
Abstract
During glomerular development, subendothelial and -epithelial basement membrane layers fuse to produce the glomerular basement membrane (GBM) shared by endothelial cells and epithelial podocytes. As glomeruli mature, additional basement membrane derived from podocytes is spliced into the fused GBM and loose mesangial matrices condense. The mechanisms for GBM fusion, splicing, and mesangial matrix condensation are not known but might involve intermolecular bond formation between matrix molecules. To test for laminin binding sites, we intravenously injected mouse laminin containing α1-, β1-, and γ1-chains into 2-day-old rats. Kidneys were immunolabeled for fluorescence and electron microscopy with domain-specific rat anti-mouse laminin monoclonal antibodies (MAbs), which recognized only mouse and not endogenous rat laminin. Intense labeling for injected laminin was found in mesangial matrices and weaker labeling was seen in GBMs of maturing glomeruli. These patterns persisted for at least 2 weeks after injection. In control newborns receiving sheep IgG, no binding of injected protein was observed and laminin did not bind adult rat glomeruli. To assess which molecular domains might mediate binding to immature glomeruli, three proteolytic laminin fragments were affinity-isolated by MAbs and injected into newborns. These failed to bind glomeruli, presumably owing to enzymatic digestion of binding domains. Alternatively, stable incorporation may require multivalent laminin binding. We conclude that laminin binding sites are transiently present in developing glomeruli and may be functionally important for GBM assembly and mesangial matrix condensation.
Keywords
O
The kidney glomerular basement membrane (GBM) is unlike most basement membranes found elsewhere in the body. During early glomerular development, the basement membrane layer beneath ingrowing endothelial cells fuses with a layer beneath immature podo-cytes to yield a common, double-thickness basement membrane located between two adherent cell sheets. Subsequently, during glomerular capillary loop expansion, additional basement membrane material, derived mainly if not exclusively from podocytes, is inserted or spliced into the fused GBM (reviewed in Abrahamson 1991). The factors that mediate these fusion and splicing events are not known, and we have speculated previously that intermolecular binding interactions between proteins in the subendothelial and subepithelial basement membrane layers could promote the union of these two basal laminae (Clapp and Abrahamson 1994). Likewise, the mesangial matrices in axial regions of developing glomeruli are very loosely organized initially but become condensed during glomerular maturation. The formation of intermolecular bonds within the loose-knit mesangial matrix might therefore also result in its condensation. To test for the presence of binding sites for basement membrane molecules within developing glomeruli, we injected soluble mouse laminin-1
Materials and Methods
Laminin Purification and Digestion
Laminin was purified from the mouse Englebreth-Holm-Swarm tumor matrix by sequential salt extraction, DEAE chromatography, and gel filtration, using a modification of the method originally reported by Timpl et al. (1979) as previously described (Abrahamson and Caulfield 1982). At a concentration of 0.34 mg/ml in 10% acetic acid, laminin was digested for 24 hr at 4C with porcine stomach pepsin (3800 U/mg; Sigma Chemical, St Louis, MO) at an enzyme:sub-strate ration of 1:50 (w/w). The reaction was stopped by dialysis into PBS, pH 7.3. For elastase digestion, 0.3 mg/ml laminin in 0.05 M ammonium bicarbonate, pH 7.8, was treated for 4 hr at 37C with porcine pancreatic elastase (135 U/mg; ICN Pharmaceuticals, Costa Mesa, CA) at an enzyme:substrate ratio of 1:100 (w/w). The reaction was terminated by adding excess PMSF and the digest was dialyzed against PBS.
Affinity Purification of Laminin Fragments
Rat anti-mouse laminin MAb IgGs 5A2, 5C1, and 5D3 were purified from ascites fluid obtained from nude mice and were characterized by Western blotting and rotary shadow electron microscopy of affinity-purified laminin digests as previously described (Abrahamson et al. 1989; Steadman et al. 1993). To prepare affinity columns, 8.0 mg of each of the MAb IgGs were coupled separately to 3.0 ml cyanogen bromide-activated Sepharose 4B (Pharmacia Biotech; Piscat-away, NJ) as before (Steadman et al. 1993). Crude pepsin digests were applied to the MAb 5C1 column and, after washing with PBS, bound pepsin-resistant fragments were eluted with the addition of 3 M potassium thiocyanate, pH 6.0. The eluate was concentrated to 0.25 mg/ml by centrifugal filtration in a Centricon-30 tube (Amicon; Beverly, MA). Crude elastase digests were first applied to the MAb 5D3 column. Unbound fractions were then pooled and chromato-graphed over MAb 5A2-Sepharose. Laminin fragments bound to MAb 5D3 and 5A2 columns were then eluted, concentrated, and then assessed by PAGE and Western blot as before (Abrahamson et al. 1989; Steadman et al. 1993). As shown in Figure 1a, the MAb 5D3 affinity-purified elastase fragment blotted only with MAb 5D3 and not MAb 5A2. Likewise, the MAb 5A2 affinity-isolated fragment reacted with that antibody but not with MAb 5D3 (Figure 1a). The pepsin-resistant fragment affinity-isolated by MAb 5C1 was also not recognized by MAbs 5A2 and 5D3 (Figure 1b).
