
Abstract
The MAL protein is the only integral membrane protein identified as being an essential component of the machinery necessary for apical transport in the canine MDCK cell line, a paradigm of polarized epithelial cells. To characterize the range of human epithelia that use MAL-mediated pathways of transport, we performed an immunohistochemical survey of normal tissues using a monoclonal antibody (MAb) specific for the MAL protein. For comparison, different types of carcinoma were also analyzed. MAL, with a characteristic strong supranuclear granular distribution, was detected in specific types of normal epithelial cells throughout the respiratory system, the gastrointestinal and genitourinary tracts, and in exocrine and endocrine glands. Absorptive cells (e.g., enterocytes), and many different types of specialized secretory cells, either organized in discrete clusters (e.g., endocrine cells in the pancreas), gathered together in an endocrine gland (e.g., thyroid), interspersed with other cells in glands (e.g., parietal cells), or dispersed singly among other cells (e.g., type 2 pneumocytes) were positive for MAL. We also analyzed a series of epithelial renal and thyroid tumors and found alterations dependent on the particular histological type of tumor. These results open potential applications of the anti-MAL antibody for the characterization of neoplastic tissue.
U
Although the specialized lipid composition of GEM rafts provides the biophysical basis for the specificity of protein recruitment, rafts require protein machinery to be operative in signaling or transport (Simons and Wandinger-Ness 1990). The MAL cDNA was originally identified during a search for genes differentially expressed during T-cell ontogeny (Alonso and Weissman 1987). MAL is an integral membrane proteolipid protein of 17 kD that selectively resides in lipid rafts in polarized epithelial Madin-Darby canine kidney (MDCK) and Fischer rat thyroid (FRT) cells, T-lymphocytes, and oligodendrocytes (Kim et al. 1995; Zacchetti et al. 1995; Martín-Belmonte et al. 1998; Millán and Alonso 1998). MAL is an itinerant protein that continuously shuttles between the trans-Golgi network, the plasma membrane, and endosomes (Puertollano and Alonso 1999). The elimination of the endogenous MAL protein in MDCK and FRT cells blocks apical transport of both membrane and secretory proteins (Cheong et al. 1999; Puertollano et al. 1999; Martín-Belmonte et al. 2000,2001). This demostrates an essential role for MAL, probably related to the process of formation of the transport carriers by polymerization of the vesicular coats (Puertollano et al. 1997,2001), as an element of the machinery for apical transport in these two cell lines.
The use of MDCK and FRT cells has been of invaluable help in establishing the role of MAL in apical transport (Cheong et al. 1999; Puertollano et al. 1999; Martín-Belmonte et al. 2000,2001). However, given the different morphology of epithelial cells, their organization in single or multiple layers, and their multiple tissue-specific functional specializations, it is not obvious which human epithelia use MAL for apical membrane trafficking. Using an already characterized monoclonal antibody (MAb) specific to MAL (Martín-Belmonte et al. 1998; Millán and Alonso 1998), in this study we have carried out an immunohistochemical analysis of the distribution of MAL in a wide variety of normal human tissues, with special emphasis on different types of epithelia and in specific types of tumors. In addition to being detected in the kidney and thyroid, as could have been anticipated from the results obtained in MDCK and FRT cells, the expression of MAL observed in the apical face in most of the epithelia examined in this study indicates that the MAL-mediated pathway of apical protein transport is employed by many different types of epithelia throughout the human body. It is of special note that alterations in the expression and/or subcellular distribution of MAL have been observed in specific types of carcinomas. The central role of MAL in membrane trafficking (Cheong et al. 1999; Puertollano et al. 1999; Martín-Belmonte et al. 2000,2001) and the information provided in this study extend to a wider range of tissues and cell types the use of MAL in raft-mediated routes of protein transport.
Materials and Methods
Immunohistochemical Analysis of Human Samples
Normal adult tissue samples were received in the Pathology Department of the Hospital de la Princesa (Madrid, Spain) as routine specimens obtained from surgery. In addition, specimens from epithelium-derived renal and thyroid neoplasms were studied. These tumors included the main varieties of renal cell carcinomas [clear-cell renal carcinoma (n = 8), papillary renal cell carcinoma (n=5), chromophobe cell type carcinoma (n = 5), sarcomatoid renal cell carcinoma (n=2), and oncocytoma (n = 4)] and the main types of follicle-derived thyroid carcinomas [(papillary carcinoma (n = 6), follicular carcinoma (n=4), and anaplastic carcinoma (n = 1)]. These cases were defined according to morphology assessed using hematoxylin-stained sections together with immuno-staining in selected cases (Rosai et al. 1992; Murphy et al. 1994).
