
Abstract
Fracture healing in long bones is a sequential multistep cascade of hemostasis, transient inflammation, chemotaxis of progenitor cells, mitosis, differentiation of cartilage, and replacement with bone. This multistep cascade is orchestrated by cytokines and morphogens. Members of the interleukin (IL)-17 family, including IL-17B, have been identified in cartilage, but their expression during fracture healing is unknown. In this study, we determined the immunolocalization of cytokines IL-17A and IL-17B, along with the IL-17 receptor (IL-17R) and IL-17 receptor-like protein (IL-17RL), during the sequence of fracture repair in a standard model. The results were extended to developmental changes in the epiphyseal growth plate of long bones. Members of the IL-17 family were localized in chondrocytes in the fracture callus. Moreover, we found significant parallels to the localization of these cytokines and their receptors in chondrocytes during an endochondral differentiation program in the epiphyseal growth plate.
Keywords
T
Interleukin (IL)-17B is a member of the IL-17 family of inflammatory cytokines (Li et al. 2000; Shi et al. 2000; Moore et al. 2002), and we have recently described the expression of IL-17B in articular cartilage (Moseley et al. 2003). IL-17 was originally identified as a pro-inflammatory cytokine expressed only in T cells (Rouvier et al. 1993; Yao et al. 1995a, b; Albanesi et al. 2000), although its receptor is ubiquitously expressed in most tissues. IL-17B shares significant homology to IL-17 but is expressed in tissues and cells other than T cells, including cartilage. A recently discovered IL-17 receptor-like molecule (IL-17RL, also known as IL-17RC) is also expressed by chondrocytes (Haudenschild et al. 2002). However, the functions of IL-17 cytokines, their receptors in cartilage, and their roles as pro-inflammatory factors in bone fracture healing remain largely unknown.
In this study, we determined the immunolocalization of IL-17A, IL-17B, IL-17 receptor (IL-17R), and IL-17 receptor-like protein (IL-17RL) during the differentiation of chondrocytes, which is an integral part of the fracture healing cascade. The regeneration of tissue in fracture healing recapitulates many of the events in cartilage and bone morphogenesis during development (Reddi 1998). Therefore, to extend the findings of the fracture healing studies to chondrocyte differentiation, we examined the localization of the IL-17 cytokines and their receptors in the epiphyseal growth plate of rats.
Materials and Methods
Fracture Model
Eight Long Evans rats were used to obtain standard closed fractures. The mean age of the rats was ∼12 weeks. All surgical procedures were performed under anesthesia using halothane and ketamine-xylazine. A lateral parapatellar knee incision was made to expose the distal femoral condyle, and a 1.25-mm-diameter K-wire was inserted from the trochlear groove into the femoral canal. A closed transverse femoral shaft fracture was created using a three point bending apparatus, following the method of Bonnarens and Einhorn (1984). Postoperative pain was managed by the administration of a subcutaneous injection of buprenorphine hydrochloride. The rats were fed a standard maintenance diet and provided water ad libitum. Unprotected weight bearing was allowed immediately postoperatively. This research protocol was approved by the Institutional Animal Care and Use Committee. At 1 and 2 weeks after fracture, four rats were killed with an excess of carbon dioxide gas and processed for histological study. Fractured femurs were harvested and fixed in 4% paraformaldehyde for 24 hr at 4C, defatted in ethanol, decalcified with 10% formic acid in citrate for 4 days at 4C, and embedded in paraffin. Paraffin sections (4 μm thick) were cut and stained with toluidine blue for histological observation. Sections were also subjected to immunohistochemistry for cytokines IL-17A and IL-17B and receptors IL-17R and IL-17RL.
Rat Growth Plate
Newborn Long Evans male rats were killed 21 days after birth. Proximal ends of tibia were harvested and fixed in 4% paraformaldehyde in phosphate buffer, defatted in ethanol, decalcified with 10% formic acid, and embedded in paraffin. Paraffin sections (5 μm thick) were cut and stained with toluidine blue for histological observation. Sections were also subjected to immunohistochemistry for IL-17A, IL-17B, IL-17R, and IL-17RL.