Detection of Laminin Binding Sites In Vivo
All experiments involving animals met institutional and NIH guidelines. Undigested mouse laminin was injected
Two to 14 days after the second
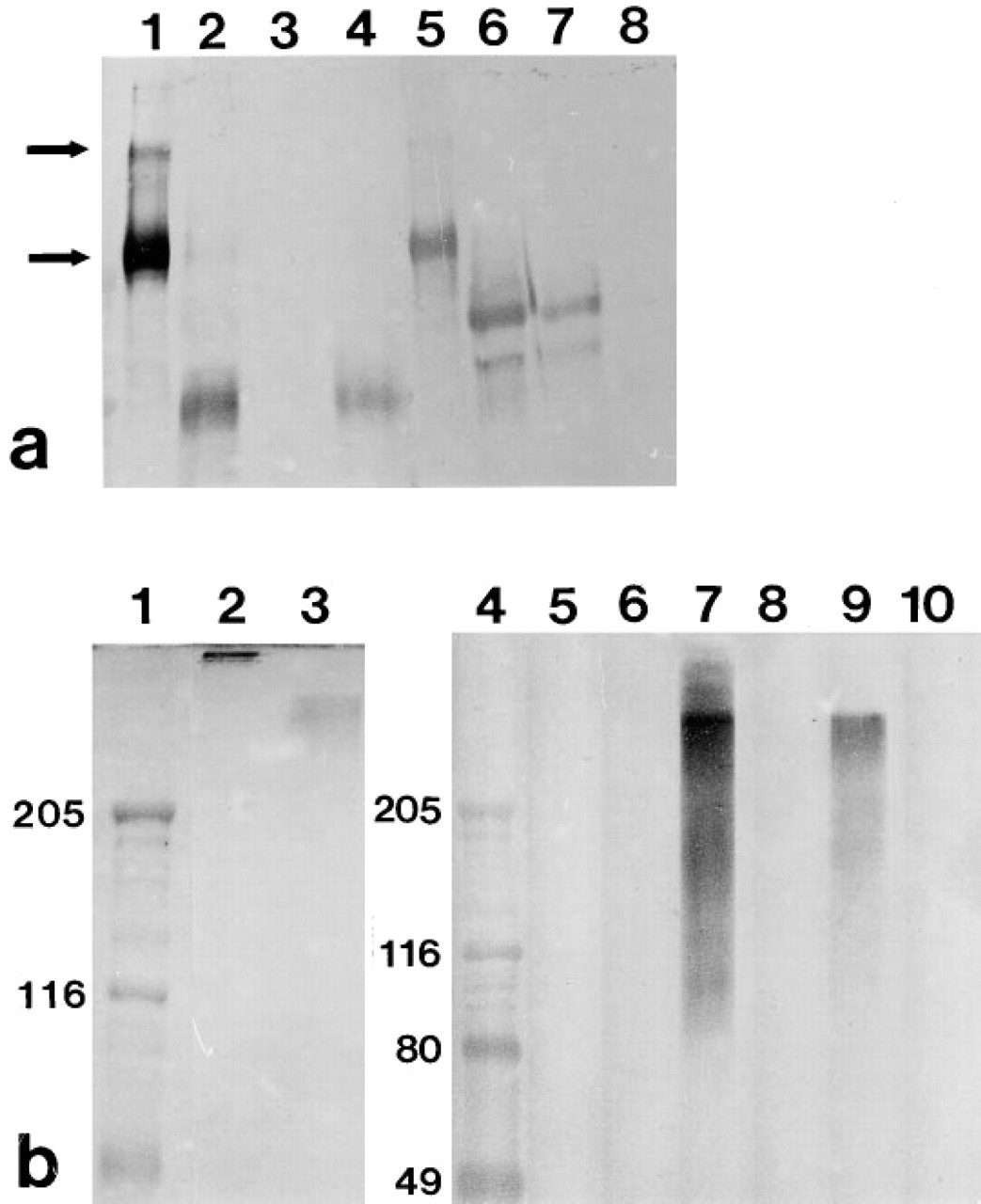
PAGE and immunoblot analysis of laminin epitopes with MAbs 5A2, 5C1, and 5D3. (
In addition to IV administration of mouse laminin into newborn rats, unfixed cryostat sections of infant rat kidneys were incubated in a humid chamber for 1 hr at room temperature with purified laminin ranging in concentration from 10 to 240 μg/ml. Sections were washed with PBS, treated with rat anti-mouse laminin MAbs, and processed for indirect immunofluorescence microscopy.
