
Abstract
Gangliosides and glycosylphosphatidylinositol (GPI)-anchored proteins contain lipid tails that tether them to the outer side of the cell membrane. This mode of association with the cell membrane enables them to take part in the organization of lipid rafts, but it also permits gangliosides and GPI-anchored proteins to be actively released from one cell and inserted into the membrane of another cell. Recently, we reported that under conditions of lipid raft isolation, Triton X-100 causes significant redistribution of both gangliosides and GPI-anchored proteins. Aiming to find a less disruptive detergent, we evaluated the effects of CHAPS, Saponin, deoxycholic acid, Trappsol, Tween 20, Triton X-100, Brij 96V, Brij 98, and SDS on brain tissue sections. At room temperature, all detergents (1% concentration) extracted significant amounts of both gangliosides and Thy-1. At 4C, the extraction was weaker, but Triton X-100, CHAPS, and deoxycholic acid caused significant redistribution of GD1a and Thy-1 from gray matter into the white matter. Both redistribution and extraction were significantly augmented when sections were incubated with detergents in the presence of primary antibodies. Of the nine tested detergents, none is the ideal choice. However, Brij 96V appears to be able to sufficiently reveal myelin epitopes while causing the least amount of artifacts. This manuscript contains online supplemental material at http://www.jhc.org. Please visit this article online to view these materials.
Keywords
G
Patterns of ganglioside expression change with cell growth and differentiation, viral transformation, ontogenesis, and oncogenesis (Hakomori 1981). In the brain, gangliosides are expressed in cell-type and developmentally specific patterns (Rösner 1993, 2003; Kotani et al. 1995; Heffer-Lauc et al. 1996, 1998; Schwarz and Futerman 1996; Letinic et al. 1998). Immunohistochemical analysis of ganglioside distribution has been a challenge for decades. Like many other glycan structures, gangliosides are highly conserved during evolution and are consequently very poor antigens for the production of antibodies (Kawashima et al. 1992; Portoukalian 2000). Various approaches have been attempted over the past years, and the problem appears to have been finally resolved by raising antibodies in mice deficient for complex gangliosides (Schnaar et al. 2002). Numerous studies of ganglioside distribution have been published [reviewed by Schwarz and Futerman (1996)], but some of the results are contradictory, and many issues remain to be resolved. For example, some authors have reported that ganglioside GD1a is present intensively in the molecular layer and very weakly in the granular layer of rat cerebellum (Kotani et al. 1993), whereas other authors have reported exactly the opposite distribution, with strong staining of the granular layer and weak staining of the molecular layer (Lunn et al. 2000). An additional challenge is the fact that the analysis of some molecules, such as MAG (myelin-associated glycoprotein, a putative receptor for GD1a), requires the use of detergents (Bo et al. 1995; Wang et al. 2002). Because detergents augment the effect of shedding and uptake, the use of detergents significantly impedes concurrent analysis of molecules such as MAG and gangliosides or GPI-anchored proteins.
Triton X-100 is among the most commonly used detergents for the analysis of membrane antigens, but we recently reported that its presence during an immunostaining procedure caused significant redistribution of gangliosides and GPI-anchored proteins (Heffer-Lauc et al. 2005). Aiming to find amore suitable detergent, we compared nine different detergents, and here we report their effects on immunohistochemical analysis of brain tissues at room temperature and at 4C.
Materials and Methods Materials
Mice lacking complex brain gangliosides (including GM1 and GD1a) were generated by disrupting B4 galnt1 (previously known as Galgt1), the gene that encodes UDP-GalNAc:GM3/GD3 N-acetylgalactosaminyltransferase (B4 galnt1-null mice) (Liu et al. 1999). The genotypes of offspring were determined using PCR as described (Sheikh et al. 1999). Adult male C57BL/6 mice (Charles River Laboratories; Wilmington, MA) and B4 galnt1-null mice were anesthetized by inhalation of IsoVet (Schering-Plough Animal Health Corp.; Union, NJ), followed by cardiac perfusion with PBS (pH 7.4) and subsequently with 4% paraformaldehyde (Merck; Darmstadt, Germany) in PBS (pH 7.4). Dissected brains were subjected to additional fixation for 24 hr (at room temperature) in a large volume of freshly prepared fixative, cryoprotected for an additional 24 hr (10% sucrose in PBS), frozen in chilled isopentane, and stored at −80C.