Cytokines and Antibodies
Rabbit polyclonal antibodies of IL-17B and IL-17RL were made against synthetic peptides corresponding to the N terminus of IL-17B (NH3-CQPRSPKSKRGKQGRP-COOH) and extracellular and cytoplasmic portions of IL-17RL (NH3-QP-RYEKELNHTQQLPDC-COOH and NH3-DSYFHPPGTPA-PGR-COOH, respectively). To affinity-purify the antipeptide IgG from the serum, the peptides were immobilized on Sulfo-Link beads (Pierce; Rockford, IL) and washed and eluted as recommended by the manufacturer. Purified antibodies were used at 2 μg/ml for Western blotting and 20–30 μg/ml for immunohistochemistry, as previously published (Haudenschild et al. 2002; Moseley et al. 2003). Recombinant human IL-17R was purchased from R&D Systems (Minneapolis, MN). Mouse monoclonal antibody raised against human IL-17A and goat polyclonal antibody against human IL-17R extracellular domain were purchased from R&D Systems and used according to the manufacturer's recommendations.
Immunohistochemistry
Immunohistochemistry of IL-17, IL-17B, IL-17R, and IL-17RL were performed by the avidin-biotin alkaline phosphatase complex method, using a Vectastain ABC-AP standard kit (Vector Laboratories; Burlingame, CA). After deparaffinization and hydration, nonspecific reactions were blocked using 5% normal serum for 30 min at room temperature. Endogenous alkaline phosphatase was blocked with Levamisole (Vector Laboratories). Specimens were incubated with the appropriate concentrations of primary antibodies at 4C overnight in a humidified chamber. Incubation with biotinylated secondary antibody was performed for 60 min at room temperature, and an avidin-biotin alkaline phosphatase complex reaction was performed for 30 min at room temperature. Color was developed with a Vector Red alkaline phosphatase substrate kit (Vector Laboratories) without any counterstaining. Normal mouse IgG, normalpreimmune rabbit IgG, or Tris-buffered saline (TBS) was used as a negative control instead of the primary antibodies, and conditions used such that there was no signal in the negative control samples. The slides were viewed with a Zeiss Axioplan 2 microscope, and images were captured on a Windows NT work-station using Zeiss Axiovision software (Zeiss; New York, NY).
Results
Characterization of Polyclonal Antibodies
The specificity of the antibody against the IL-17 cytokines was confirmed by Western blotting. The antibody targeting the N terminus of IL-17B detects both the glycosylated and deglycosylated purified recombinant human IL-17B generated in yeast. There is a single band detected in bovine calf cartilage homogenate corresponding in size to the deglycosylated recombinant protein. There is no cross-reactivity with purified recombinant human IL-17A. These results are shown in Figure 1. The antibodies against IL-17RL protein have been previously described (Haudenschild et al. 2002; Moseley et al. 2003) and do not cross-react with IL-17R.
Localization of IL-17 Cytokines and Receptors
Endochondral Ossification During Fracture Healing. A histological section of a fractured femur at 1 week after fracture showed abundant newly formed callus at the fracture site, including intramembranous ossification and endochondral ossification (Figure 2A). The endochondral ossification area consisted of primitive chondrocytes and prehypertrophic chondrocytes, sandwiched by newly formed trabecular bone and mesenchymal stromal cells in fibrous tissue (Figure 2B). Hypertrophic chondrocytes were not found at this stage, but at 2 weeks after fracture, the region of endochondral ossification was mostly filled with hypertrophic chondrocytes (Figure 3B).
