uniform labeling pattern specifically on plasma membranes of glomerular endothelial cells. These electron microscopic findings confirm evidence presented previously by light microscopic immunofluorescence (Ricono et al. 2003) and immunohistochemistry (Pusztaszeri et al. 2006) for the presence of PECAM-1 on glomerular endothelial cells. There was no anti-PECAM IgG-HRP labeling of mesangial cells or podocytes or within the GBMs (Figure 2). Identical labeling patterns were observed for both Epas1/LacZ mice.
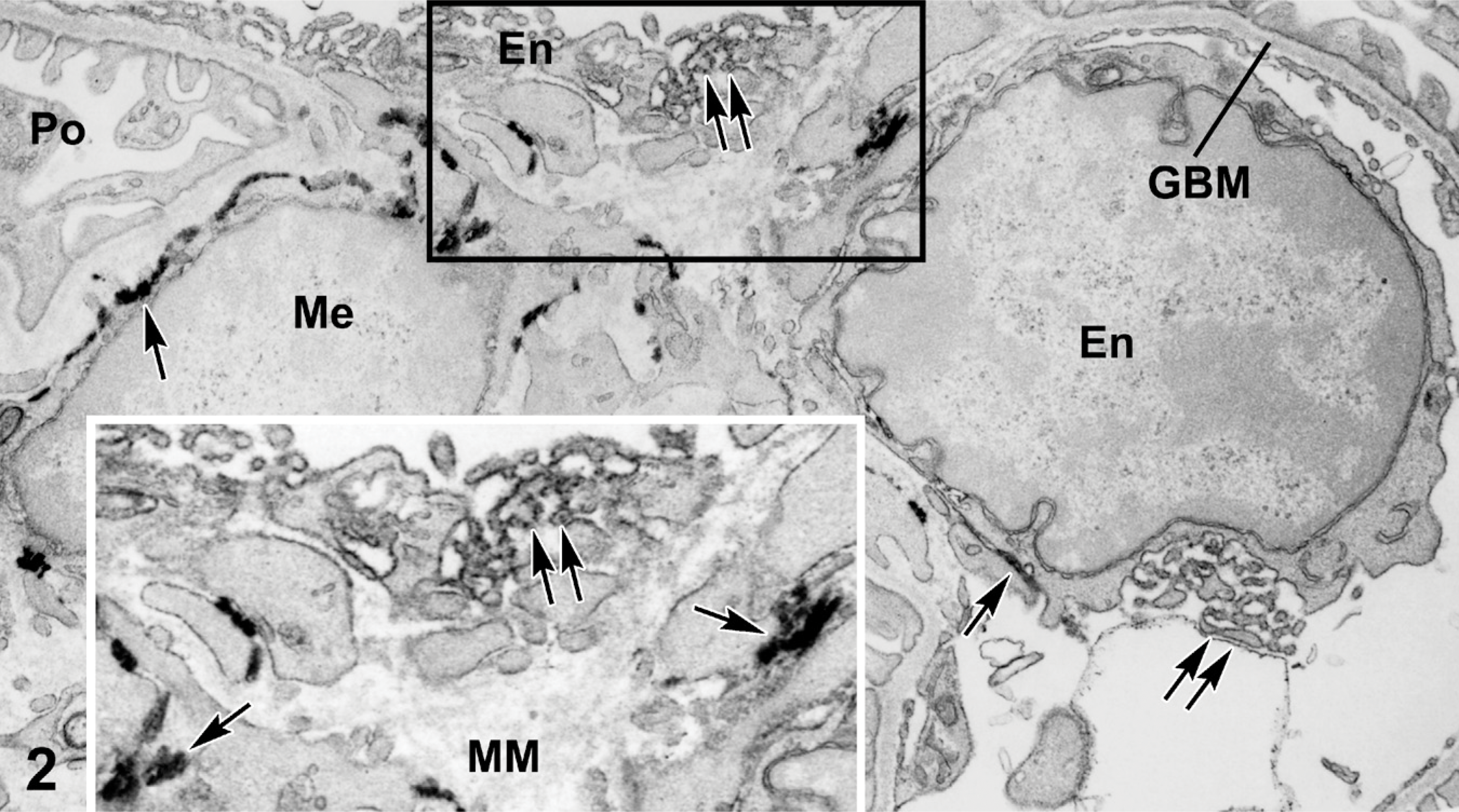
Portion of glomerulus from a 4-week-old Epas1/LacZ mouse that received an intraperitoneal injection of anti-platelet endothelial cell adhesion molecule-1 (PECAM-1) IgG-HRP. β-galactosidease reaction product is again viewed as extremely electron dense deposits on membranous structures (arrows) of endothelial cells (En) and mesangial cells (Me). Anti-PECAM IgG-HRP produces a qualitatively different and more diffuse peroxidase reaction product on plasma membranes of endothelial cells only (double arrows). GBMs, mesangial matrices (MM), and podocytes are not lableled by HRP or β-galactosidase. Boxed area is shown at higher magnification in the inset to show the qualitatively different reaction products for β-galactosidase and HRP.