All samples were fixed for several hours in 10% neutral buffered formalin and subjected to routine tissue processing and paraffin embedding. Sections of 5-μm thickness were prepared from paraffin-embedded tissues and mounted on poly-L-lysine-coated glass microslides. Antigen retrieval was accomplished by subjecting deparaffinized sections to pressure-cooker unmasking for 60 sec in 200 mM citrate buffer, pH 6.0. The tissue was then blocked with a 1:20 dilution of normal rabbit serum in 10 mM Tris-HCl saline buffer, pH 7.6, as previously described (Marazuela et al. 1995). The sections were sequentially incubated with a 1:50 dilution of a mouse ascites stock of anti-MAL 6D9 MAb and peroxidase-conjugated rabbit anti-mouse IgG (Dako; Glostrup, Denmark). Each incubation was followed by three washes with Tris-buffered saline. Then sections were developed with Graham-Karnovsky medium containing 0.5 mg/ml of 3,3'-diaminobenzidine tetrahydrochloride (Sigma Chemical; St Louis, MO) and hydrogen peroxide. Sections were counter-stained with Carazzi's hematoxylin, dehydrated, and mounted by routine methods. At least four coded samples from each tissue were examined by two independent observers.
Results
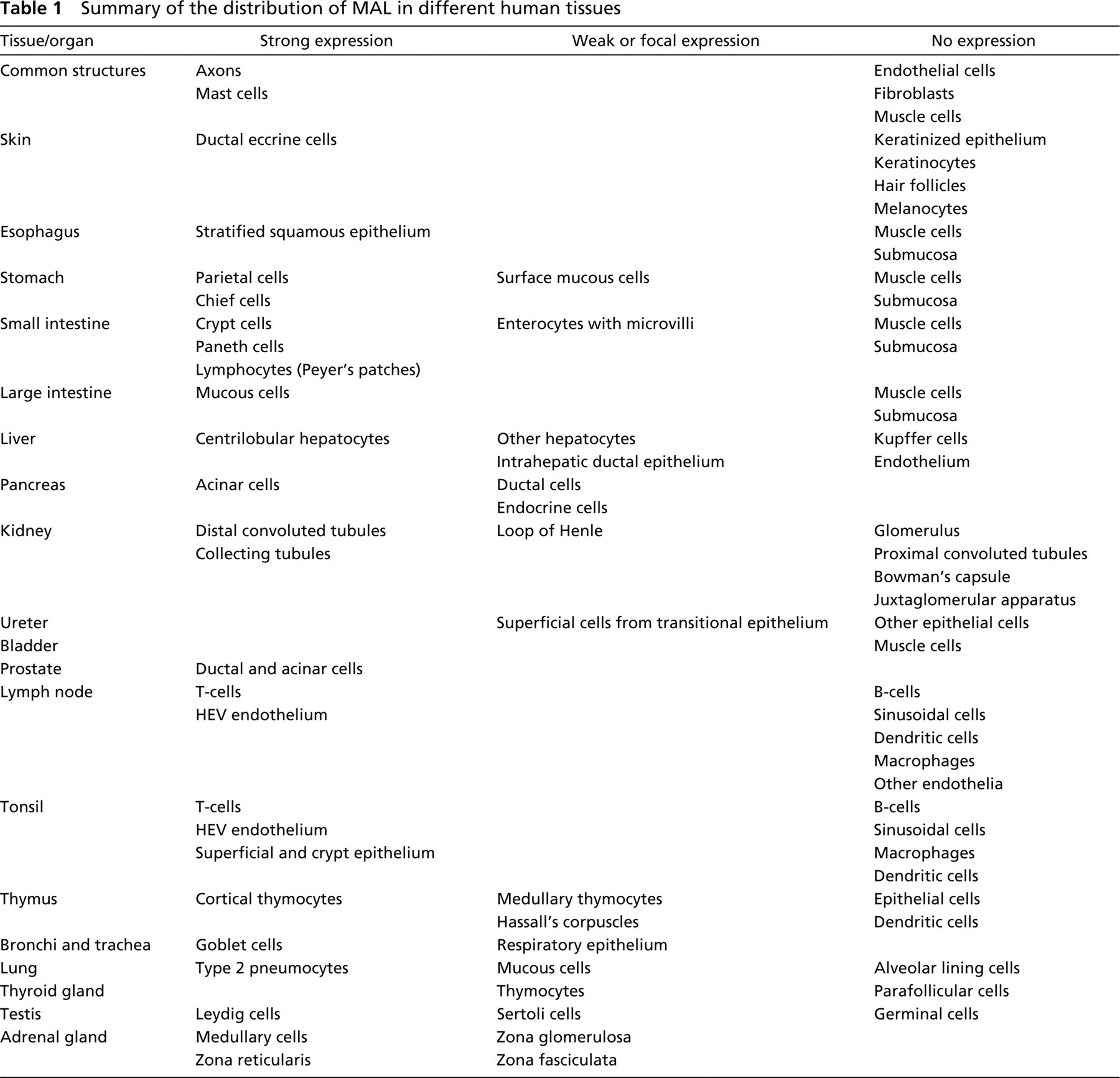
The generation, characterization, and specificity of the anti-MAL antibody (MAb 6D9) used in this study have been reported previously (Martín-Belmonte et al. 1998; Millán and Alonso 1998). Because the use of paraffin-embedded or unfixed frozen tissues gave similar patterns (our unpublished results), only the results with the former are shown. Common features observed during our immunohistochemical analysis of human tissue specimens with the anti-MAL MAb 6D9 were the expression of MAL