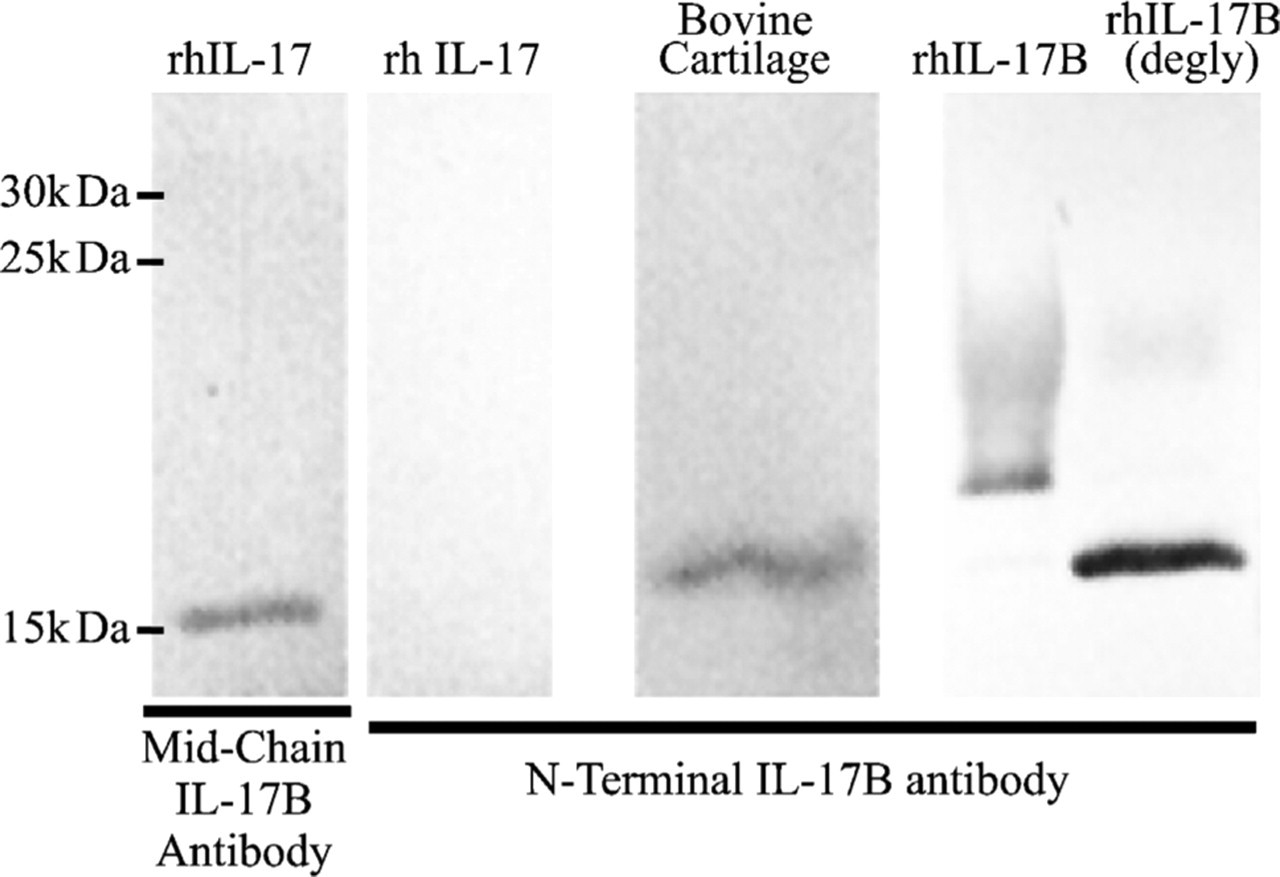
Immunoblot showing specificity of interleukin (IL)-17B antibodies. Lane 1: purified IL-17A cross-reacts with the pan-specific IL-17 mid-chain antibody. Lanes 2–5: the antibody designed against the N terminus of IL-17B recognized only IL-17B as its native form in bovine cartilage homogenate (Lane 3) and as a recombinant protein expressed in Pichia pastoris with or without deglycosylation (Lanes 4 and 5). No cross-reactivity of this antibody with pure recombinant IL-17A is detected (Lane 2).
IL-17A was detected both 1 and 2 weeks after fracture and localized only in prehypertrophic chondrocytes in the middle of the endochondral ossification (Figures 2C and 3C).
On the other hand, IL-17B was detected strongly in primitive and prehypertrophic chondrocytes 1 and 2 weeks after fracture (Figures 2D and 3D). Osteoblasts in the newly formed trabecular bone and mesenchymal stromal cells at the fracture site were positive for IL-17B at 2 weeks (Figure 3D). There was little IL-17B in hypertrophic chondrocytes at 2 weeks.
The receptors of IL-17s were widely localized at the fracture site after 1 week. IL-17R was localized in primitive and prehypertrophic chondrocytes, osteoblasts in the trabecular bone, and mesenchymal stromal cells in the fibrous tissue (Figures 2E and 3E). The intensity of staining of IL-17R in prehypertrophic chondrocytes close to bone was slightly weaker than in the other cells. The staining pattern of IL-17RL was similar to that of IL-17R, with the exception that the localization of IL-17RL in prehypertrophic chondrocytes was barely detected (Figures 2F and 3F).