Immunohistochemical Analysis
Coronal 35-μm frozen sections were collected into 50 mM Tris/HCl, 1.5% (w/v) NaCl, pH 7.4 (TBS). All steps of immunostaining of free-floating sections were performed at 4C. The only exception was the pretreatment with selected detergents (and glycerol) at 1% concentrations in TBS: CHAPS (Calbiochem; San Diego, CA); saponin, deoxycholic acid, Trappsol, Tween 20, SDS, glycerol, and Triton X-100 (Sigma; St Louis, MO); Brij 96V (Fluka; St Louis, MO); and Brij 98 (Aldrich; St Louis, MO). Pretreatment was performed either at 4C or ambient temperature, as indicated for each experiment. After detergent pretreatment, all sections were washed and transferred to 4C. Sections were blocked for 2 hr in TBS containing 1% bovine serum albumin (Sigma) plus 5% goat serum (Gibco; Carlsbad, CA), with or without selected detergents. Blocking solution with detergent was used also for diluting antibodies in one of the experiments (as indicated). In all other steps and experiments, blocking solution without detergents was used. All steps were performed on an orbital shaker.
Primary antibodies were incubated overnight in blocking solution. Anti-GM1 and anti-GD1a were produced as described previously (Schnaar et al. 2002); anti-Thy-1 antibodies were from BD Biosciences (San Diego, CA) and anti-MAG antibodies were from Calbiochem. Anti-GM1 antibody was used at 0.30 μg/ml and anti-GD1a at 0.06 μg/ml; anti-MAG was diluted at 1:500, and anti-Thy-1 was diluted 1:2000. Sections were then washed with TBS and incubated for 6 hr in 2 μg/ml biotin-conjugated goat anti-mouse IgG or alkaline phosphatase–conjugated goat anti-rat (Fc specific; Jackson ImmunoResearch, West Grove, PA). Sections were washed as above and (in the case of biotin-conjugated secondary antibody) incubated with Avidin DH-biotinylated alkaline phosphatase conjugate for 2 hr and developed with BCIP/NBT alkaline phosphatase substrate following the manufacturer's protocol (Vector Laboratories; Burlingame, CA). Control sections were done by omission of primary antibody. Sections were mounted and covered with VectaMount (Vector Laboratories).
Lipid extraction for the visualization of MAG was performed by dehydration in gradient of ethanol, chloroform/methanol extraction, and subsequent rehydration in gradient of ethanol prior to immunohistochemistry.
All tissue sections for one experiment were incubated for exactly the same time in all steps, and all images were collected using the same settings; thus, the differences in the intensity of staining reflect differences in the amount of bound antibody.
Results
In an effort to enable concurrent immunohistochemical analysis of myelin membrane receptors (like MAG) and gangliosides, we analyzed the effects of different detergents: CHAPS, saponin, deoxycholic acid, Trappsol, Tween 20, Triton X-100, Brij 96V, Brij 98, SDS, and glycerol. Published protocols reported use of these detergents in concentrations from 0.1% to 10% (Bo et al. 1995; Wang et al. 2002) in various steps of immunohistochemistry protocols (either only in pretreatment or blocking, during incubation with primary antibodies, or even in all steps). Recently, we reported that even lower concentrations of Triton X-100 cause significant redistribution of lipid-anchored antigens (Heffer-Lauc et al. 2005), but because 1% concentration was reported in most publications (including protocols for the isolation of lipid rafts), in this study, we used all detergents in 1% concentration.
Effects on the distribution of GD1a ganglioside of 2-hr pretreatment of brain sections with detergents at room temperature are shown in Figure 1. When immunostaining was performed without any detergents, GD1a was located primarily on neuronal bodies, with practically no staining of the white matter. All detergents, to some extent, significantly extracted GD1a from the gray matter. CHAPS, deoxycholic acid, Triton X-100, Brij 96V, and SDS were particularly detrimental and removed nearly all GD1a from the tissue.
Fluidity of membranes is much lower at lower temperatures, and as shown in Figure 2, we found that detergents were much less efficient in extracting GD1a when used at 4C. Previously, we reported that at this temperature, Triton X-100 causes redistribution of GD1a from gray matter into the white matter of cerebellum (Heffer-Lauc et al. 2005). We found that CHAPS and deoxycholic acid are even more efficient in this, whereas other detergents cause less or no redistribution.
