
Abstract
Bronchiolization is a key process in fibrosing lung in which the proliferative status of bronchiolar epithelium changes, leading to abnormal epithelial morphology. Within the context that caveolin-1 acts to suppress epithelial proliferation, we postulated that stimulating epithelial injury would lead to caveolin-1 downregulation and encourage proliferation. The present study evaluates the expression of caveolin-1, especially in bronchiolization, in C57BL/6J mice with bleomycin-induced lung fibrosis and in various types of re-epithelialization in human interstitial pneumonias (IPs). Immunohistochemically, levels of caveolin-1 decreased in the bronchiolar epithelium of mice treated with bleomycin. Levels of caveolin-1 mRNA in the whole lung were decreased at 7 and 14 days. Caveolin-1 mRNA was also decreased in laser-capture microdissection- retrieved bronchiolar epithelial cells at 7 days. Among patients with 12 IPs, including four usual IPs (UIPs) and eight nonspecific IPs (NSIPs), whole lung caveolin-1 was significantly decreased compared with 12 controls at both mRNA and protein levels. By scoring immunointensity, caveolin-1 was significantly reduced in bronchiolization and squamous metaplasia as well as in bronchiolar epithelium in 23 IPs (12 UIPs and 11 NSIPs) compared with bronchiolar epithelium from seven controls. These data suggested that loss of caveolin-1 is associated with abnormal re-epithelialization in lung fibrosis.
E
Caveolae are 50- to 100-nm vesicular invaginations of the cell surface plasma membrane that are thought to arise due to the local accumulation of cholesterol, glycosphingolipids, and the 21- to 24-kDa integral membrane protein caveolin-1, which is a principal component of caveolar membranes and plays physiological roles in intracellular vesicular transport such as transcytosis and endocytosis and signal transduction (Liu et al. 2002; Cohen et al. 2004). In normal adult lungs, caveolin-1 is preferentially located in endothelial cells, type I epithelial cells, smooth muscle cells, and fibro-blasts (Cohen et al. 2004) and is detectable in bronchiolar epithelial cells (Kasper et al. 1998; Kato et al. 2004). Targeted caveolin-1 disruption results in profound disruption of the vascular system and thickening of alveolar septa and fibrosis in the lung, accompanied by marked hypertrophy of type II epithelial cells in mice (Drab et al. 2001). Caveolin-1 is indeed downregulated in alveolar epithelium in experimental irradiation-induced lung injury (Kasper et al. 1998) and in bleomycin-induced lung fibrosis (Barth et al. 2006), implying that the loss of alveolar caveolin-1 could be an early indicator of serious type I epithelial cell injury in the process of fibrogenesis. Analysis of global gene expression using microarrays has recently revealed that caveolin-1 is downregulated in whole lung tissue of patients with idiopathic pulmonary fibrosis (IPF) (Wang et al. 2006). On the other hand, caveolin-1 overexpression prevents cell transformation (Engelman et al. 1997; Galbiati et al. 1998) and promotes cell-cycle arrest and senescence (Galbiati et al. 2001; Volonte et al. 2002), suggesting that caveolin-1 could suppress the proliferation and migration of cancer cells. The reduced expression of caveolin-1 in various human cancer cells such as breast, lung, colorectal, and ovary, as well as in sarcomas, supports this notion (Williams and Lisanti 2005; Torres et al. 2006). We thus speculated that caveolin-1 is specifically downregulated in abnormal re-epithelialization and is associated with the proliferative phenotype in lung fibrosis. For more than two decades, bleomycin-induced lung injury in experimental animals has been used as a model to investigate the cell biology and histopathology of pulmonary lesions that resemble human IPF (Thrall and Scalise 1995). Studies of this model have helped uncover the complexity of mechanisms involved in human disease. Although bleomycin may damage structural cells of the lungs directly (Simon and Paine 1995; Sato et al. 1999), its principal mode of action in leading to IPF-like pathology seems to be via endogenous mediators of inflammation, fibrinolysis, and proliferation. Here we assessed caveolin-1 expression from the perspective of bronchiolization in mouse bleomycin-induced lung fibrosis and in various types of abnormal re-epithelialization in human IPs.
Materials and Methods
Animals and Experimental Protocols
Male C57BL/6J mice (6-8 weeks old) were housed in plastic chambers with free access to food and water in a pathogen-free animal facility. After an IP injection of ketamine and xylazine for sedation and anesthesia, 0.05 U of bleomycin (Blenoxane; Nippon Kayaku, Tokyo, Japan) was intratracheally administered as described (Betsuyaku et al. 2000). After 7 and 14 days, the animals were killed, and their lungs were processed as described below. Mice that had not undergone manipulation served as controls. The Ethical Committee on Animal Research of Hokkaido University School of Medicine approved all experimental protocols and procedures associated with the study.
Tissue Processing of Mouse Lungs
Immediately after the animals were killed, the thorax was opened, and whole lungs were perfused with saline through the right ventricle to remove blood. The lungs were fixed by inflation with 10% buffered formalin for 24 hr and embedded in paraffin for immunohistochemistry (n=2-3 per time point) or inflated with diluted Tissue-Tek OCT (Sakura Finetek; Torrance, CA) and then stored frozen at −80C for RNA and protein extraction as described (n=5 per time point) (Betsuyaku et al. 2001).
Patients and Tissue Collection
The study population was comprised of 23 patients with IP, including 12 with usual interstitial pneumonia (UIP) and 11 with nonspecific interstitial pneumonia (NSIP). Patients with UIP were 65 ± 1 year old, female/male ratio was 6/6, and nine patients were smokers, whereas patients with NSIP were 59 ± 4 years, female/male ratio was 8/3, and two patients were smokers. Applying the diagnostic criteria of the American Thoracic Society/European Respiratory Society (ATS/ERS 2002) international multidisciplinary consensus classification, each diagnosis was based on the standard clinical criteria and histopathological analyses of lung tissues obtained by video-assisted thoracoscopy-guided lung biopsy or surgical lobectomy, as previously described (Odajima et al. 2006). All control lung specimens were obtained from 17 patients who had never smoked (3 males and 14 females, aged 64 ± 2 years) and who underwent lung lobectomy for small peripheral tumors. Immediately after biopsy or lobar resection, tissues were inflated with diluted Tissue-Tek OCT (Sakura Finetek) as described above, frozen as soon as possible, and stored at −80C before RNA and protein extraction (IPs, n=12 with 4 UIPs and 8 NSIPs among 23 IPs; controls, n=12) or were fixed by manual inflation with 10% neutral-buffered formalin using a 23-gauge needle and embedded in paraffin for immunohistochemistry (IPs, n=23 with 12 UIPs and 11 NSIPs; controls, n57), in which only two control lungs were overlapped for both studies.
Written informed consent was obtained from all patients, and the Ethics Committee of Hokkaido University School of Medicine approved the study. Vital capacity (VC), forced expiratory volume in 1 sec (FEV1), and diffusing capacity for carbon monoxide (DLCO) were measured in all patients (CHESTAC-55V; Chest Co., Tokyo, Japan) before surgery. All patients also underwent high-resolution computed tomography scans, arterial blood gas analysis, and serum Krebs von den Lungen-6 (KL-6) measurements (Yokoyama et al. 1998).