The cellular distribution of the IL-17 cytokines and receptors in unfractured bone was not different from that observed in the cortical portions of bone adjacent to the fracture callous (data not shown).
Growth Plate in Rat Proximal Tibia. The cellular pathways of bone fracture healing mimic aspects of the embryonic and postnatal development of bone and cartilage. We therefore wanted to test whether IL-17 cytokines and receptors are also expressed in developing bones. To this end, we examined rat proximal tibias 21 days after birth. The growth plate consists of the resting, proliferating, prehypertrophic, and hypertrophic chondrocytes (Figure 4A). The growth plate is morphologically classified into four zones (in proximal toward distal order): the progenitor, proliferating, prehypertrophic, and hypertrophic zones (Figure 4B).
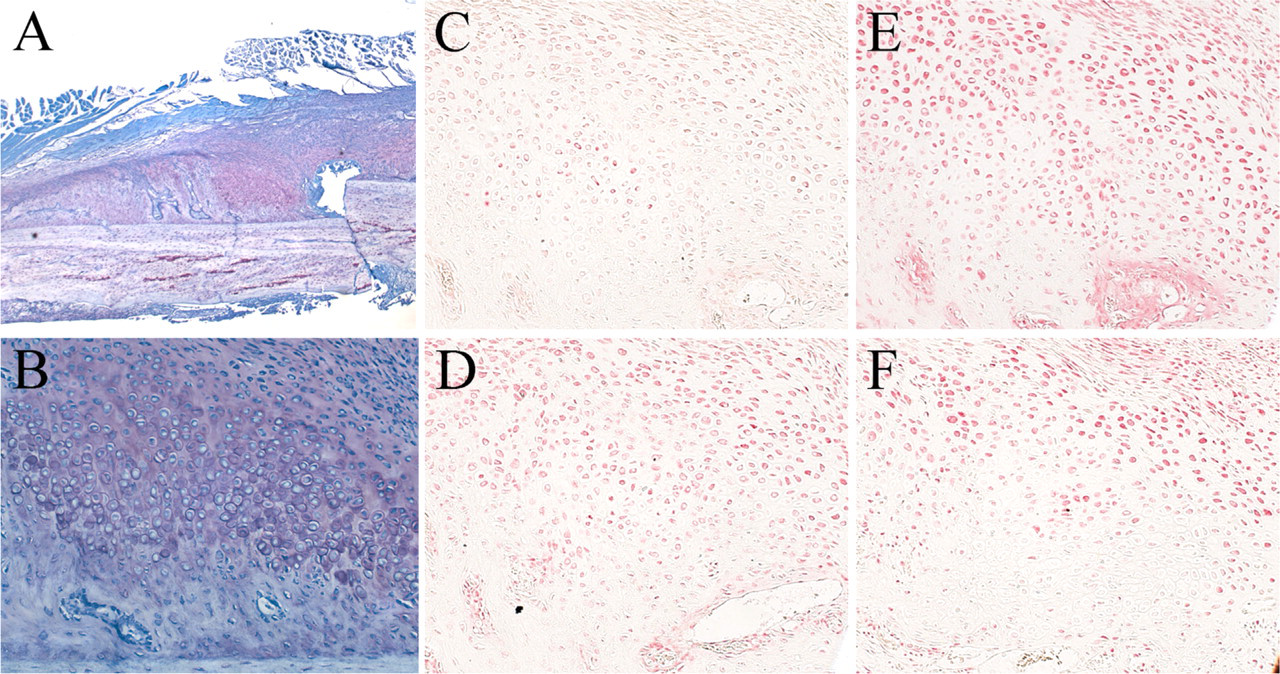
One-week fracture healing time point. (
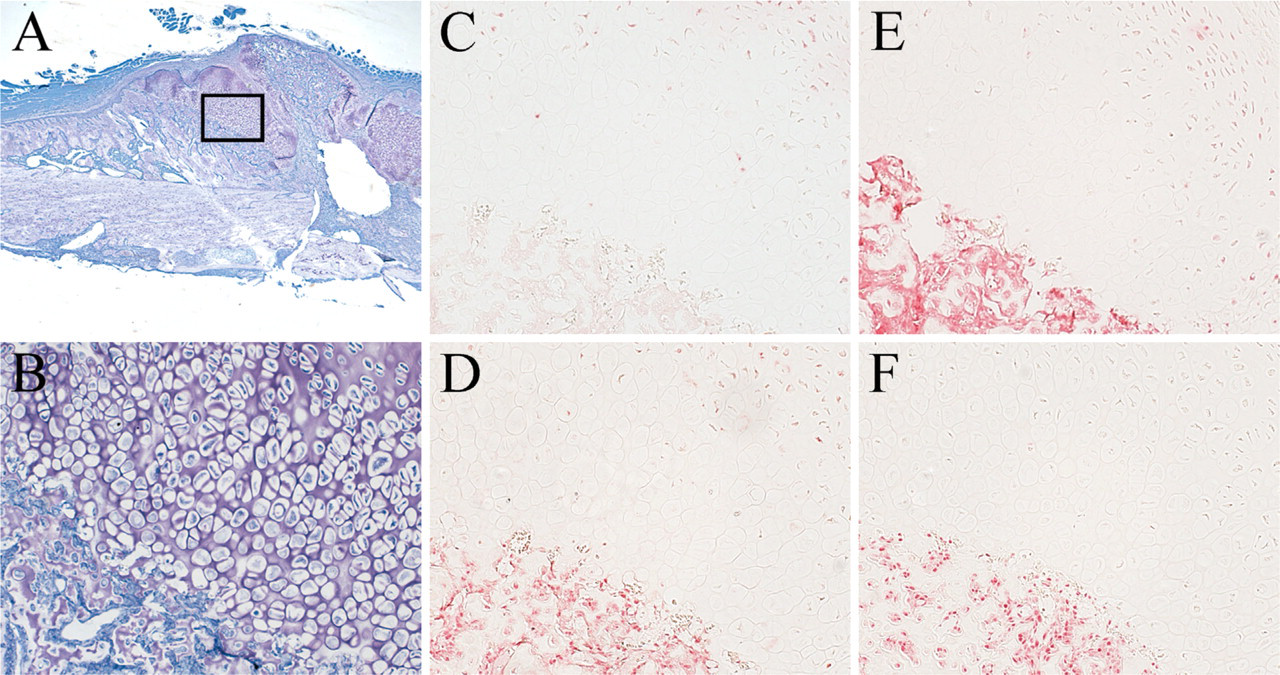
Two-week fracture healing time point. (
IL-17A was weakly localized in the prehypertrophic zone, slightly shifting to the proliferating zone across the boundary of both zones. There was little overlap between cells expressing the IL-17 cytokine and those expressing IL-17R (Figure 4C).
IL-17B was localized in some but not all prehypertrophic chondrocytes and progenitors but not hypertrophic and proliferating chondrocytes (Figure 4D).
The localization of IL-17 receptors was very similar to that of IL-17B. IL-17R and IL-17RL were strongly detected in both prehypertrophic chondrocytes and progenitor cells of the growth plate (Figures 4E and 4F). There was weak or no expression of the IL-17R or IL-17RL in proliferating and hypertrophic cells.
Discussion
IL-17 cytokines were originally described as proinflammatory molecules secreted by T cells. Chronic inflammation of the synovium and progressive destruction of articular cartilage and bone are well documented in rheumatoid arthritis (RA). In RA, T cells infiltrate the synovium and secrete detectable amounts of IL-17A into the synovial fluid (Chabaud et al. 1999), which induces a multitude of factors contributing to disease progression including degradation of the articular cartilage and erosion of the underlying bone. The annotation of the genomes led to the identification of a family of IL-17 cytokines (IL-17A through IL-17F) produced by tissues other than the immune system and a family of IL-17 receptors also produced in many tissues. The dramatic effect of IL-17A in joint disease led us to perform a closer examination of the expression of these cytokines in development and wound healing of bone.