Thy-1 is a GPI-anchored protein with a relatively large hydrophilic area that is tethered to the membrane by a relatively small lipid portion of the molecule. We have previously shown that Triton X-100 causes a redistribution of Thy-1 very similar to that of GD1a. We repeated the same experiments as shown in Figures 1 and 2 for GD1a with antibodies against Thy-1 and obtained similar results as for GD1a (presented in supplementary Figures 1 and 2).
Unlike GD1a and Thy-1, which are located mainly in the gray matter, GM1 is found predominately in the hydrophobic myelin sheets of the cerebellar white matter. In our previous study, we did not observe any redistribution of GM1 with Triton X-100. As shown in Figure 3, at 4C, none of the studied detergents caused redistribution of GM1 from white matter into the gray matter. However, some of the detergents (particularly SDS and deoxycholic acid) extracted significant amounts of GM1 from tissue sections, resulting in markedly reduced GM1 staining. This effect was even more prominent when the incubation with detergents was performed at room temperature (see supplementary Figure 3).
In the experiments presented above, tissue sections were treated with detergents for 2 hr before the blocking step and all other steps were performed at 4C (from blocking up to development with the substrate). However, in some published protocols, detergents are present in blocking and also during the incubation with primary antibodies. This not only prolongs the incubation in the presence of detergents, but also adds the effect of antibodies, which act as a large bulky handle that can pull lipid-anchored antigens out of the membrane. Aiming to evaluate the effects of this prolonged incubation with detergents in the presence of primary antibodies, we designed and performed the following experiment (presented in Figure 4).
Tissue sections of cerebellum and medulla from wild-type mice were coincubated for 16 hr with sections of spinal cord from B4 galnt1-null mice (lacking complex gangliosides, including GD1a) in 300 μl blocking solution containing antibodies against GD1a. When there was no detergent in the blocking solution, GD1a was found to be located primarily in the gray matter of cerebellum. There was no GD1a staining in B4 galnt1-null mice. However, when CHAPS, deoxycholic acid, or Triton X-100 was included in the blocking solution and solution of primary antibody, GD1a redistributed into the white matter of wild-type mouse cerebellum and also into the white matter of B4 galnt1-null mice spinal cord. Although it could be speculated that these detergents only revealed previously cryptic GD1a in the white matter of wild-type mouse cerebellum, the appearance of GD1a in B4 galnt1-null mice could only be explained by its redistribution from wild-type mouse cerebellum into B4 galnt1-null mice spinal cord during their coincubation in the presence of detergents. Interestingly, other detergents studied did not cause this type of redistribution.
Discussion
Immunohistochemical analysis is a common basic method that is a prerequisite for subsequent functional studies and in vivo imaging. However, antibodies are bulky molecules that need a significant amount of space to be able to bind to corresponding epitopes. Many molecules are either packed too close together or masked by other molecules, and the analysis of these structures is possible only when detergents are used to enable access of antibodies to the studied epitopes. Triton X-100 is one of the detergents most commonly used for the immunohistochemical analysis of membrane antigens and the isolation of membrane rafts (Edidin 2003). However, we recently reported that Triton X-100 can cause significant redistribution of gangliosides and GPI-anchored proteins (Heffer-Lauc et al. 2005) and that its use can compromise both the results of the immunostaining procedure and the composition of isolated lipid rafts. Although treatment with detergents is needed to detect some membrane antigens, such as MAG (see supplementary Figure 4), Triton X-100 might not always be the best choice, despite the fact that it is widely used and very efficient. Aiming to find a better detergent, we compared nine detergents at both room temperature and at 4C.
