
Abstract
cp27 is a novel gene involved in early vertebrate development that features a distinct protein localization pattern in developing tooth organs. During initial tooth development, CP27 was detected at the epithelial-mesenchymal interface of dental lamina stage tooth organs. At later stages of tooth development, CP27 was localized in the stellate reticulum, the oral mucosa mesenchyme, and alveolar bone. The significant changes in the highly restricted distribution pattern suggest that CP27 might be involved at several different levels during tooth development.
Keywords
T
To reveal the localization pattern of CP27 during tooth development, we prepared tissues from mice at stages E11, E13, E15, E19, and 3 days postnatal, according to Baylor College of Dentistry animal care guidelines. For paraffin immunohistochemistry, tissues were fixed in formalin, dehydrated, and embedded in paraffin as previously described (Diekwisch et al. 1997). Immunoreactions revealing CP27 localization patterns were performed using a polyclonal antibody against two CP27-derived oligopeptides that were designed using antigenicity criteria. Immunoreactions were performed as previously described (Diekwisch et al. 1997). The concentration of the primary antibody was 1:100. Anti-rabbit IgG at a concentration of 1:100 was used as secondary antibody. The primary antibody was incubated for 1 hr and the secondary antibody was incubated for 25 min. Signals were detected using AEC as a substrate. In between these steps, slides were washed in PBS. The following controls were used to determine the specificity of the anti-CP27 antibodies: (a) for tissue controls, the specificity of the antibody was evaluated in tissues with known immunoreactivity; (b) antibody controls by using a dilution series; (c) controls with preadsorbed anti-CP27 antibody to exclude unspecific binding; (d) controls with pre-immune serum to control for binding to serum components; (e) omission of primary antibody as a systemic control; (f) control peptide antibodies against a 10-amino-acid sequence of the amelogenin gene to test the specificity of the peptide sequence; and (g) a positive control peptide antibody.
At the beginning of tooth development, CP27 IHC delineated a distinct and highly confined localization pattern. During the dental lamina stage (E11) CP27 was localized in the basement membrane of the dental lamina and in mesenchymal cells adjacent to the dental lamina (Figure 1A and 1B). In the mesenchymal cells, CP27-related epitopes were localized either extracellularly or in the periphery of the cytoplasm, but not in the epithelium. During the bud stage (E13), CP27 was localized in the basal lamina of the oral epithelium. At this stage (E13), CP27 was also localized in a 100-nm-diameter half dome-shaped area of mesenchymal cells surrounding the developing tooth buds (Figure 1C and 1D). In contrast, the condensed dental mesenchyme and the dental follicle did not demonstrate any CP27 immunoreactivity. The immunopositive areas in the mesenchyme were restricted to extracellular matrices and the mesenchymal stroma. In the bell stage (E15) and all of the following stages until 3 days postnatal, CP27 was present in the stellate reticulum, in the oral mucosa and, to a lesser degree, in the dental pulp (Figures 1E-1H). Methodological controls were negative (data not shown). Same-sized control peptide antibodies against amelogenin sequence did not react with the stellate reticulum, the alveolar bone, the oral mucosa, and the dental lamina basement membrane (data not shown). Outside of the developing tooth organ, CP27 was localized in the periphery of developing organs, including periosteum, perichondrium, and pericardium (data not shown).
In the present study we documented a tightly restricted temporospatial localization pattern for CP27 during tooth development. CP27 was highly condensed in significant areas during tooth formation, including the epithelial-mesenchymal interface of dental lamina stage tooth organs, the dental papilla, the alveolar bone, the stellate reticulum, and the oral mucosa mesenchyme. A series of control reactions including controls with similar-sized peptide antibodies against sequence fragments from other genes established the specificity of the CP27 antibody.
CP27 featured a highly distinct localization pattern in the epithelial-mesenchymal interface of dental lamina stage tooth organs. The basement membrane of the dental lamina epithelium in dental lamina stage tooth organs is a site of pivotal importance for the onset of tooth development. This basement membrane is the primary site for epithelial-mesenchymal interaction signal transduction. It transmits signals from the neural crest-derived dental mesenchyme to the oral epithelium and reciprocal signals from the tooth-forming epithelium towards the dental mesenchyme. Indirectly, these signals provide evidence for the significance of the dental lamina epithelium basement membrane as a potential site of epithelial-mesenchymal information exchange. The distinct localization of CP27 in the basement membrane of the early developing tooth organ suggests that CP27 might play an important role during the epithelial-mesenchymal interactions at the onset of tooth development.
Our results further revealed that CP27 is a constitutive element of the stellate reticulum ECM of cap and bell stage tooth organs. The stellate reticulum is a population of star-shaped cells between the dental sac and the developing tooth crown that has remarkably large intercellular spaces (Sasaki 1990). The origin, fate, and function of the stellate reticulum are not completely understood. The stellate reticulum may act to protect the developing tooth cusps or merely to define their spatial growth. Based on its close association and similarities in gene expression, the stellate reticulum may be a part of the enamel knot-based signaling center for the developing enamel organ (Vaahtokari et al. 1991), or it might be involved in calcium transport during enamel biomineralization (Sasaki 1990). On a protein level, the stellate reticulum is rich in ECM glycoproteins, including fibronectin, E-cadherin, and syndecan1 (Lesot et al. 1981; Sasaki 1990). Most recently, the stellate reticulum has been identified as a source of stem cells in the developing tooth organ (Harada et al. 1999).