Immunoelectron Microscopy
Each anti-laminin MAb was also conjugated directly to activated horseradish peroxidase (HRP), exactly as described previously (Abrahamson et al. 1989), and was concentrated to 1.0 mg/ml. Newborn rats that had received IV injections of mouse laminin on Days 2 and 3 after birth were anesthetized on Day 4 and then re-injected through the left saphenous vein with 0.2 ml of one of the MAb-HRP conjugates. Adult rats received 1.5 ml MAb-HRP 2 days after the second laminin injection. Animals were returned to their litters and, 1 day after the MAb-HRP injection, they were anesthetized a final time. Kidneys were fixed in situ by subcapsular injection of 1.6% paraformaldehyde and 3% glutaraldehyde in 0.1 M sodium cacodylate, pH 7.4 (Karnovsky's fixative). Tissue slices 40 μm thick, were processed for peroxidase histochemistry and electron microscopy as before (Abrahamson and St. John 1992).
Results
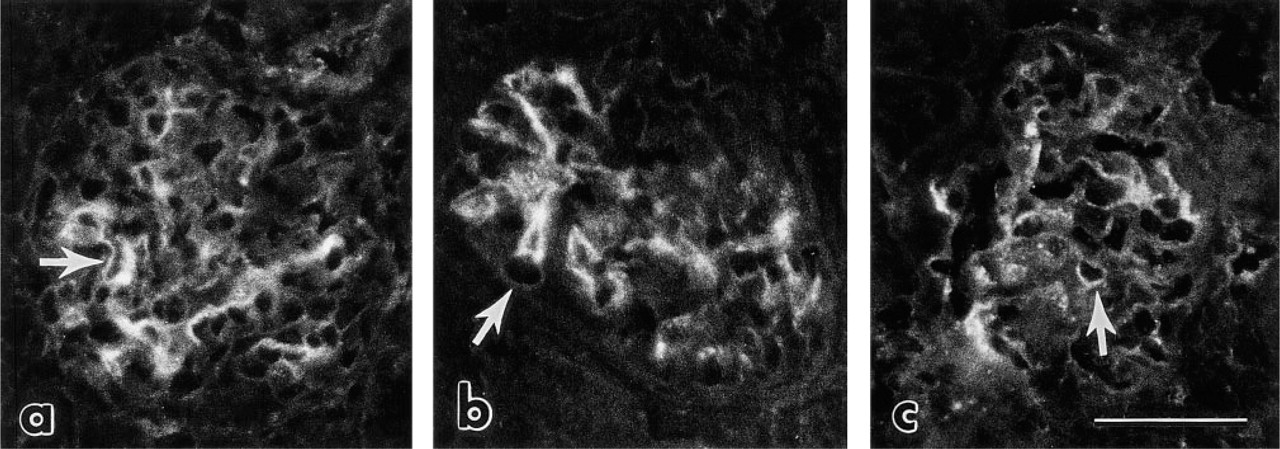
Preliminary studies showed that, after a single IV injection of intact mouse laminin into anesthetized newborn rats, there was no detectable binding of the injected protein to immature rat glomeruli in vivo. However, after two daily injections of exogenous laminin into 1-, 2-, or 3-day-old rats, immunofluorescence examination showed intense localization of injected laminin specifically to maturing glomeruli (Figure 2). As shown in Figures 2b, 2c, and 2d, anti-laminin MAbs 5A2, 5C1, and 5D3 all labeled glomeruli in similar patterns. The injected laminin bound abundantly to developing me-sangial matrices and lengths of peripheral capillary loop GBMs were positive as well (Figure 2). However, less mature glomeruli located beneath the capsule were only weakly labeled or unlabeled (Figure 2a). Because considerable nephrogenesis continues in rats and mice during the first week after birth, the immature glomeruli in superficial layers of the cortex that contained only small amounts of injected laminin were just beginning to form when the first laminin injection was given 4 days earlier. There was no binding of injected laminin to developing tubule basement membranes, perivascular matrices, or other structures in the infant rat kidney (Figure 2).