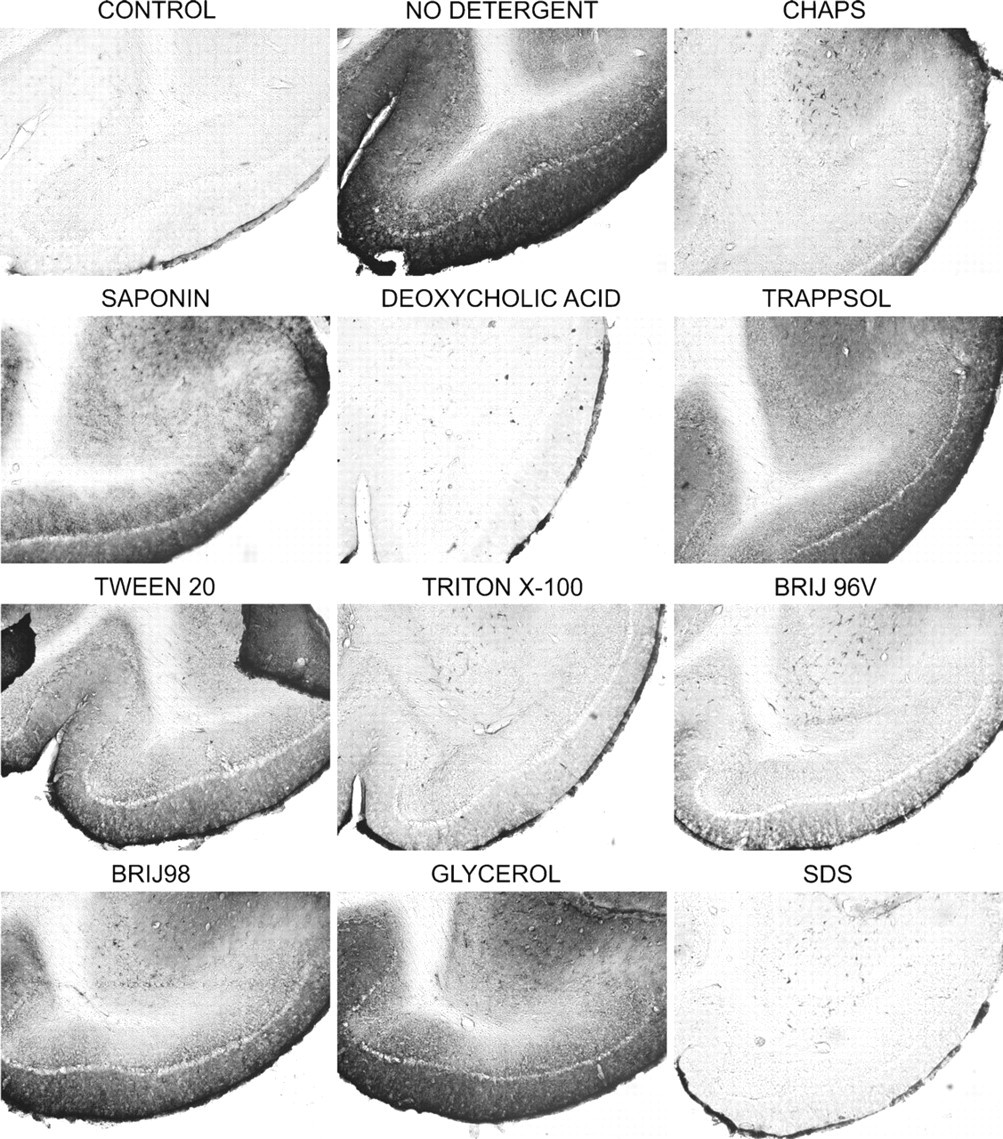
Effects of detergents on the immunohistochemistry of GD1a at room temperature. Wild-type mouse brain sections were preincubated with different detergents for 2 hr at room temperature and immunostained with anti-GD1a antibody. Significant extraction of GD1a was observed with CHAPS, deoxycholic acid, Triton X-100, and SDS, and to a lesser extent with Saponin, Trappsol, Tween 20, Brij 98, and glycerol.
At room temperature, nearly all detergents studied extracted a significant amount of lipid-anchored antigens from the tissues (Figure 1 and supplementary Figures 1 and 3). Depending on the type of detergent and the volume of buffer, extracted gangliosides and GPI-anchored proteins either remained in the solution or entered other tissues, primarily highly hydrophobic sheets of myelin. The only exception was GM1, which is physiologically located predominantly in myelin; detergents only extracted GM1 from the tissue, and it never entered any other tissue (Figure 3 and supplementary Figure 3). This observation is consistent with our previously reported results (Heffer-Lauc et al. 2005) and probably reflects differences in physicochemical properties between GM1 and other more-hydrophilic gangliosides. Even at 4C, some detergents, such as Triton X-100, CHAPS, and deoxycholic acid, caused significant extraction and redistribution of gangliosides and GPI-anchored proteins (Figure 2 and supplementary Figure 2). Other detergents, particularly Saponin, Trappsol, and Tween 20, were less deleterious. However, it should be noted that these are weaker detergents than Triton X-100, and that they might not be as efficient in revealing masked transmembrane epitopes. Also, different lipid-linked molecules behave differently in different detergents.