Optimization of fixation conditions was critically important for the dual localization of β-galactosidase and HRP reaction products. Among various fixative concentrations tested, mixtures consisting of 2% paraformaldehyde and 0.4% glutaraldehyde preserved adequate cell ultrastructure and allowed sufficient enzyme activity. Higher concentrations of paraformaldehyde and glutaraldehyde resulted in much weaker and inconsistent dual labeling (data not shown). On the other hand, lower concentrations of aldehydes resulted in poor tissue morphology. Similarly, the sequence in which tissues were incubated with DAB and Bluo-gal substrates also affected results. Development of tissue for β-galactosidase histochemistry before peroxidase provided good β-galactosidase localization but weak or no labeling for HRP, possibly because of the prolonged incubation of tissue at 37C. To determine whether formation of DAB or Bluo-gal reaction products was compromised by performing the dual peroxidase and β-galactosidase histochemical reactions on the same tissue, separate Vibratome sections from anti-laminin β1 IgG-HRP-injected Flk1/LacZ mouse kidneys were processed for peroxidase histochemistry alone or Bluo-gal reaction alone. Slides from each individual reaction were compared with those that had undergone dual peroxidase and Bluo-gal development. In both cases, kidney tissue that underwent individual reactions contained similar amounts of DAB and Bluo-gal reaction products as those that had been serially reacted in the sequence described here. Additionally, ultrathin sections taken from the outermost surfaces of Vibratome sections produced the most consistent dual labeling. Sections taken from deeper regions within the blocks were weakly labeled, signifying the importance of thorough exposure of reactive tissues to the β-galactosidase and peroxidase substrates.
Discussion
Using the methods described here, we developed an approach for the dual ultrastructural localization of β-galactosidase and immunoperoxidase in LacZ transgenic mice. In the two separate cases presented, we could clearly distinguish the separate β-galactosidase and HRP reaction products in the electron microscope. Nevertheless, all of the usual caveats regarding tissue histochemistry and immunolocalization apply. Although the reaction product for β-galactosidase appeared similarly electron dense and in approximately equal amounts for both Flk1/LacZ and Epas1/LacZ mice, this will not occur for every transgenic mouse expressing the LacZ reporter. Indeed, the distribution and relative abundance of reaction product will likely vary depending on the cell expressing the transgenic reporter gene and the level of its transcription. Additionally, the ability to detect and discriminate an immunoperoxidase reaction product will certainly depend on the amount of bound antibody and its distribution. Moreover, in both cases described here, antibody-HRP conjugates were administered in vivo that bound to antigens accessible through the bloodstream. Whether this combined reporter gene/immunohistochemical approach will work for detection of cryptic, intracellular epitopes is therefore uncertain. On the other hand, successful immunolabeling of intracellular antigens can usually be achieved through the application of antibody conjugates to lightly fixed tissue sections, and the conditions used here may work satisfactorily for many antibodies.
Increasingly, new transgenic animals expressing the LacZ reporter gene are becoming available. Additionally, numerous antibody-HRP conjugates are available commercially or readily prepared in the laboratory as described here. In view of the versatility of immunoperoxidase techniques for immunolocalization, Western blotting, and ELISA, an ability to simultaneously localize immunoperoxidase in LacZ transgenics should therefore be advantageous. We conclude that, in cases where standard light microscopic approaches show that relatively high levels of LacZ expression and bound antibody are both present, the methods described here can be used reliably for the dual localization of LacZ transgenic cells and immunoperoxidase at the electron microscopic level.
Footnotes
Acknowledgments
This study was supported by National Institutes of Health Grants DK052483 and DK065123.
We thank Eileen Roach for help with the figures and Larysa Stroganova for genotyping.