The ability to distinguish between the extrinsic mouse laminin that we injected and the intrinsic native laminin synthesized within the rat kidney was crucial for our experimental approach. As shown in Figure 3a, when cryostat sections from rat kidneys were incubated with rat anti-mouse laminin MAbs there was no binding whatsoever of the MAbs to basement membranes or other structures in the sections. This result therefore shows that the rat anti-mouse laminin MAbs did not recognize autologous rat laminin. Importantly, when glomeruli from newborn animals that had received two IV injections of sheep IgG as controls were then labeled with anti-sheep IgG–fluorescein, no binding of the injected IgG was observed (Figure 3b). This shows that the binding of injected laminin seen in Figure 2 did not merely reflect nonspecific assimilation of injected protein into developing glomeruli. Likewise, when adult rats that had received the same amount of injected laminin/g body weight as the newborns were assessed, no binding of injected laminin was seen in kidneys taken from mature animals (data reviewed but not shown). In addition, to assess whether the laminin binding sites identified in vivo (Figure 2) might be accessible ex vivo, we incubated unfixed cryosections of newborn rat kidneys at room temperature with mouse laminin ranging in concentration from 10 to 240 μg/ml. In all cases there was no binding of the extrinsic laminin to newborn kidney sections in vitro (reviewed but not shown).