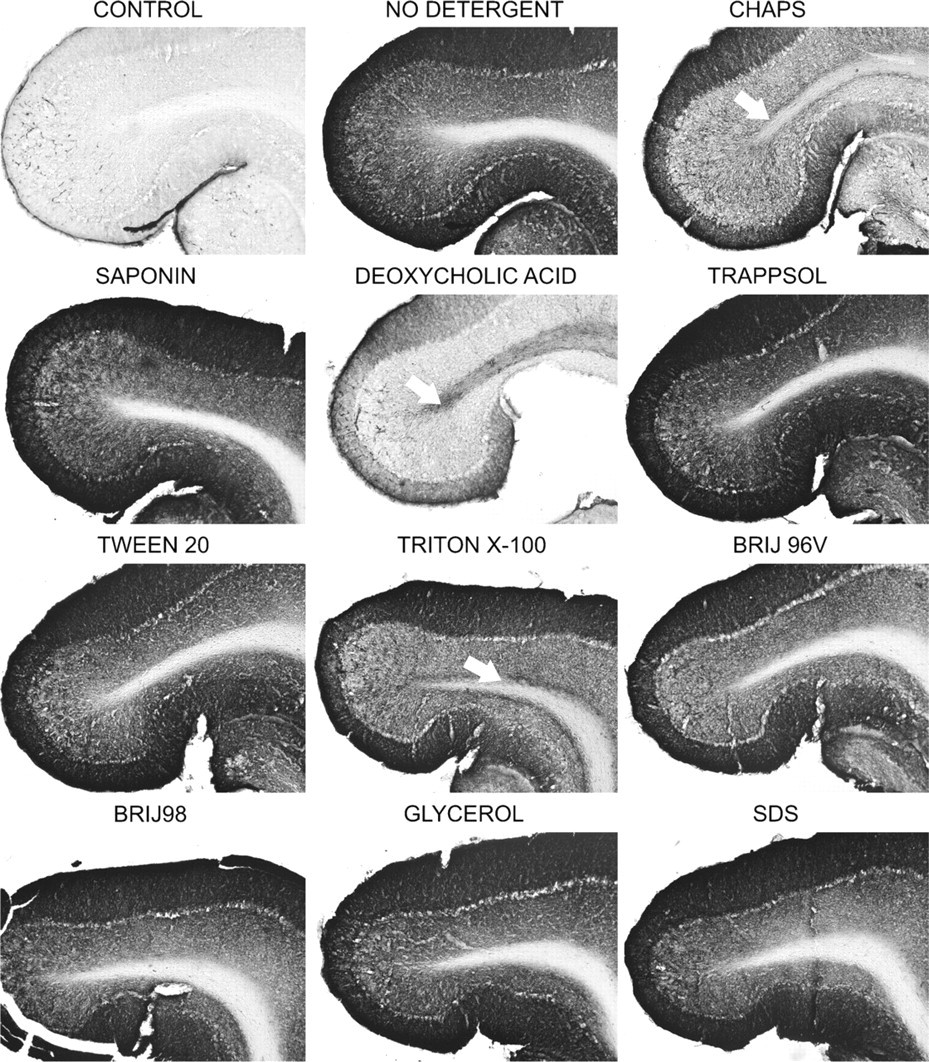
Effects of detergents on the immunohistochemistry of GD1a at 4C. Wild-type mouse brain sections were preincubated with different detergents for 2 hr at 4C and immunostained with anti-GD1a antibody. CHAPS, deoxycholic acid, and Triton X-100 caused significant redistribution of GD1a from gray matter into the white matter of cerebellum (marked with arrows), whereas other detergents (particularly Saponin, Trappsol, Tween 20, Brij 96V, and Brij 98) did not have this effect.