Immunohistochemistry
Caveolin-1 immunohistochemistry proceeded as described (Betsuyaku et al. 2003b) using a CSA kit (Dako Japan; Kyoto, Japan) according to the manufacturer's protocol. Tissue sections were incubated with a rabbit anti-caveolin-1 antibody (N20 sc894; Santa Cruz Biotechnology, Santa Cruz, CA) diluted 1:1000 for mice and 1:2000 for humans overnight at 4C. Sections were counterstained with hematoxylin. Staining of endothelial cells and lymphocytes served as internal positive and negative controls, respectively, for caveolin-1. Sections were stained for Ki-67 with a mouse anti-Ki-67 antibody (clone MIB-1; Dako Japan) and 0.1 mM EDTA buffer (pH 8.0) for antigen retrieval in an automated immunostainer (Ventana NX; Ventana Japan, Yokohama, Japan).
Semiquantitative Immunohistochemistry for Caveolin-1 in Human Lungs
Because the amount and distribution of abnormal epithelial cells within the lungs was heterogeneous, immunostained tissue sections from one block per patient were thoroughly investigated to identify more than one group of each cell type. Staining intensity of caveolin-1 expression was then scored for each cell type at the same magnification using a visual scoring method with grades ranging from 0 to 3 (0, no staining; 1, moderate staining; 2, intense staining; 3, very intense staining), as described (de Boer et al. 2000; Odajima et al. 2006). Intensity was scored twice in a blinded manner, in which the observer (NO) was unaware of any clinical details of the patients. Based on significant Spearman's rank correlation coefficients between two observations for caveolin-1 (p<0.0001), only the latter data set is presented.
Laser-Capture Microdissection (LCM) of Bronchiolar Epithelial Cells in Mouse Lung
Bronchiolar epithelial cells were selectively harvested from the lungs by LCM using a PixCell II System (Arcturus Engineering; Mountain View, CA) as described (Betsuyaku et al. 2001). After the samples were captured on transfer films (CapSure Macro LCM Caps, LCM0211; Arcturus Engineering), nonspecific attached components were removed using adhesive tape (CapSure Cleanup Pad, LCM0206; Arcturus Engineering). Bronchiolar epithelial cells were retrieved from the junction of the terminal bronchioles and alveolar ducts and proximally along airways of up to ∼250 μm in diameter, as described (Betsuyaku and Senior 2004). At least six bronchioles were randomly selected, and cells were collected from each mouse specimen using a total of 5000 laser bursts. We analyzed mRNA in non-stained serial sections (whole lung tissue specimens) adjacent to the sections examined by LCM.
Quantitative Reverse Transcriptase-Polymerase Chain Reaction
Total RNA was extracted using the RNeasy Mini Kit (Qiagen; Hilden, Germany). cDNA templates were synthesized using reverse transcriptase (RT) (Applied Biosystems; Foster City, CA), and mRNA levels were quantified by the 5′-exonuclease-based fluorogenic polymerase chain reaction (PCR) using a 7300 Real Time PCR System (Applied Biosystems) as described (Betsuyaku et al. 2001, Betsuyaku and Senior 2004), with TAKARA master mix (TAKARA BIO INC.; Shiga, Japan) according to the manufacturer's instructions. The relative amount of target mRNA in the samples was assessed by interpolating threshold cycles from a standard curve. TaqMan Gene Expression Assays probes were Hs00184697_m1 for human caveolin-1 and Mm00483057_m1 for murine caveolin-1 (Applied Biosystems), and the levels were normalized against glyceraldehyde-3-phosphatase-dehydrogenase mRNA(human) and β2-microglobulin mRNA (mouse).
Western Blotting
Frozen lung tissues were homogenized in lysis buffer (T-PER Tissue Protein Extraction Reagent; Pierce Biotechnology, Rockford, IL) containing the protease inhibitors phenylmethylsulfonyl fluoride (100 mM), antipain (1.2mg/ml), aprotinin (2 mg/ml), pepstatin A (0.5 mg/ml), and leupeptin (1 mg/ml). After the samples were centrifuged at 10,000 × g for 5 min at 4C, the total protein concentration of the supernatant was determined using the BCA method (Pierce Biotechnology). Mouse and human samples (15 and 10 μg of protein, respectively) were resolved by electrophoresis under reducing conditions as described (Betsuyaku et al. 2003a). Transferred membranes were incubated overnight at 4C with rabbit anti-caveolin-1 antibody (N20 sc-894) diluted 1:2000 followed by horseradish peroxidase-conjugated anti-rabbit immunoglobulin (Dako Japan) diluted 1:20,000. Membranes were treated with the Immobilon Western Chemiluminescent HRP Substrate (Millipore Corporation; Billerica, MA) and then specific antibody binding was visualized by exposure to photographic film. Because the use of β-actin as a normalizing control is limited in human lung diseases (Glare et al. 2002), loading homogeneity was determined based on an equal amount of total protein in each sample. Immunoreactive signals were analyzed using an Epson 2000 Scanner (Nagano, Japan) and calculated using Scion Image software (Scion Corporation; Frederick, MD).
Statistical Analysis
Results are expressed as means ± SEM. Data were statistically analyzed using the single factor ANOVA, and Student's unpaired t-test was applied to comparisons between groups. Two groups of immunohistochemical results were compared using the Mann-Whitney U test, and differences in the immunointensity of different epithelial cell types between IPs were analyzed using the Kruskal-Wallis test. Spearman's rank correlation coefficients were calculated to assess test-retest reliability (StatView J 5.0; SAS Institute Inc., Cary, NC); p<0.05 was considered statistically significant.
Results
Localization of Caveolin-1 in Normal and Fibrotic Lungs in Mice
We immunohistochemically localized caveolin-1 in normal and fibrotic mouse lungs. Caveolin-1 was ubiquitously expressed in type I cells and endothelial cells in the normal lungs (Figure 1A) and remained in those cells in alveolar walls even in the damaged lungs induced by bleomycin (Figures 1B-1F). Caveolin-1 was also detected at the apical side of some bronchiolar epithelial cells (Figure 1A), whereas bronchiolar caveolin-1 was barely detected in these cells at 7 days after bleomycin treatment (Figure 1B). At 14 days, caveolin-1 was negative in bronchiolar epithelial cells and in bronchiolization within fibrotic areas (Figures 1C and 1D). Alveolar bronchiolization was identified as cells resembling bronchiolar epithelium lining normal or thickened alveolar walls, often in an acinar formation, as previously described (Betsuyaku et al. 2000). Caveolin-1 was also barely detected in fibroblastic foci (Figures 1E and 1F).
Caveolin-1 mRNA and Protein in Fibrotic Lung in Mice
Whole lung caveolin-1 mRNA expression was significantly decreased in mice at 7 and 14 days after intratracheal bleomycin administration compared with controls (0.6 ± 0.1 and 0.7 ± 0.1, respectively, vs 1.5 ± 0.1, p<0.01) (Figure 2A). Figure 2B shows a representative Western blot of caveolin-1 protein at 22 kDa in whole lung homogenates. Although caveolin-1 protein levels were decreased after bleomycin treatment, differences did not reach statistical significance at 7 days and 14 days after bleomycin treatment compared with untreated controls (3.0 ± 1.5 AU vs 7.8 ± 2.0, p=0.05; 3.8 ± 1.3 vs 7.8 ± 2.0, NS, respectively) (Figure 2C).