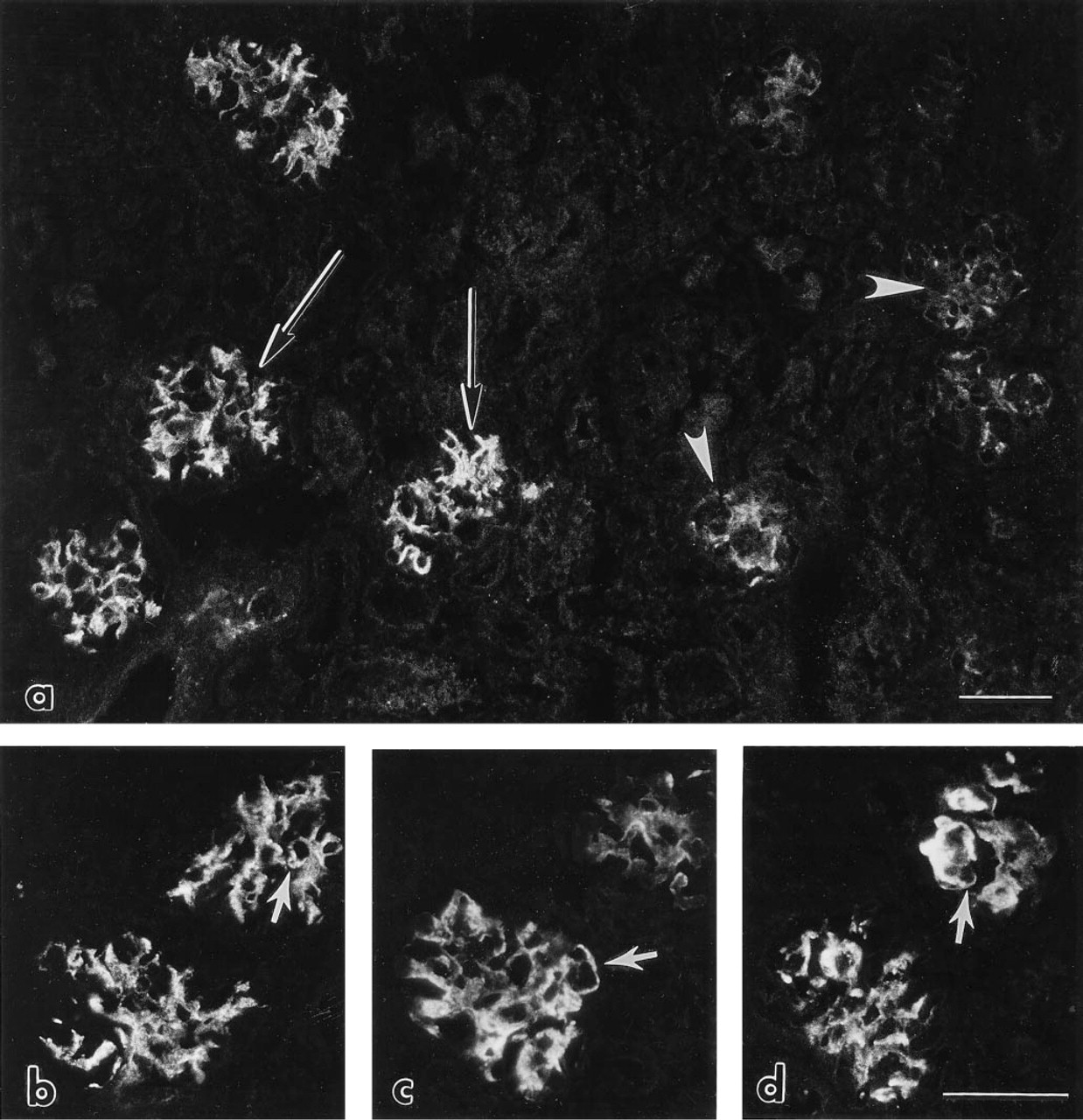
Immunofluorescence photomicrographs of newborn rat kidney after IV injection of mouse laminin on Days 2 and 3 after birth. Kidney was removed 2 days after the second injection and labeled with rat anti-mouse laminin MAbs. (
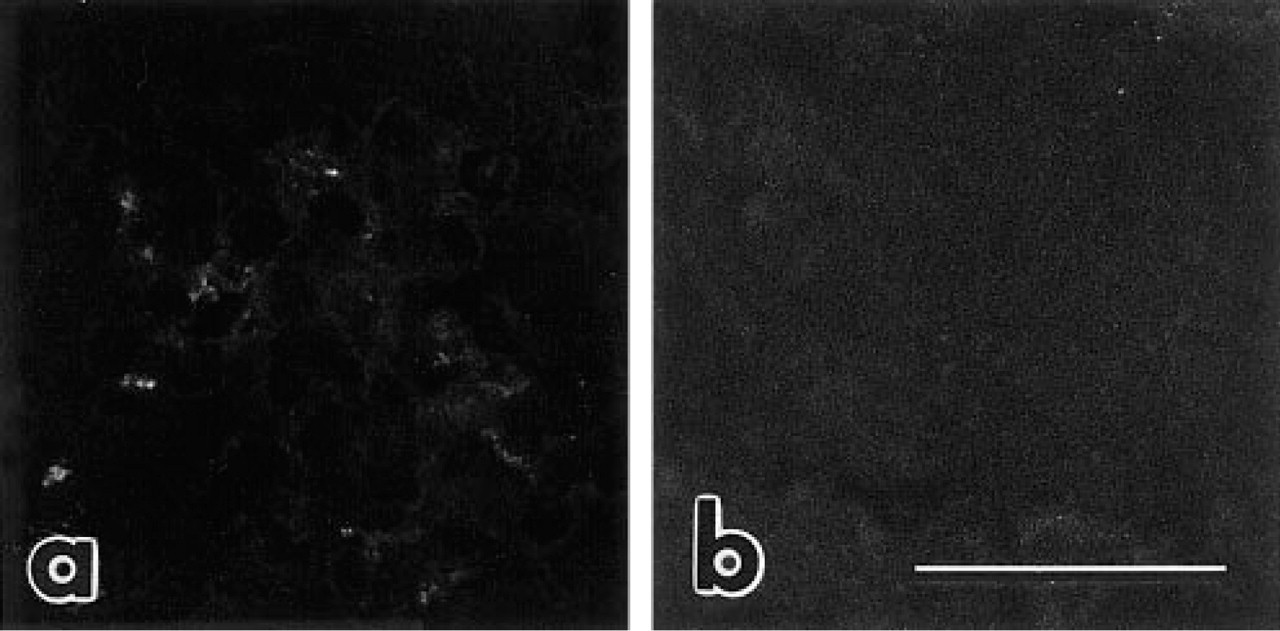
When control section from uninjected newborn rat kidney is labeled with rat anti-mouse laminin MAb 5C1, there is no binding of the MAb to intrinsic rat GBM laminin (
To examine the in vivo binding of laminin in greater detail, newborn and adult rats that had received the same amount of injected laminin/g body weight over a 2-day period were injected with MAb 5D3-HRP conjugates. As shown in Figure 4a, an intense peroxidase reaction product was seen in glomeruli of newborn rats, especially within mesangial matrices. A weak reaction product was also seen in peripheral GBMs of many capillary loops, and the same binding patterns were seen after in vivo labeling with 5A2- and 5C1-HRP. Those GBMs that were weakly labeled were found in loops that were still undergoing basement membrane splicing and podocyte foot process development, whereas capillary loops within the same glomeruli that appeared ultrastructurally mature did not contain injected laminin (Figure 4a). In adult rats there was no binding of injected laminin to mesangial matrices, GBMs, or elsewhere in kidneys (Figure 4b), and this was consistent with the immunofluorescence results.