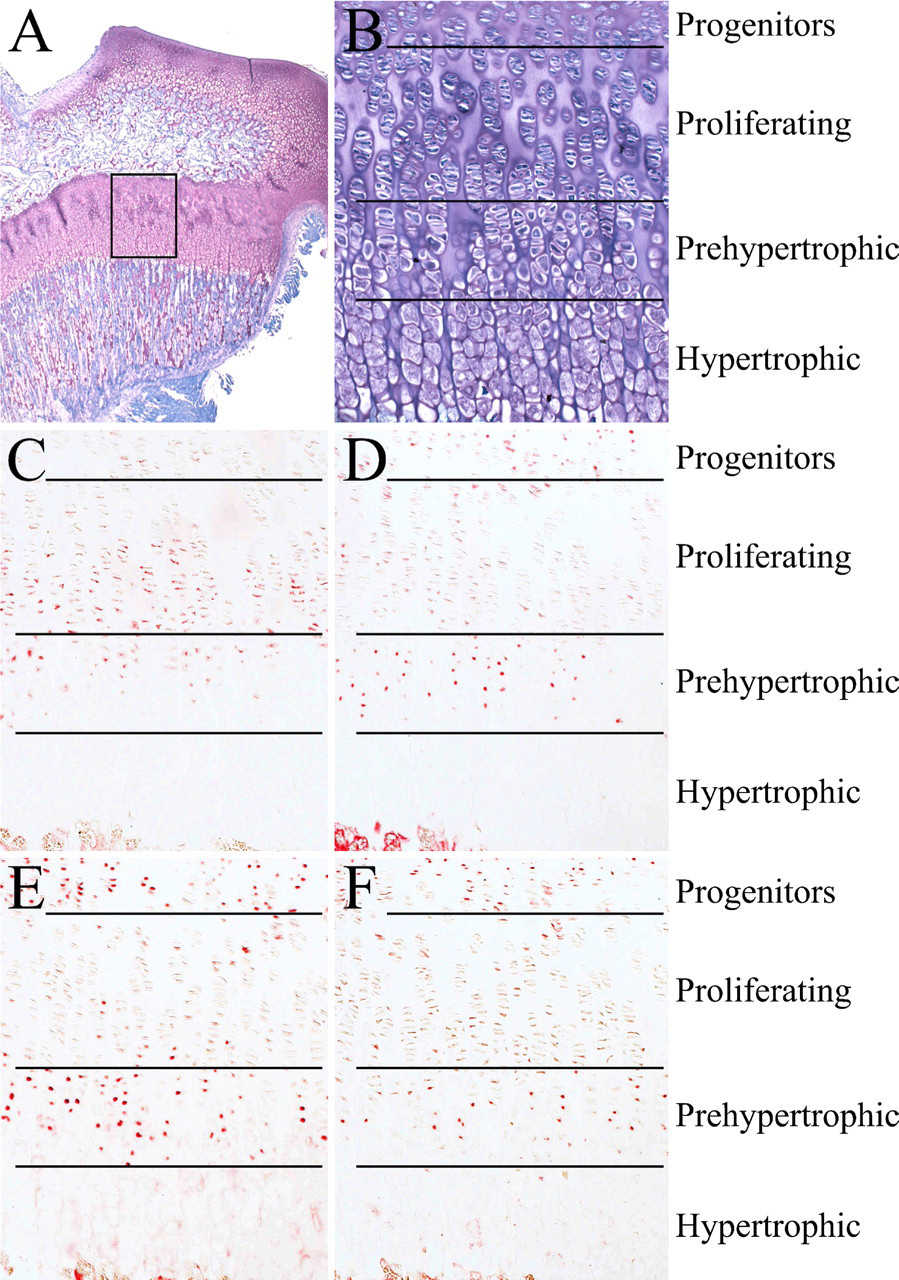
Rat epiphyseal growth plate. (
The consensus in the current literature on IL-17A is that it is primarily expressed by T cells, with little or no expression in other cell types (Lubberts 2003; Moseley et al. 2003; Kawaguchi et al. 2004; Kolls and Linden 2004). We detected IL-17A by immunohistochemistry in the prehypertrophic cells of the healing fracture at 1 week (Figure 2C), and to a lesser extent at 2 weeks after fracture (Figure 3C). There are two possible sources of IL-17A during fracture healing. First, the IL-17A may have been produced by T cells during the inflammatory stage of fracture healing and was detectable by immunohistochemistry because it remained associated with the matrix. T cells, if present, are only prominent during the first 24–48 hr after fracture, and generally do not persist to 1 or 2 weeks. Alternatively, the IL-17A may have been produced by the prehypertrophic chondrocytes to function in an autocrine manner. In this study, we were unable to distinguish between these possibilities because of the inherent limitations of immunohistochemistry. Intriguingly, we also detected IL-17A in the prehypertrophic chondrocytes of 21-day-old rat epiphyseal growth plates where there was no inflammation. This lends credibility to our notion of a non–T cell–derived source of IL-17A, such as an autocrine secretion of IL-17A by chondrocytes during the fracture healing process. This parallels the observed localization of IL-17A in the epiphyseal growth plate. The expression of IL-17A during embryonic and postnatal development needs to be explored more thoroughly at both the mRNA and protein levels.