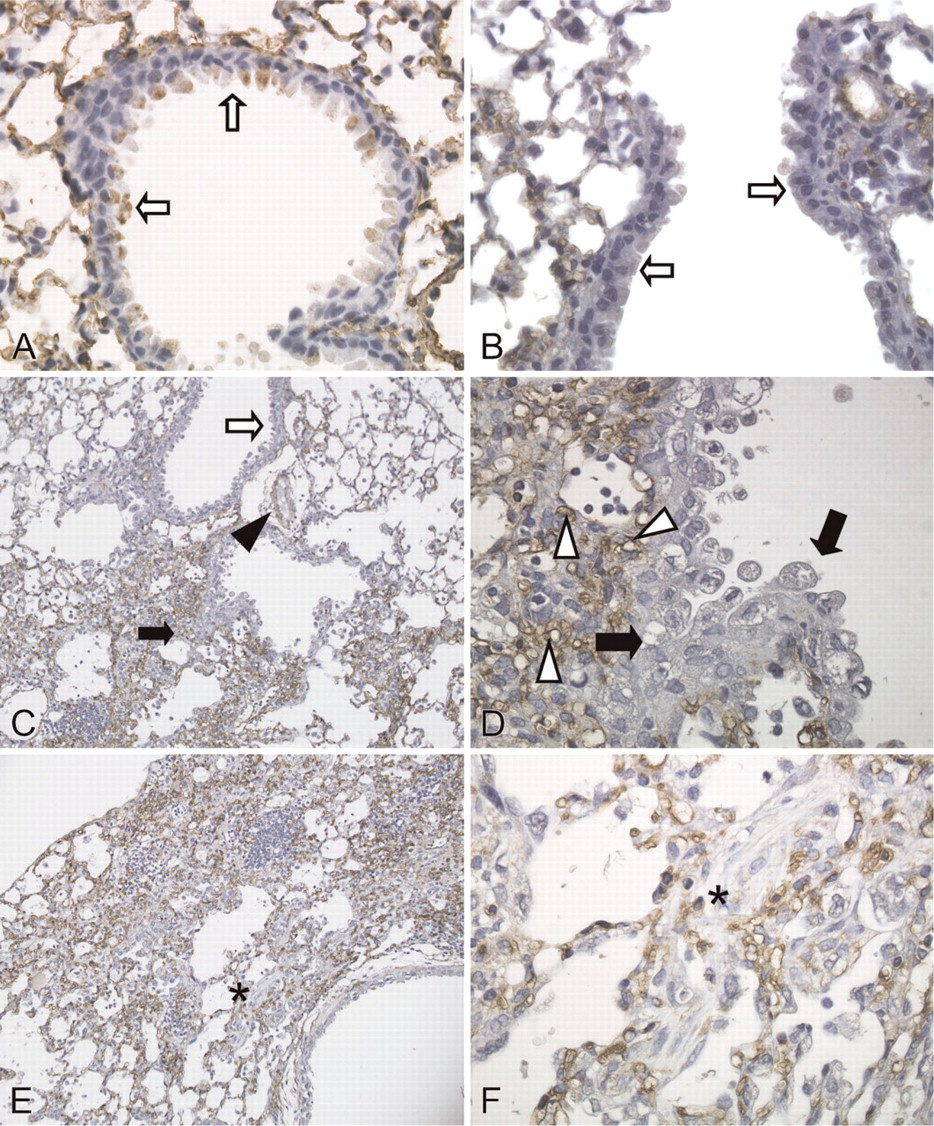
Immunohistochemical localization of caveolin-1 in mouse lungs. (
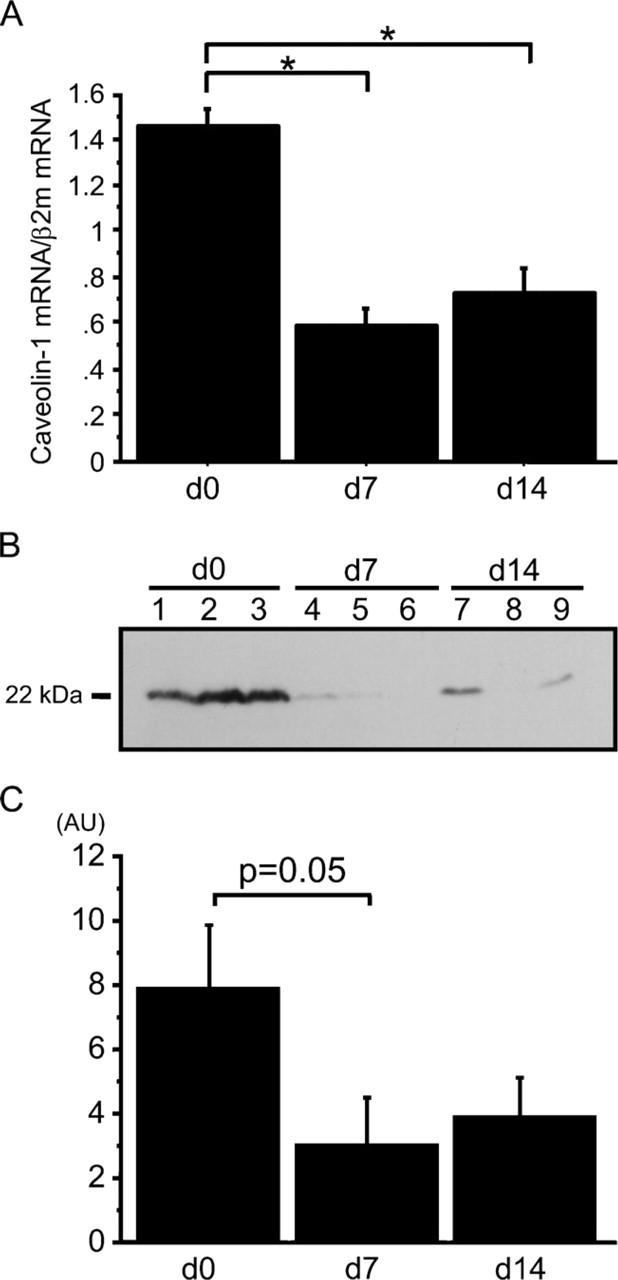
Expression of caveolin-1 mRNA and protein in whole lungs of mice. d0, untreated control mice; d7, 7 days after bleomycin treatment; d14, 14 days after bleomycin treatment (n=5 for each point). (
Caveolin-1 mRNA in LCM-retrieved Bronchiolar Epithelial Cells in Mice
We harvested bronchiolar epithelial cells from the lungs using LCM to quantify caveolin-1 mRNA expression in vivo. Figure 3 shows the selective retrieval of terminal bronchiolar epithelium from the mouse lung 7 days after intratracheal bleomycin using LCM. After bleomycin administration, numerous inflammatory cells appeared in the interstitium and additional collagen was deposited around bronchoalveolar junctions (Figure 3A). The tissue retrieved by LCM was confined to terminal bronchiolar epithelium (Figures 3B and 3D), and the tissue remaining after LCM further demonstrated the selectivity of this procedure for removing terminal bronchiolar epithelium (Figure 3C). LCM was not applicable at 14 days after bleomycin exposure. Fibrous deposition at that time surrounded the bronchiolar epithelial cells, thus requiring a more intense or longer laser pulse, either of which enlarged the field and resulted in the retrieval of additional cell types. Caveolin-1 mRNA was detected in bronchiolar epithelial cells retrieved using LCM, and expression levels were significantly decreased at 7 days after bleomycin administration compared with normal lungs (0.3 ± 0.1 vs 0.6 ± 0.1, p<0.01) (Figure 4).