To investigate the relative stability of the bound laminin, kidneys from rats that had received IV injections of laminin 2 days after birth were removed 3, 5, 7, 9, or 14 days after the second injection. Immunofluorescence labeling showed that over this time period considerable amounts of injected laminin remained bound within glomeruli (Figure 5). Mesangial areas continued to be intensely labeled and linear GBM labeling was also found in glomerular capillary loops (Figure 5). The labeling patterns seen 2 weeks after the last laminin injection were the same for MAbs 5A2, 5C1, and 5D3 (Figure 5).
Having shown that IV injected laminin bound specifically to developing GBMs and mesangial matrices and remained bound for at least 2 weeks after injection, we sought to identify the molecular domains on laminin that mediate this binding. Laminin was digested with pepsin or elastase and newborn rats received
The ages, numbers of animals injected with various preparations of intact laminin and proteolytic fragments of the molecule, and whether or not these injections resulted in glomerular binding are summarized in Table 1. The dominant variable for detection of injected laminin in glomeruli was the age of the animal at the time of injection. Animals that were 1, 2, or 3 days old when they received the first injection always contained injected laminin in their glomeruli. Beginning ~5 days after birth, however, injected laminin was variably seen in glomeruli of young rats and there was no incorporation of extrinsic laminin in adults (Table 1).
Discussion
Several conclusions can be drawn from our results. First, injections of mouse laminin-1 into newborn rats resulted in incorporation of this protein into mesangial matrices and GBMs of immature glomeruli. In contrast, injected laminin did not bind to other basement membranes of developing kidneys and, when injected into adult rats, there was no association with glomerular matrices or other tissues in the mature kidney. These findings therefore demonstrate that laminin binding sites are transiently present within basement membranes of developing glomeruli. Moreover, these sites were specific for intact laminin, and other proteins injected as controls, as well as proteolytic fragments of laminin, failed to bind to developing glomerular matrices. Binding of injected laminin to the developing GBM was stable and there were no perceptible changes in glomerular patterns of bound laminin for at least 2 weeks after injection.
Second, binding sites for laminin in immature glomeruli were identified only after
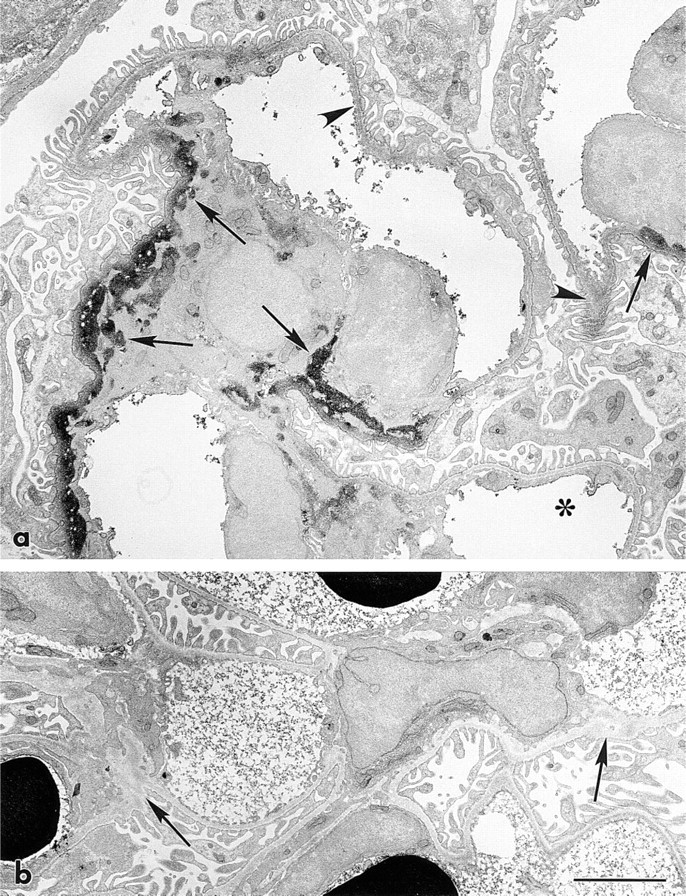
To investigate binding of injected laminin to developing GBM in greater detail, kidneys from newborn and adult rats that had received IV injections of laminin were labeled with MAb 5D3-HRP and processed for immunoelectron microscopy. In a maturing stage glomerulus from a newborn rat, intense peroxidase reaction product is found in mesangial matrices (arrows) and some much weaker reaction product is also seen in peripheral loop GBM (arrowheads) (
To evaluate the stability of bound laminin within the GBM, kidneys from young rats that had received IV injections on Days 2 and 3 after birth were examined on Day 16, 2 weeks after the second injection. (

Affinity-purified proteolytic fragments of laminin were injected IV into newborn rats and kidney sections were processed for immunofluorescence as before. Injection with elastase fragment affinity-purified with MAb 5A2. Section labeled with MAb 5A2. Unlike what is shown after injections of intact laminin (Figures 2, 4, and 5), the enzymatic fragment failed to bind developing glomeruli in vivo. Identical results were obtained after injection of MAb 5C1 and 5D3 affinity-purified fragments. Likewise, there was no labeling of sections taken from rats that received injections of crude elastase or pepsin digests. Bar = 60 μm.