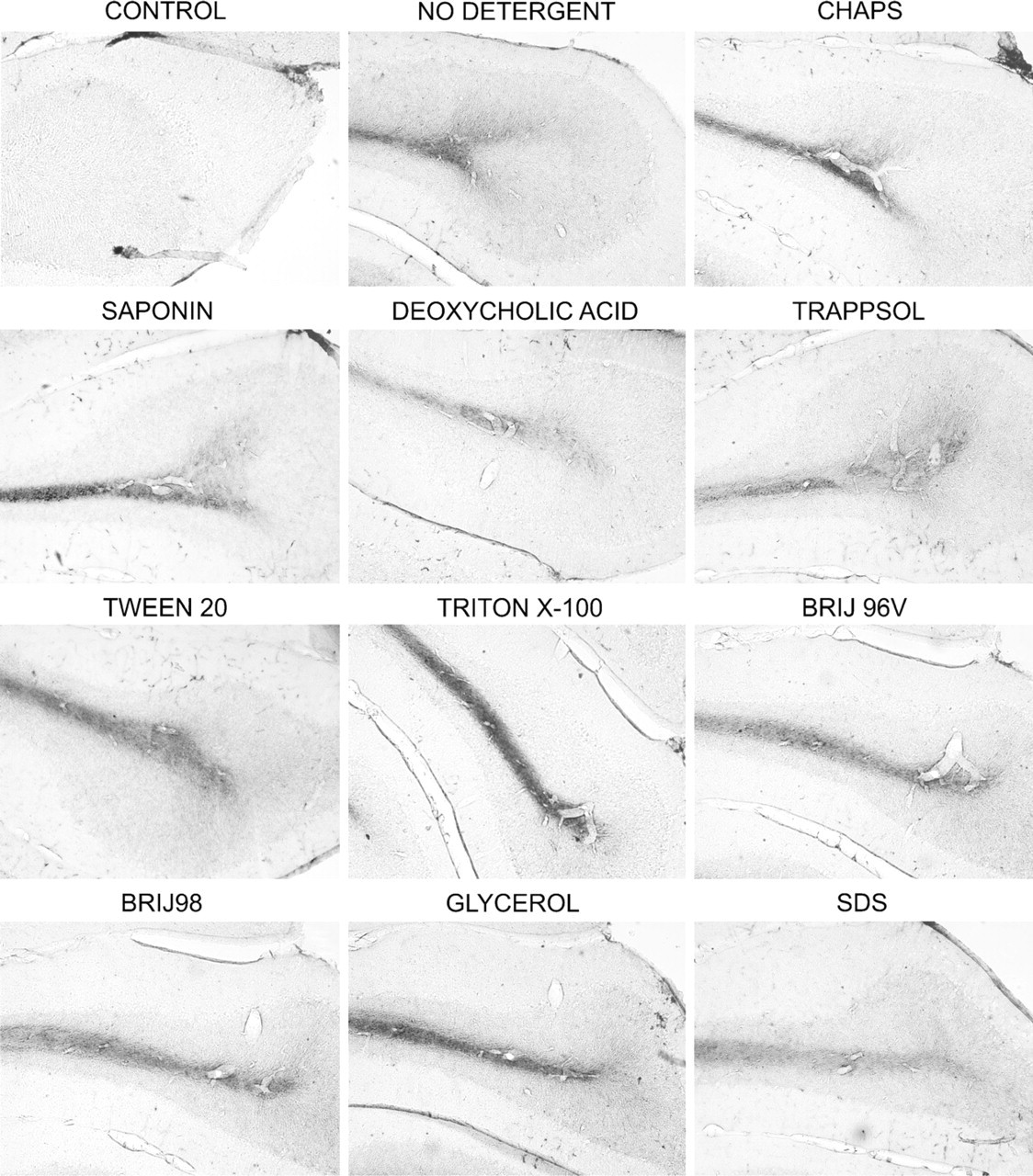
Effects of detergents on the immunohistochemistry of GM1 at 4C. Wild-type mouse brain sections were preincubated with different detergents for 2 hr at 4C and immunostained with anti-GM1 antibody. Some of the detergents (particularly SDS and deoxycholic acid) extracted significant amounts of GM1 from tissue sections, resulting in markedly reduced GM1 staining, whereas Triton X-100 and CHAPS enhanced staining.