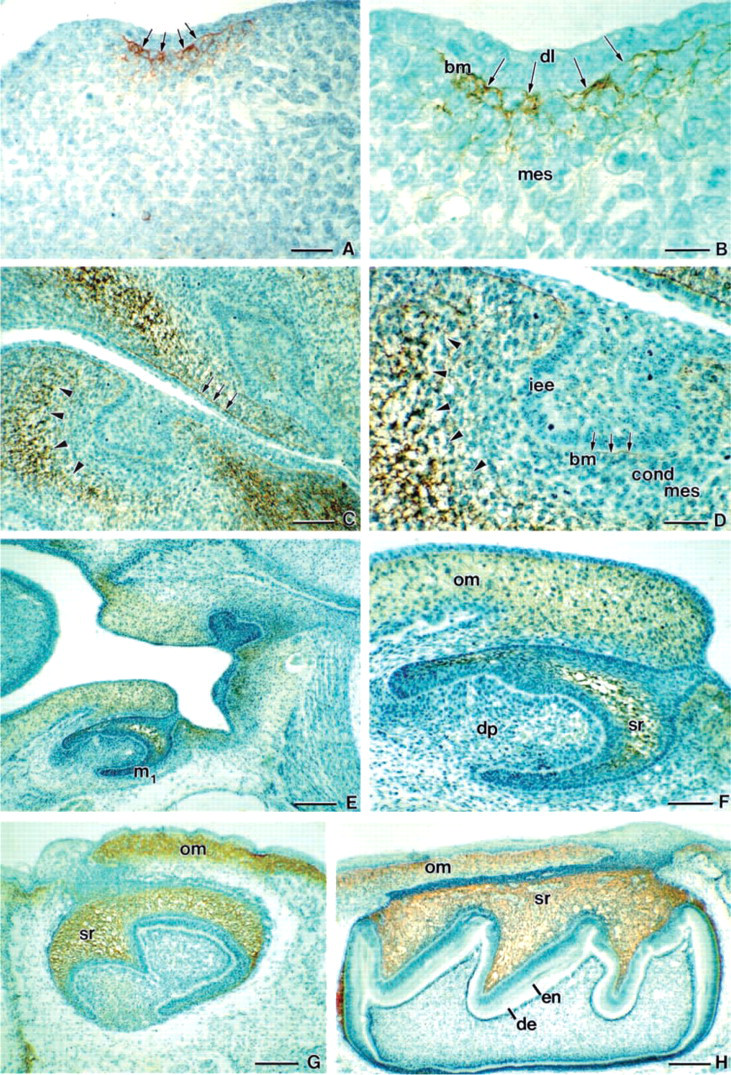
Immunolocalization of CP27 epitopes in developing teeth. During initial stages of tooth formation (E11), CP27 was localized in the basement membrane (bm) of the dental lamina (dl) and in mesenchymal cells adjacent to the dental lamina (
Our finding of a distinct localization of CP27 in the stellate reticulum of developing teeth is consistent with other findings from our group reporting the collapse of the stellate reticulum and subsequent inhibition of tooth crown formation after inhibition of CP27 function (Diekwisch and Luan in press). There are several possibile ways that CP27 may function in the stellate reticulum to facilitate tooth crown formation. CP27 may regulate ECM assembly in the stellate reticulum, including the establishment of a proteoglycan matrix that has been shown to be an essential part of the stellate reticulum (Sasaki 1990). The presence of CP27 gene products in the stellate reticulum may also play a role in the enamel knot-based signaling during tooth crown formation (Vaahtokari et al. 1991). A related study suggests (Luan and Diekwisch in press) that, on a cellular level, CP27 might exert its functions as a signaling molecule in an ECM-related pathway. The distinct localization of CP27 in the ECM and on the cell surface as revealed in the present study is definitely congruent with this hypothesis, and there is a possibility that CP27 in the stellate reticulum acts in conjunction with other matrix molecules to determine cellular behavior of this important signaling center during tooth development.
There were distinct differences between the microscopic CP27 mRNA localization as reported earlier (Diekwisch et al. 1999) and the protein tissue localization patterns reported in the present study. These differences might be explained by (a) the possibility of protein export and (b) the possibility of mRNA production in the tissues labeled by immunodetection at stages before our in situ mapping of mRNA signals. There have been a number of reports on molecules shifting their distribution patterns rapidly during early embryonic development, including Fgf-8 and Bmp-4 (Bei and Maas 1998). Although CP27 may be quite different from the growth and transcription factors involved in early embryogenesis, changes in CP27 distribution might nevertheless aid in the fulfillment of multiple functions during early embryonic development.
The significant changes in the highly restricted CP27 localization pattern suggest that CP27 might be involved in tooth formation at several different levels during development. Our immunolocalization studies have localized CP27 in the ECM of the dental lamina basement membrane and of the stellate reticulum. Our findings are consistent with CP27 function studies that establish CP27 as an essential gene product for normal tooth crown formation (Diekwisch and Luan in press) and additional studies reporting CP27 loss of function behavior similar to that of an ECM protein (Luan and Diekwisch in press). Together, these data indicate that CP27 is an ECM protein and is present in key locations during tooth development.
Footnotes
Acknowledgements
Support for these studies by NIH grant DE13095 to T.G.H.D. is gratefully acknowledged.