Patient Characteristics
Table 1 summarizes the clinical characteristics of patients with IP. Mean interval between onset of symptoms and pathological diagnosis was 13.7 months (UIP, 15.3 months; NSIP, 20.1 months). Two NSIP patients were medicated with oral corticosteroids. None of the patients or normal controls had received any drugs that might cause drug-induced pneumonitis at the time of this study. Vital capacity (% pred) and DLCO (% pred) in IP patients were lower than normal controls. Age, pulmonary function tests, and serum arterial blood gas values did not differ significantly between patients with UIP or NSIP.
Caveolin-1 mRNA and Protein in Human IPs
We evaluated caveolin-1 mRNA in whole lung homogenates using quantitative RT-PCR. Whole lung caveolin-1 mRNA was significantly decreased in IP patients (n=12) including four UIPs and eight NSIPs compared with normal controls (n=12) (0.5 ± 0.1 vs 1.1 ±0.1, p<0.01) (Figure 5A). Consistent with the mRNA results, Western blotting showed that the protein levels of caveolin-1 in whole lung were significantly decreased in patients with IP (n=12) including 4 UIPs and 8 NSIPs, compared with normal controls (n=12) (2.7 ± 1.0 AU vs 9.7 ± 1.2, p<0.01) (Figures 5B and 5C).
Localization of Caveolin-1 in Human IPs
Caveolin-1 staining was intense in type I epithelial, endothelial, and smooth muscle cells as well as in the apical side of bronchiolar epithelial cells of normal lungs (Figure 6A), but not in type II epithelial cells. In lungs of all patients with NSIP, alveolar walls were thickened with edema, fibrosis, inflammatory cell infiltration, and abnormal re-epithelialization. Epithelial cells lining bronchioles within fibrotic areas of IP (Figure 6B), along with areas of abnormal re-epithelialization such as squamous metaplasia (Figure 6C) and non-ciliated (Figure 6D) and ciliated (Figure 6E) bronchiolization were slightly stained for caveolin-1. Ki-67 staining also demonstrated that the rate of proliferation was increased in bronchiolized epithelial cells (see inset in Figure 6D). In addition, caveolin-1 was barely detectable in fibroblastic foci (Figure 6F) as well as in alveolar macrophages and lymphocytes in IPs. In lungs of all patients with UIP, fibrotic zones showed temporal heterogeneity with dense acellular collagen, scattered fibroblastic foci with intervening nearly normal alveoli, honeycombing with complete destruction of the architecture, and abnormal re-epithelialization. Distribution of caveolin-1 in UIP was similar to that of NSIP. Immunoreactive intensity of caveolin-1 was scored for bronchiolar epithelial cells and each type of abnormal re-epithelialization in each patient (Table 2). Caveolin-1 scores did not differ significantly among the types of epithelial cells between UIP and