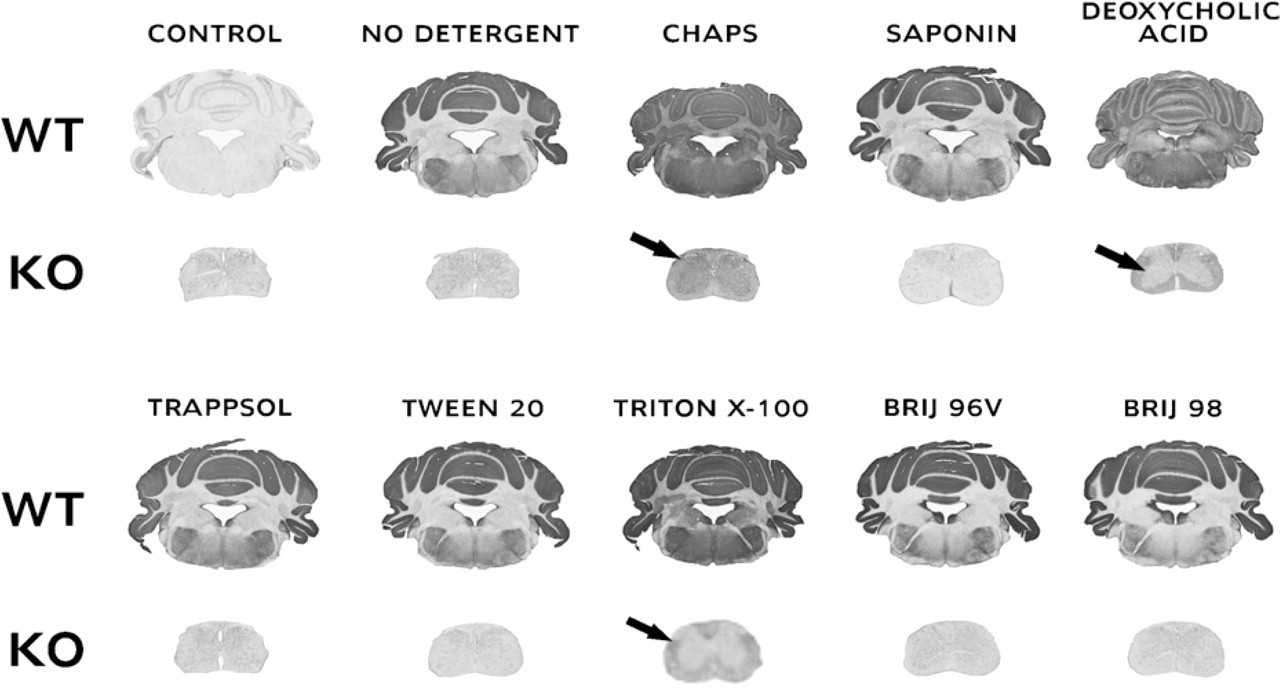
Effects of detergents on transfer of GD1a from wild-type mouse brain into spinal cord of B4 galnt1-null mice. Wild-type mouse brain sections (cerebellum and pons) were coincubated for 16 hr with slices of spinal cord from B4 galnt1-null mice in 300 μl blocking solution containing antibodies against GD1a and different detergents. CHAPS, deoxycholic acid, and Triton X-100 caused significant redistribution of GD1a into the white matter of wild-type mouse brain and also into the white matter of B4 galnt1-null mice spinal cord (marked with arrows). WT, wild type; KO, knockout.
In conclusion, the use of detergents has significant adverse affects on the immunochemical analysis of gangliosides and GPI-anchored proteins. Different detergents and detergent concentrations, and varying time and temperature, were found to affect lipid-anchored molecules, leading to immunohistochemical localization artifacts. Triton X-100 is used in most protocols for brain tissue staining and isolation of lipid rafts, but its use is associated with significant redistribution of lipid-anchored molecules, both within and between tissue sections. Although this might not be a problem in some studies, when gangliosides and GPI-anchored protein molecules are being examined, Triton X-100 should be avoided because it might significantly compromise obtained results. The same is true for CHAPS and deoxycholic acid. Use at room temperature of any of the detergents studied will cause significant extraction of lipid-anchored molecules, especially GPI-anchored proteins.