In the growth plate, IL-17A was detected in proliferating cells, where the receptor for IL-17A receptor was not detectable (Figure 4). A possible interpretation is that the cytokine is produced by proliferating cells and stored in the extracellular matrix to act on the cells as they reach the prehypertrophic stage when they express the receptor. This would involve an “autocrine with time-delay” mechanism. However, this is speculation, because the IL-17 cytokines were not readily detected in the extracellular matrix of the growth plate (Figure 4). Alternatively, IL-17A may act in a paracrine manner, produced by the proliferating cells and diffusing out to act on the surrounding progenitor cells and prehypertrophic cells that express the receptor.
IL-17A can be a stimulator of osteoclastogenesis through the up-regulation of osteoclast differentiation factor (ODF or osteoprotegerin) (Kotake et al. 1999; Moseley et al. 2003). Thus, it is possible that IL-17A plays a role not only in bone formation but also in bone remodeling. The IL-17A knockout mouse shows no apparent skeletal defects, but perhaps this can be explained by a compensatory effect of IL-17F, which has very similar effects as IL-17A and can signal through the same receptor (Hymowitz et al. 2001; Zhou et al. 2007). However, the functions of IL-17A and IL-17F are not completely redundant, because the IL-17A knockout mouse displays increased resistance to collagen-induced arthritis (Nakae et al. 2003).
IL-17B was localized in prehypertrophic chondrocytes of both the growth plate and the fracture callus. This finding is supported by the identification of IL-17B protein and mRNA in adult bovine articular cartilage (Moseley et al. 2003), which provides evidence that the cytokine is synthesized by chondrocytes in normal cartilage. Cartilage development is in large part dependent on the actions of anabolic growth factors including BMPs and cartilage-derived morphogenetic proteins (CDMPs). Catabolic cytokines, including IL-17, IL-1, and tumor necrosis factor-α, lead to the breakdown of cartilage matrix, and their overexpression correlates with the progression of arthritis. Both cartilage development and homeostasis of normal adult tissue likely require a precise balance of the anabolic and catabolic activities.
IL-17R and IL-17RL localization is very similar in both fracture healing and in the tibial growth plate, which is consistent with the generation of a heterodimeric receptor complex consisting of IL-17R and IL-17RL (Toy et al. 2006), and it has recently been shown that siRNA inhibition of either receptor protein affects IL-17A signal transduction (Zhou et al. 2007). Both receptor proteins are strongly localized in progenitors cells, not detected in the proliferating cells, strong in prehypertrophic chondrocytes, and undetectable again in hypertrophic chondrocytes. There may be a common regulation of these proteins.
In conclusion, we described the localization of the cytokines IL-17A and IL-17B during bone fracture healing and in the epiphyseal growth plate of rats. We also examined the localization of IL-17R and IL-17RL and showed that they are localized in a very similar pattern. In general, there are very good parallels between the localization of these IL-17 cytokine and receptor molecules during fracture healing and in the growth plate. In the future, cartilage-specific gene knockouts and conditional mutants of the IL-17 signaling system in mice will shed more light on the role of IL-17A and IL-17B in embryonic chondrogenesis and in tissue regeneration. After all, tissue regeneration is, in part, a recapitulation of embryonic development and morphogenesis (Reddi 1998).
Footnotes
Acknowledgements
This work was supported by the Lawrence J. Ellison endowment and the Lawrence J. Ellison Chair.