Third, injected laminin bound abundantly to glomeruli of 1-3-day-old rats, in which extensive GBM and mesangial matrix formation is still taking place. In contrast, only variable glomerular binding was seen when laminin was injected into 5–10 day-old rats, in which glomerular development is concluding. No binding was detected when laminin was injected into adults. The absence of binding of injected laminin in older animals might have been due to the progressive acquisition of permselective sieving properties of the mature glomerular capillary wall, which would therefore minimize binding of circulating laminin to potential GBM binding sites. However, mesangial areas of adult glomeruli are considered to be much more permeable to circulating macromolecules than the GBM, but injected laminin localized only to mesangial matrices and GBMs in developing rats, and this was never seen in adults. Laminin binding in newborns was also not immediate, and only those developing glomeruli that had been exposed to circulating laminin over a period of several days were positive. Considering these findings, we believe that the in vivo glomerular binding of injected laminin in infant rats reflects accumulation of this protein in GBMs and mesangial matrices undergoing extracellular assembly.
Northern analysis, ribonuclease protection assays, and/or in situ hybridization studies have shown that mRNAs encoding the α1- and α2-chains of collagen Type IV, the core protein of perlecan, and the α1-, β1-, and γ1-chains of laminin are all present at relatively high levels during kidney development (Ekblom et al. 1990; Vanden Heuvel and Abrahamson 1993; Miner and Sanes 1994; Vanden Heuvel et al. 1996). With the exception of the perlecan transcript (Vanden Heuvel and Abrahamson 1993), these mRNAs normally decline to low or imperceptible levels in the mature kidney. Immunolocalization studies have shown, however, that collagen Type IV and laminin polypeptide chains contained within the early GBMs of the most immature glomeruli (located in subcapsular areas) differ from those of more mature glomeruli located deeper in the cortex of the same kidney. Specifically, the most immature glomeruli label for the α1- and α-2-chains of collagen Type IV along with a “fetal” isoform of laminin containing α1, β1 and γ1 laminin chains (Miner and Sanes 1994; Durbeej et al. 1996). In contrast, the GBMs of maturing stage glomeruli lack these polypeptides and instead include the α3-, α4-, and α5-chains of collagen Type IV and the α5, β2, and γ1 laminin chains (Miner and Sanes 1994; Durbeej et al. 1996).
Binding of intravenously injected mouse laminin, crude elastase and pepsin digests, affinity-isolated laminin fragments, and sheep IgG to rat glomeruli

Numbers in parentheses show numbers of rats analyzed at each time point.
+, all maturing stage glomeruli were intensely positive; ±, some glomeruli were variably positive, many were weak/negative; -, all glomeruli were negative.
What regulates the appearance and disappearance of different GBM protein isoforms during development is not known, but these changes are important for the morphological and physiological maturation of the glomerulus. For example, homozygote mutant mice lacking the α3-chain of Type IV collagen die of renal failure 3–4 months after birth (Miner and Sanes 1996). GBMs in mutants at the time of death are mul-tilaminated and resemble those of humans with Alport syndrome. In addition to a loss of the collagen α3(IV)-chain in these mice, there is also an absence of collagen α4- and α5(IV)-chains and abnormally increased amounts of collagen α1- and α2(IV)-chains, collagen VI, and perlecan (Miner and Sanes 1996). Glomerular defects are also observed in mice genetically deficient for the β2-chain of laminin, and these animals die before 1 month of age (Noakes et al. 1995). Unlike mice with α3(IV) collagen deficiencies, however, GBMs in these animals are ultrastructurally normal and contain the appropriate complement of GBM collagens and perlecan. In contrast, the laminin β1-chain, which ordinarily is replaced by the β2-chain, persists in mature glomeruli of these β2 chain mutants, and unusually broad podocyte foot processes are also present (Noakes et al. 1995). As is the case in α3(IV) collagen deficiency, mechanisms accounting for dysregulation of other basement membrane gene products in laminin β2 mutants are not known but may reflect sensory feedback at the level either of gene expression or of post-translational GBM protein assembly (Noakes et al. 1995). Hence, in the absence of correct collagen or laminin chains, a compensatory synthetic response of different collagens and/or laminins may occur. Molecular mismatches between incorrect isoforms may then take place that result in structural and functional GBM defects.