NSIP. Therefore, we combined the UIP and NSIP scores for further analysis. Because there were no differences in staining scores for caveolin-1 between non-ciliated and ciliated bronchiolization (0.4 ± 0.2 vs 0.3 ± 0.1, NS), we averaged the staining score for caveolin-1 in bronchiolization for both cell types. The score was significantly lower in bronchiolization, squamous metaplasia, and bronchiolar epithelial cells in fibrotic areas when compared with bronchiolar epithelium in control lungs (n=7) (0.4 ± 0.1, 0.1 ± 0.1, and 0.4 ± 0.1, respectively, vs 1.9 ± 0.3, p<0.01) (Figure 7). Caveolin-1 expression among bronchiolization, squamous metaplasia, and bronchiolar epithelium did not differ significantly in IP patients.
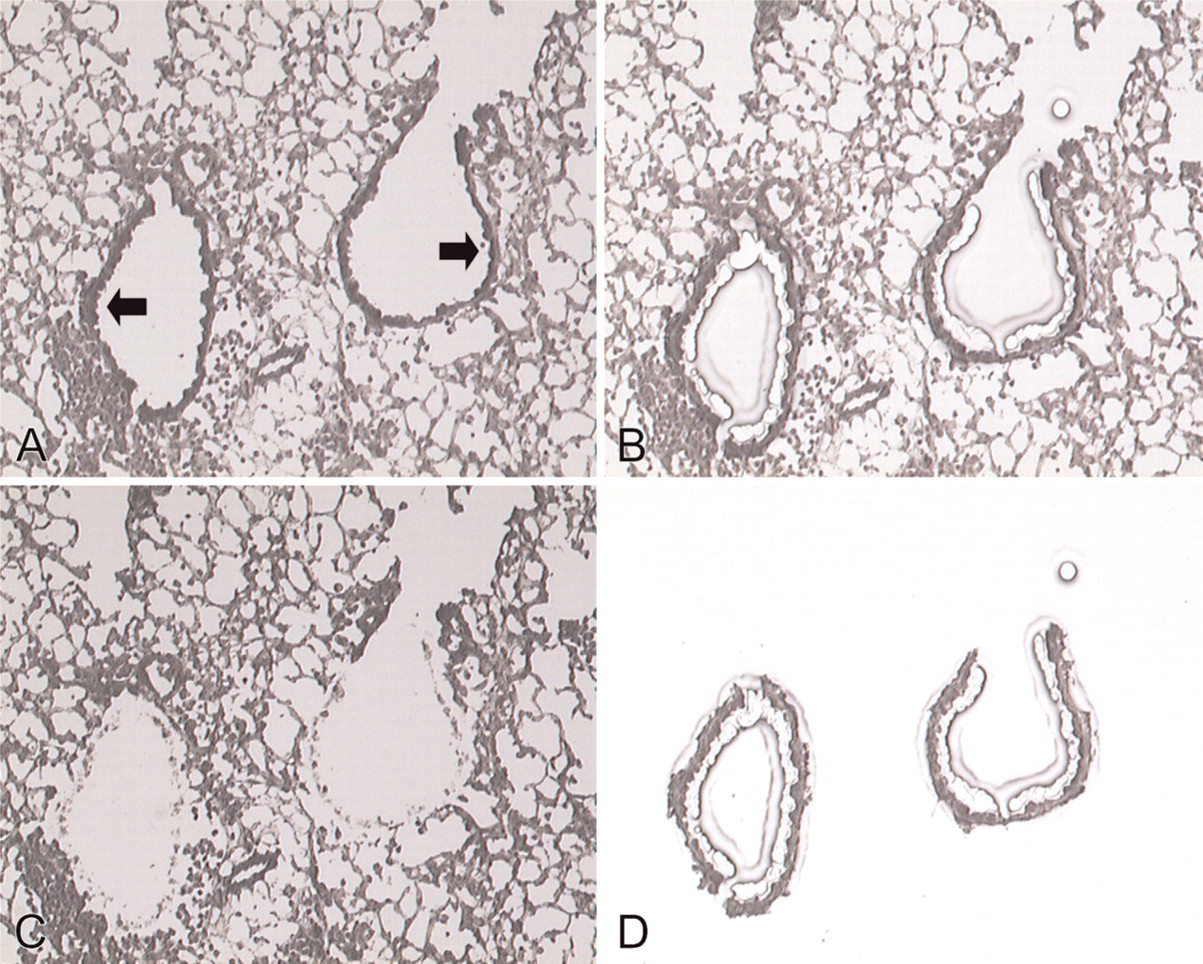
Laser-capture microdissection (LCM) of bronchiolar epithelial cells from bleomycin-treated lung at 7 days. (
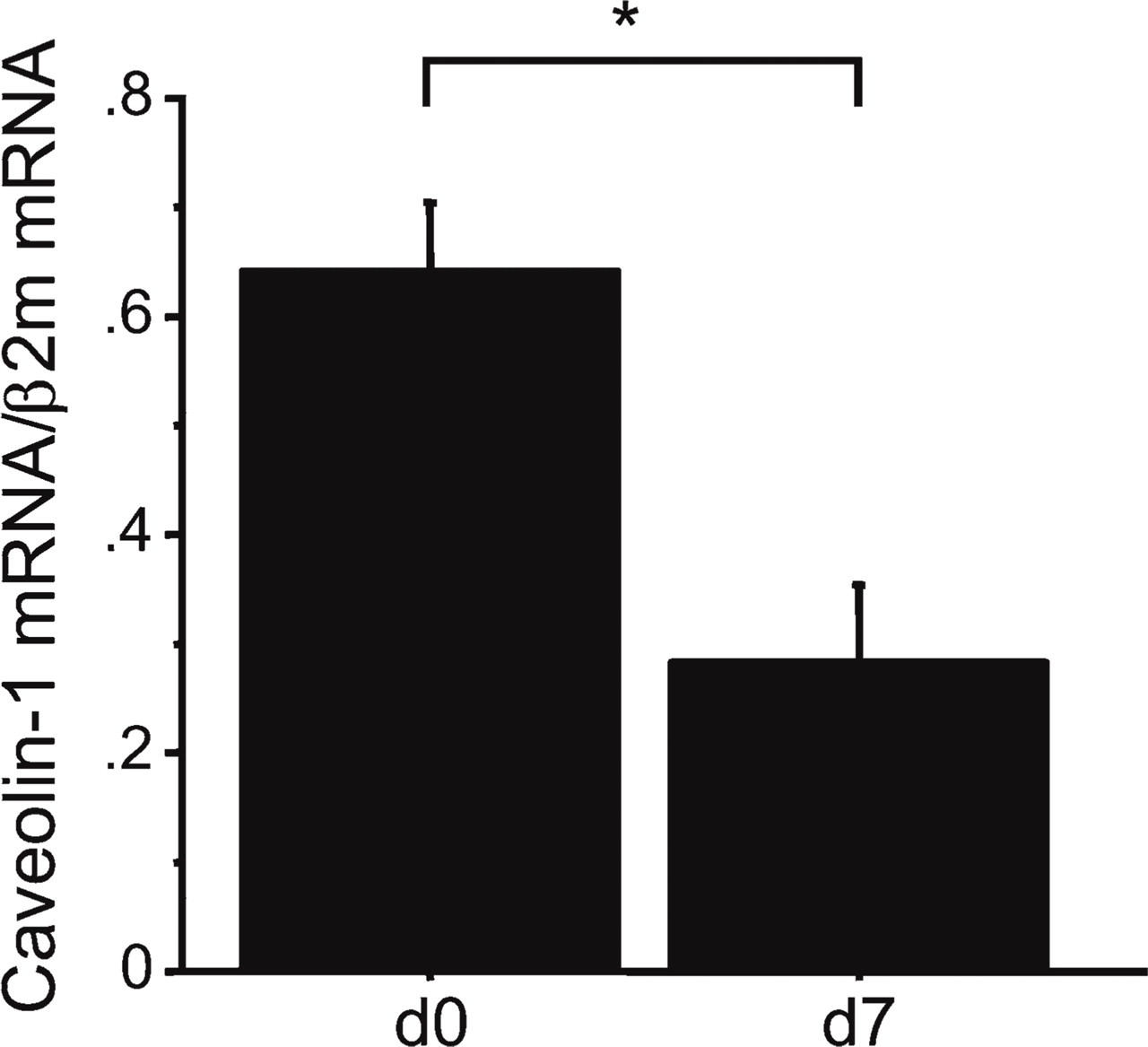
Expression of caveolin-1 mRNA in mouse bronchiolar epithelial cells retrieved using LCM Caveolin-1 mRNA in bronchiolar epithelial cells retrieved using LCM was significantly decreased compared with these cells in untreated lungs (n=5 for each point; (∗p<0.01 compared with untreated lung). β2m, β2-microglobulin; AU, arbitrary units.
Discussion
We demonstrated in this study that levels of caveolin-1 decrease in bleomycin-induced lung injury in mice and in human IPs. Furthermore, we found that caveolin-1 is lost in the aberrant re-epithelialization observed in fibrotic regions in mice and humans.
Cell renewal in injured bronchioles and alveoli comprises diversified repair mechanisms, and whether the appearance of abnormal epithelial cells protects against or promotes fibrosis is uncertain (Portnoy and Mason 2004). We previously reported that gelatinase B [matrix metalloproteinase (MMP)-9] is induced in bronchiolar epithelial cells and in bronchiolization in bleomycin-induced lung fibrosis, and that a gelatinase B deficiency results in defects in alveolar bronchiolization (Betsuyaku et al. 2000). We also reported that expression of the extracellular matrix metalloproteinase inducer (EMMPRIN) increases in bronchiolization in mouse (Betsuyaku et al. 2003a) and in human IP (Odajima et al. 2006). These data imply that EMMPRIN affects re-epithelialization via its capacity to induce MMP expression, leading to bronchiolar epithelial cell migration and proliferation. Excessive caveolin-1 is known to negatively regulate glycosylation and cell surface clustering of EMMPRIN (Tang et al. 2004; Tang and Hemler 2004), which might consequently impair MMP induction. Thus, we speculate that loss of caveolin-1 in bronchiolar epithelial cells might contrarily facilitate glycosylation and self-aggregation of EMMPRIN, leading to excessive MMP induction in those cells in lung fibrosis.