For concurrent immunohistochemical localization of lipid-anchored molecules and myelin proteins, the length of exposure to detergents should be reduced to a minimum, and all steps of the immunostaining procedure should be performed at 4C, because this significantly reduces detergent-induced artifacts. None of the nine detergents studied seems to be the perfect solution. Saponin, Trappsol, Tween 20, and Brij 98 caused less redistribution than other detergents, but their downside was that they were also not efficient in revealing cryptic epitopes like MAG. Brij 96V appears to be the least distorting choice of detergent; when used at 4C, it caused the least amount of redistribution but still sufficiently revealed myelin epitopes (Table 1). However, the behavior of lipid-linked epitopes varies, and other detergents might be optimal for other systems. In any case, it is very important to keep in mind that numerous control experiments are needed to avoid artifacts when lipid-linked molecules are studied in the presence of detergents.
Summary of the observed effects of different detergents
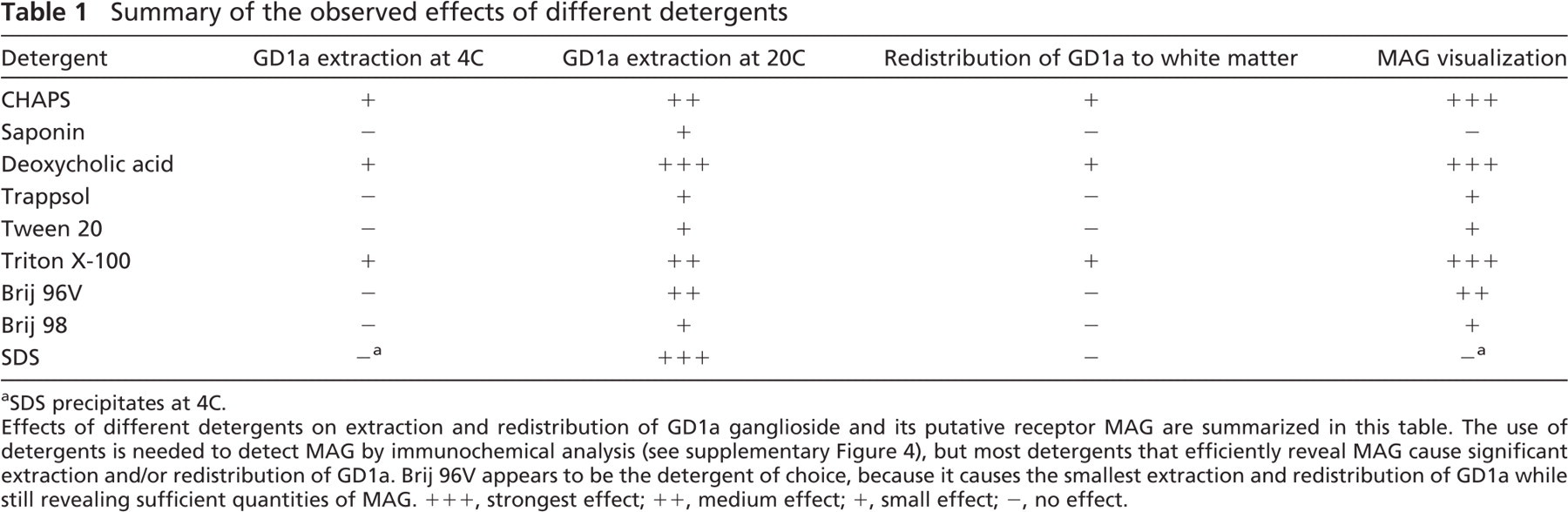
SDS precipitates at 4C.
Effects of different detergents on extraction and redistribution of GD1a ganglioside and its putative receptor MAG are summarized in this table. The use of detergents is needed to detect MAG by immunochemical analysis (see supplementary Figure 4), but most detergents that efficiently reveal MAG cause significant extraction and/or redistribution of GD1a. Brij 96V appears to be the detergent of choice, because it causes the smallest extraction and redistribution of GD1a while still revealing sufficient quantities of MAG. +++, strongest effect; ++, medium effect; +, small effect; −, no effect.
Footnotes
Acknowledgements
This work was supported by grants 219-0061194-2158 and 219-0061194-2023 from the Croatian Ministry of Science, Education, and Sport and by Grant 2R37NS037096-24 from the National Institutes of Health.