When we examined young rats that had received the last injection of mouse laminin 2 weeks earlier, we found that injected laminin remained associated with GBMs of mature glomeruli. Furthermore, labeling with MAb 5A2, which recognizes an epitope on the short arm (Abrahamson et al. 1989) of the laminin β1-, but not the β2-chain (Martin et al. 1995), showed that the mouse laminin incorporated into rat glomeruli still contained the 5A2 β1-chain epitope. As described previously (Abrahamson and St. John 1992), however, GBMs of immature glomeruli in mice express the 5A2-reac-tive laminin epitope, but those of maturing glomeruli progressively become negative. Earlier, we speculated that this progressive loss of epitopes from developing GBMs may be due to (a) enzymatic processing during GBM fusion/splicing or (b) replacement of one lami-nin isoform with another (Abrahamson and St. John 1992). That the 5A2 β1-chain epitope on the injected laminin persisted in GBMs of maturing rat glomeruli strongly suggests that the epitope loss that normally occurs is not due to enzymatic removal. In addition, unlike mice genetically deficient for the laminin β2-chain, in which compensatory synthesis of laminin β1-chain results in glomerular pathology (Noakes et al. 1995), there were no obvious structural defects imposed in rats by the atypical presence of laminin containing the β1-chain. We believe, however, that relative to the native laminin synthesized by the rat, only minor amounts of injected mouse laminin actually became stably incorporated into rat GBMs.
The domains on laminin known to bind basement membrane proteins with high affinity in vitro include the highly homologous N-termini (Domain VI) of the α-, β-, and γ-chains, which interact with other laminin molecules to form laminin polymers (Yurchenco and Cheng 1993; Yurchenco and O'Rear 1994) and to Domain III on the γ chain that binds to the G3 globular domain at the C-terminus of entactin (Mayer et al. 1993; Beck and Gruber 1995). The most prominent heparin binding domain is found at the C-terminal G domain of the laminin α-chain (Timpl and Brown 1994; Yurchenco and O'Rear 1994; Beck and Gruber 1995). To identify which domains of injected laminin mediated its binding to developing GBM in vivo, we digested laminin with pepsin or elastase and affinity-isolated three subfragments with domain-specific MAbs. As shown earlier (Abrahamson et al. 1989; Steadman et al. 1993), the fragment recognized by MAb 5A2 corresponds to the N-terminal domain of the short arm of the laminin β1-chain, the 5C1 affinity-purified P1 fragment represents a central, three-armed region of the molecule, and the 5D3 affinity-purified fragment corresponds to the C-terminal end of the long arm. Within each of these fragments are regions containing binding domains for laminin, entactin, and per-lecan, respectively (Yurchenco and O'Rear 1994; Beck and Gruber 1995). Although abundant glomerular binding was seen after
In conclusion, what we have shown in this study is that, when injected into newborn rats, intact mouse laminin bound specifically to GBMs and mesangial matrices of developing glomeruli. This injected laminin remained bound for at least 2 weeks after injection, and the β1-chain of this laminin, as recognized by the binding of MAb 5A2, also persisted. We suspect that the binding sites we have identified in developing glomeruli are probably crucial for the formation of intermolecular crosslinks during extracellular basement membrane assembly and may be important specifically for GBM fusion and mesangial matrix condensation in vivo. Because three separate proteolytic fragments of laminin failed to bind, however, our results suggest further that multivalent linkages between laminin and its ligands are required for stable incorporation into matrices. Future studies aimed at determining precisely which laminin domains mediate its binding, and to which molecules these laminin domains bind, should provide a much better understanding of basement membrane assembly and organization in vivo.
Footnotes
Acknowledgements
Supported by NIH grants DK34972 and DK52483. The hybridoma core facility at UAB is supported by AR20614, and the electron microscopy core is a shared facility of the Comprehensive Cancer Center (CA13148).
We thank Barry Robert for critically reviewing an earlier version of the manuscript.