Clinical characteristics of IP patients
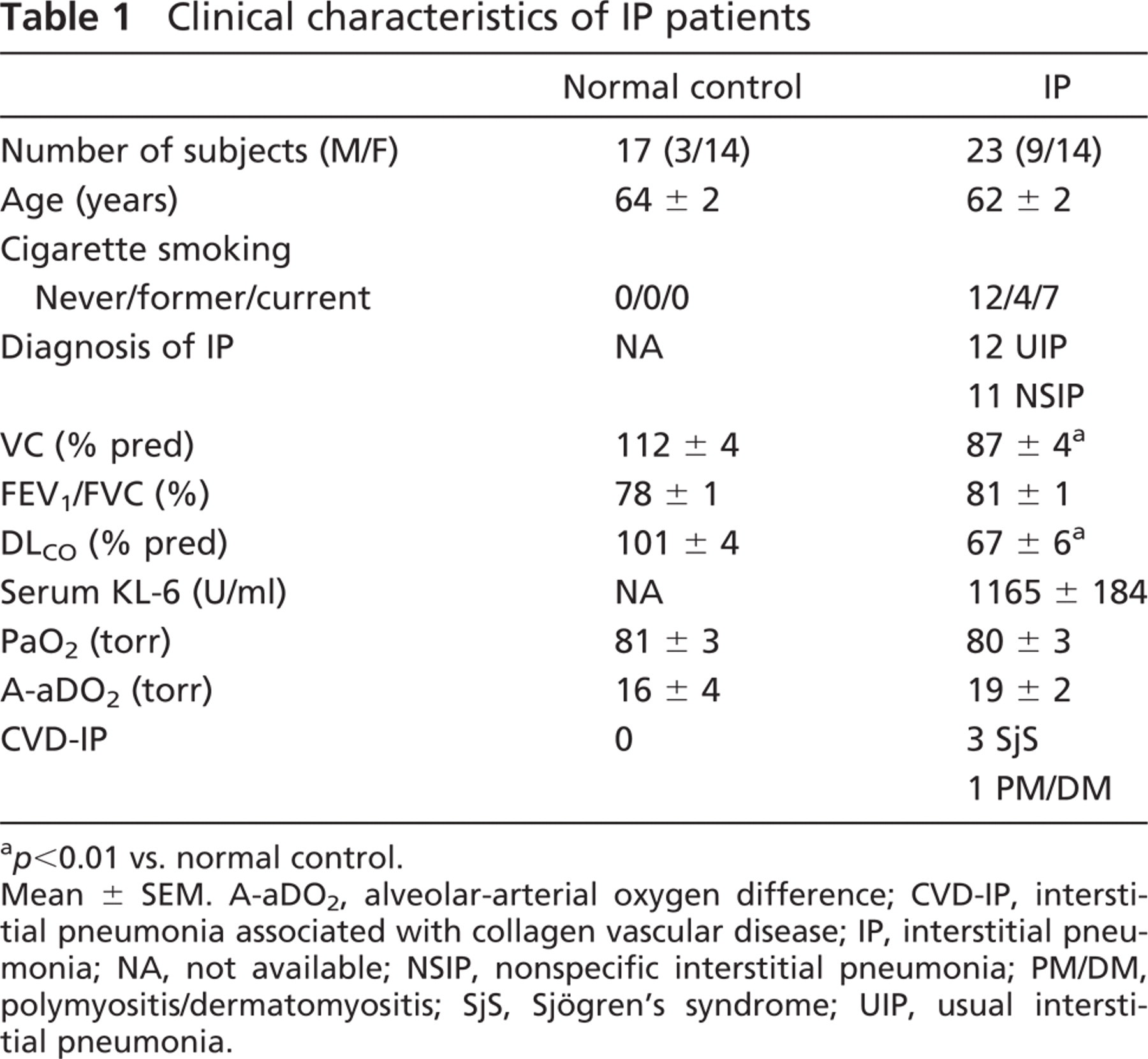
p<0.01 vs. normal control.
Mean ± SEM. A-aDO2, alveolar-arterial oxygen difference; CVD-IP, interstitial pneumonia associated with collagen vascular disease; IP, interstitial pneumonia; NA, not available; NSIP, nonspecific interstitial pneumonia; PM/DM, polymyositis/dermatomyositis; SjS, Sjögren's syndrome; UIP, usual interstitial pneumonia.
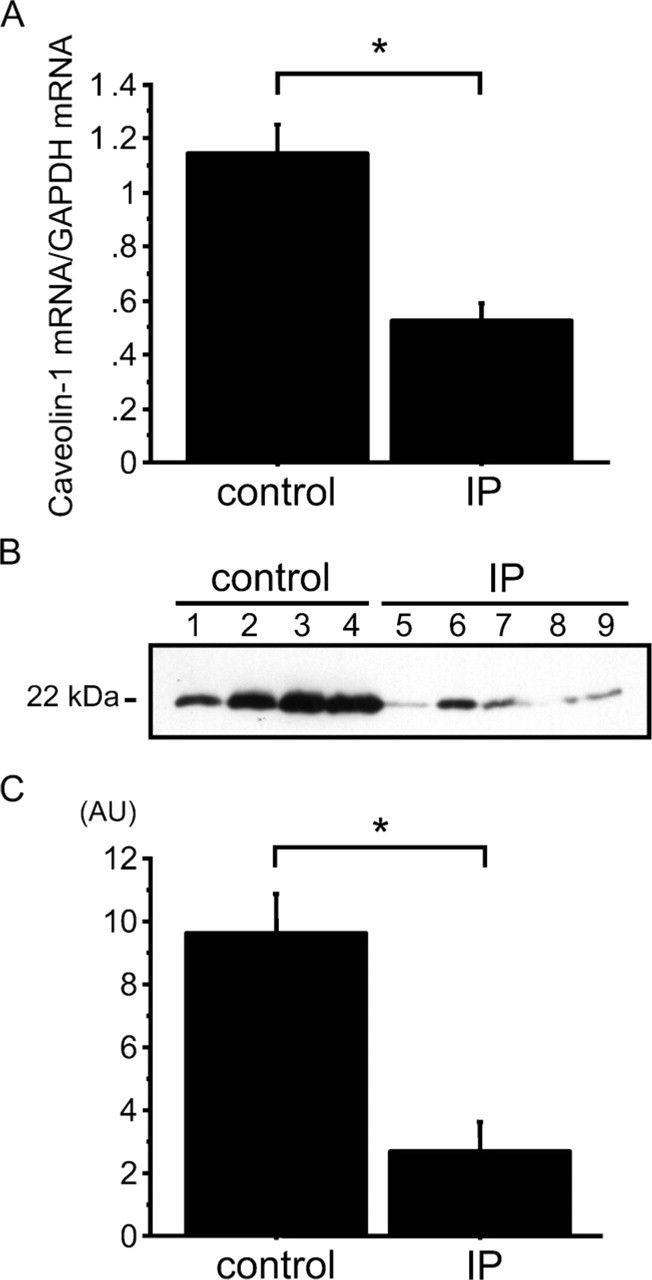
Expression of caveolin-1 mRNA and protein in human lungs. (
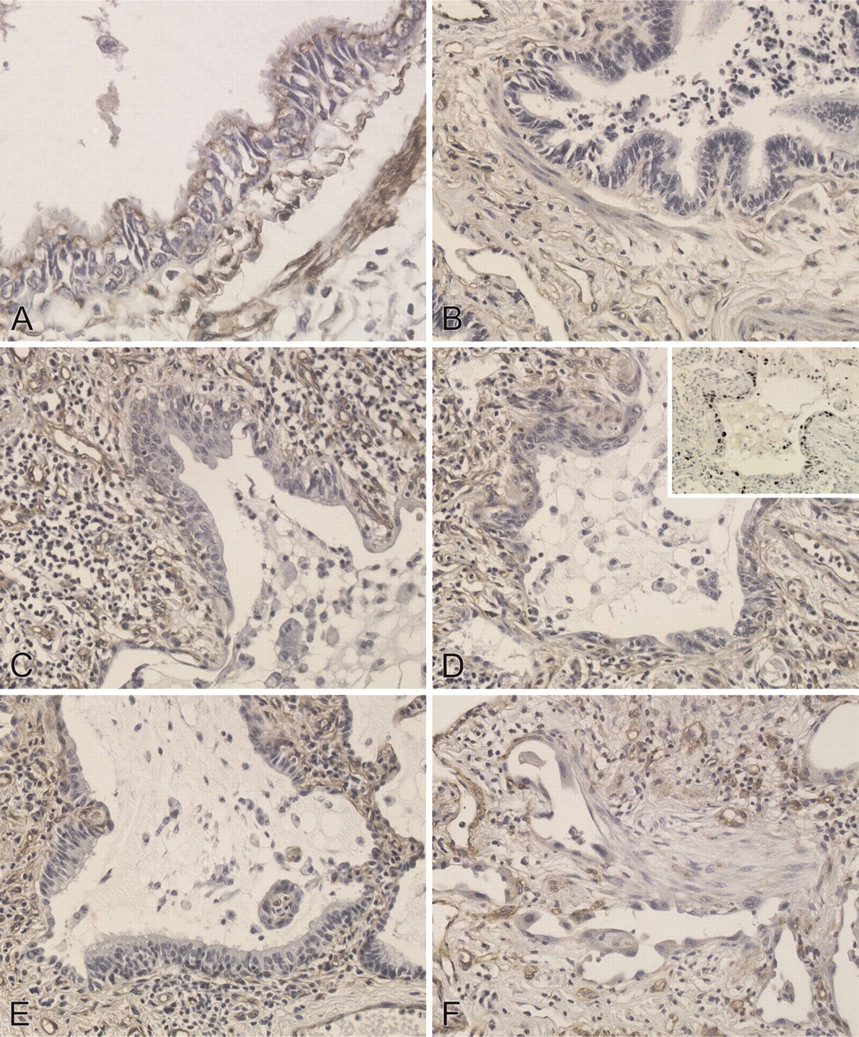
Immunohistochemical localization of caveolin-1 in human lungs. (
Immunohistochemical staining scores for caveolin-1 in bronchiolar epithelium and re-epithelialization in IP patients
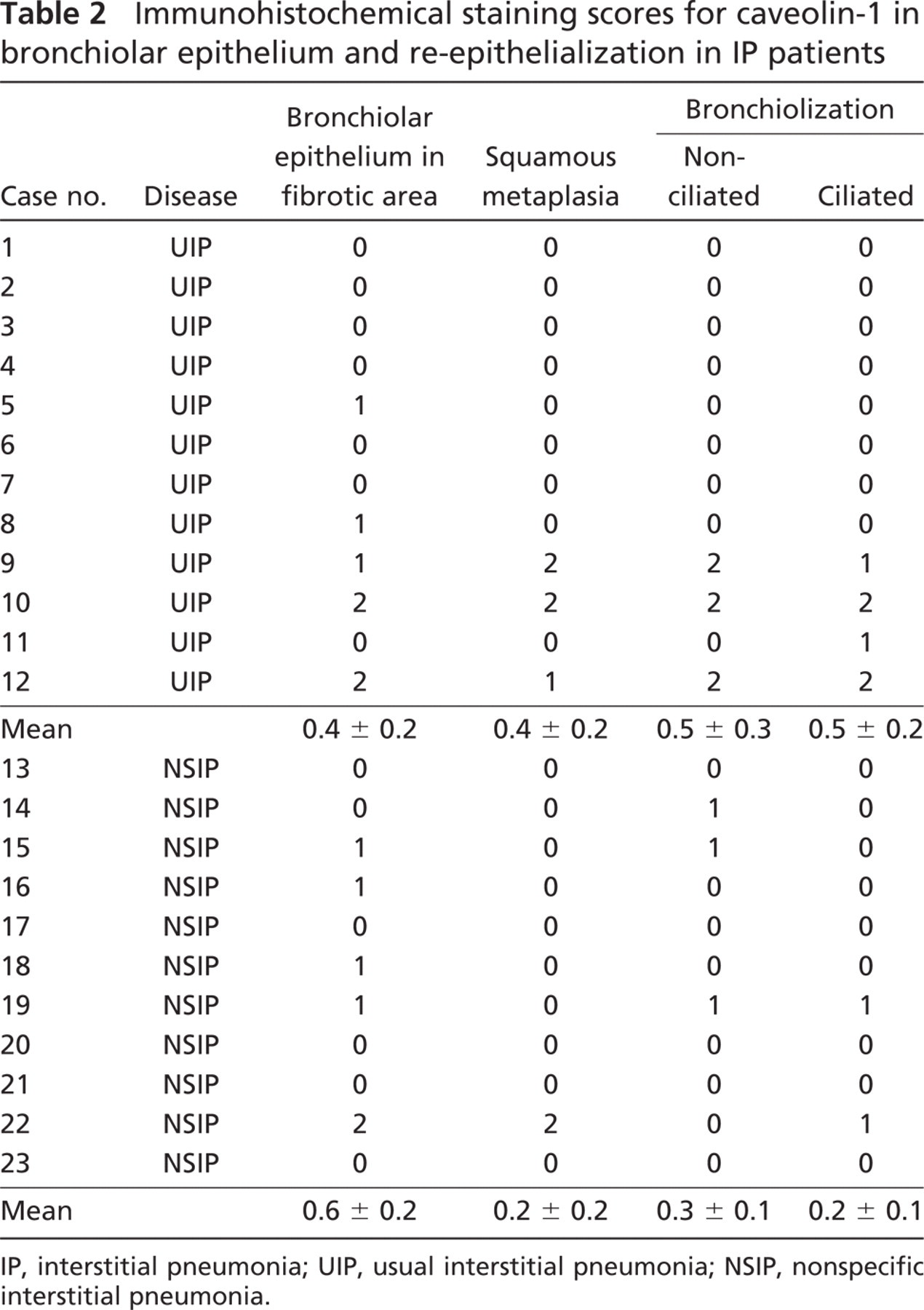
IP, interstitial pneumonia; UIP, usual interstitial pneumonia; NSIP, nonspecific interstitial pneumonia.
The role of caveolin-1 in airway epithelium has been ignored, partly because of inconsistent identification; caveolin-1 has been detected in bronchiolar epithelial cells by immunohistochemistry in some studies (Kasper et al. 1998; Kato et al. 2004), but not in others (Kogo et al. 2004; Hnasko and Ben-Jonathan 2005; Krasteva et al. 2006). Although these discrepancies have not been explained, the present study demonstrated the steady-state expression of caveolin-1 in bronchiolar epithelial cells at both mRNA and protein levels.
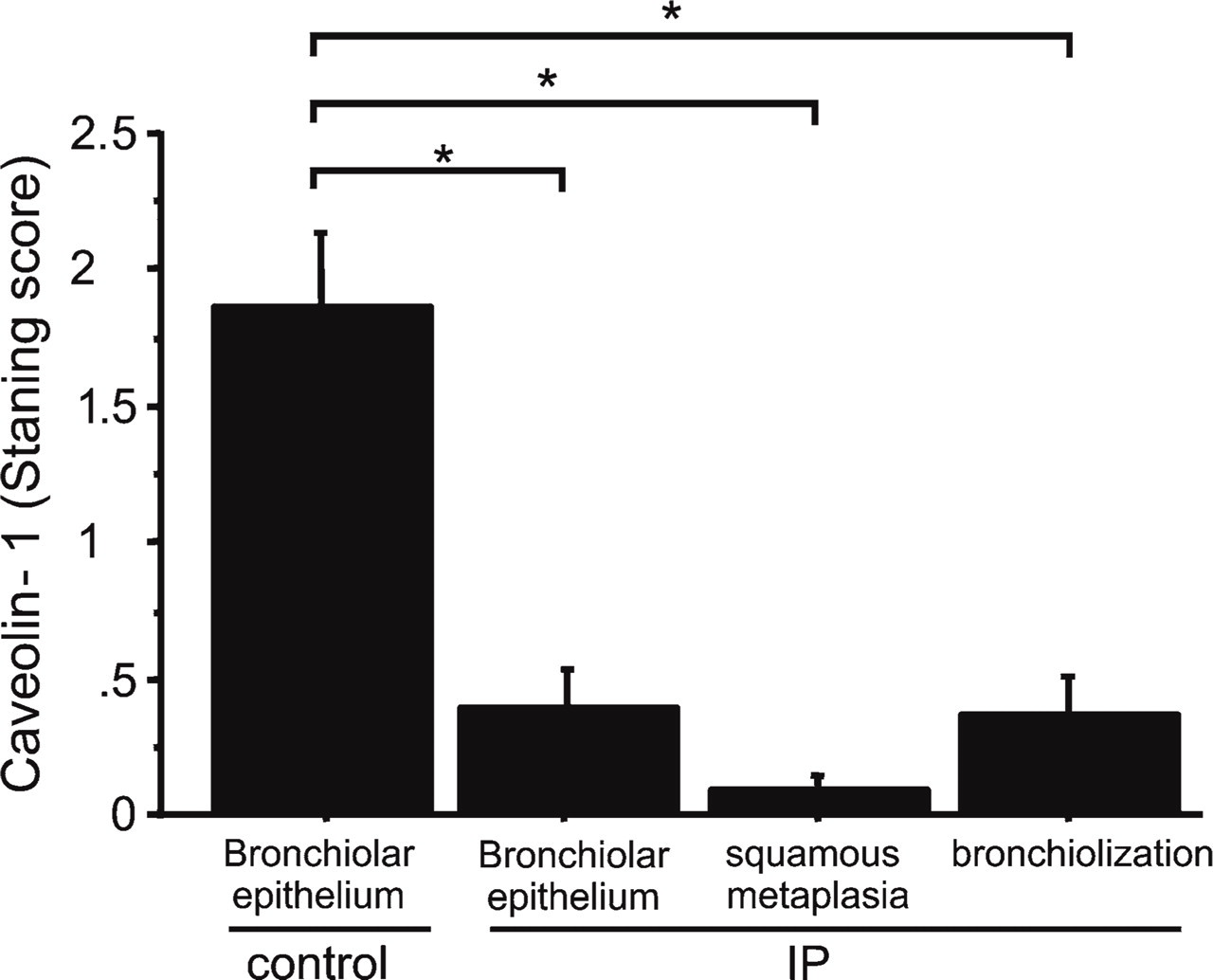
Immunohistochemical staining scores for caveolin-1. Scores for caveolin-1 in bronchiolar epithelium or each type of re-epithelialization is significantly lower in IP lungs (n=23) compared with bronchiolar epithelium in control lungs (n=7) (∗p<0.01).
Fibroblastic foci did not express caveolin-1 in the fibrotic lungs of mice and humans in this study, which agrees with the recent finding that fibroblasts harvested from IPF lungs express less caveolin-1 than those from normal lungs (Wang et al. 2006). They also showed that caveolin-1 could suppress transforming growth factor β1-induced extracellular matrix production in cultured fibroblasts. The loss of caveolin-1 in either epithelial or stromal cells independently contributes to the development of mammary hyperplasia and mammary cell transformation (Williams et al. 2006). Together the decrease of caveolin-1 at mRNA and protein levels in whole lung homogenate was probably due to the loss of caveolin-1 in bronchiolization and in fibroblasts as well as in type I epithelial cells (Kasper et al. 1998; Koslowski et al. 2004). The differences at protein levels were detectable in the human specimens but were not statistically significant in the mouse model. Mouse samples are whole lung homogenates in which the fibrosis is sparsely distributed, whereas the human tissue specimens are specifically driven from fibrotic regions. Thus, the decrease of caveolin-1 limited in fibrotic areas might be masked in mouse lung homogenates.
Caveolin-1 suppresses the proliferation and migration of various types of cancer cells in vitro and in vivo (Cohen et al. 2004; Torres et al. 2006). The scaffolding domain of caveolin-1 is a module that inhibits the growth stimulatory activity of several signaling molecules such as tyrosine and serine/threonine kinases, including epidermal growth factor receptor (Couet et al. 1997), mitogen-activated protein kinase, and some of their downstream components (Razani et al. 2002). Caveolin-1 also interacts with and inactivates some factors in the prosurvival phosphatidylinositol 3-kinase/ Akt pathway and platelet-derived growth factor receptor (Liu et al. 1996). It was reported that these signaling molecules are upregulated and associated with lung fibrosis in murine epithelial cells exposed to asbestos (Ramos-Nino et al. 2003). Together the loss of caveolin-1 in bronchiolar epithelial cells in lung fibrosis might progress bronchiolization via the signal transduction of these molecules.
In tumor cells, downregulation of caveolin-1 leads to the loss of E-cadherin, increases transcriptional activity of β-catenin, causes epithelial-to-mesenchymal transition (EMT), and enhances tumor invasion (Lu et al. 2003). In lung fibrosis, β-catenin is reportedly accumulated in nuclei of bronchiolar epithelial cells as well as in regenerated epithelial cells (Chilosi et al. 2003). We then speculate that the downregulation of bronchiolar caveolin-1 might be associated with β-catenin activation, leading to aberrant re-epithelialization and/or EMT in lung fibrosis.
In summary, we demonstrated a significant decrease of caveolin-1 expression, especially in bronchiolar epithelium and in abnormal re-epithelialization in fibrotic lungs.
Footnotes
Acknowledgements
We thank Ms. Yoko Suzuki for technical assistance with laser-capture microdissection.