in specific epithelia as well as in exocrine and endocrine glands, and the absence of detectable MAL expression in fibroblasts, endothelial cells, and skeletal and smooth muscle (compiled in Table 1). In addition, mast cells and the axons of peripheral nerves also showed strong immunostaining with the anti-MAL MAb (Figure 1A).
Sections of skin, including keratinizing squamous epithelium, dermis with adnexa, and subcutaneous fibroadipose tissue, were examined. All layers of keratinized squamous epithelium and subcutaneous fibroadipose tissue were negative (our unpublished results). Reactivity was also found in the apical portion of ductal eccrine cells of the sweat glands in both the apical and basal layers (Figure 1B), as well as in the acrosyringium (our unpublished results).
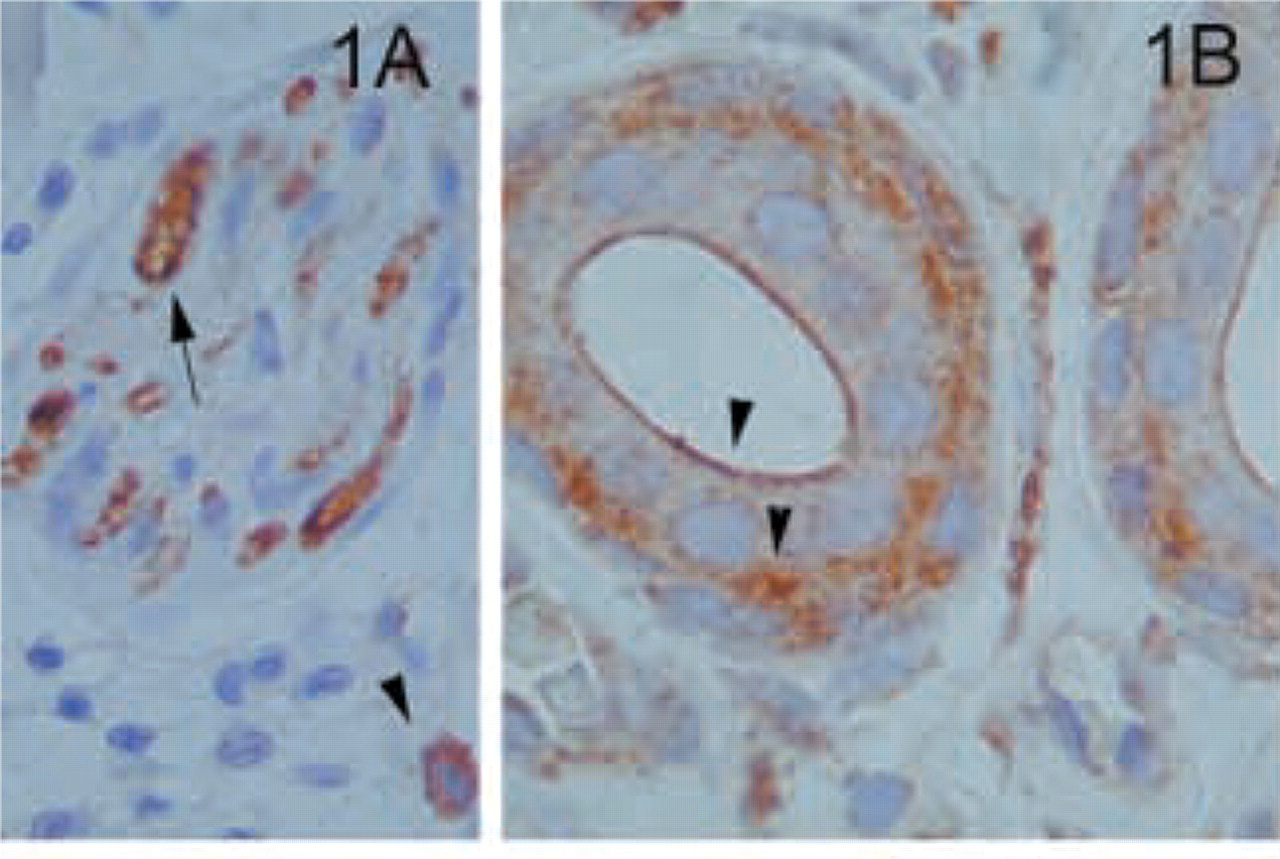
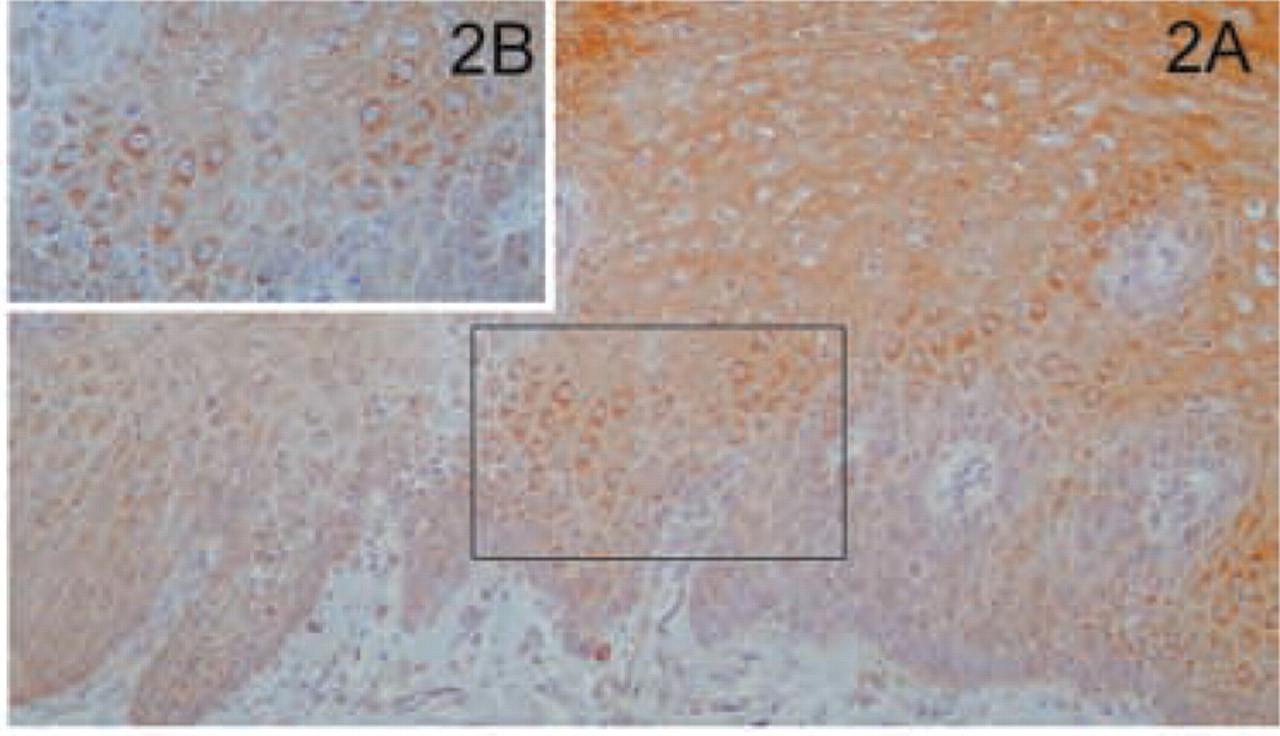
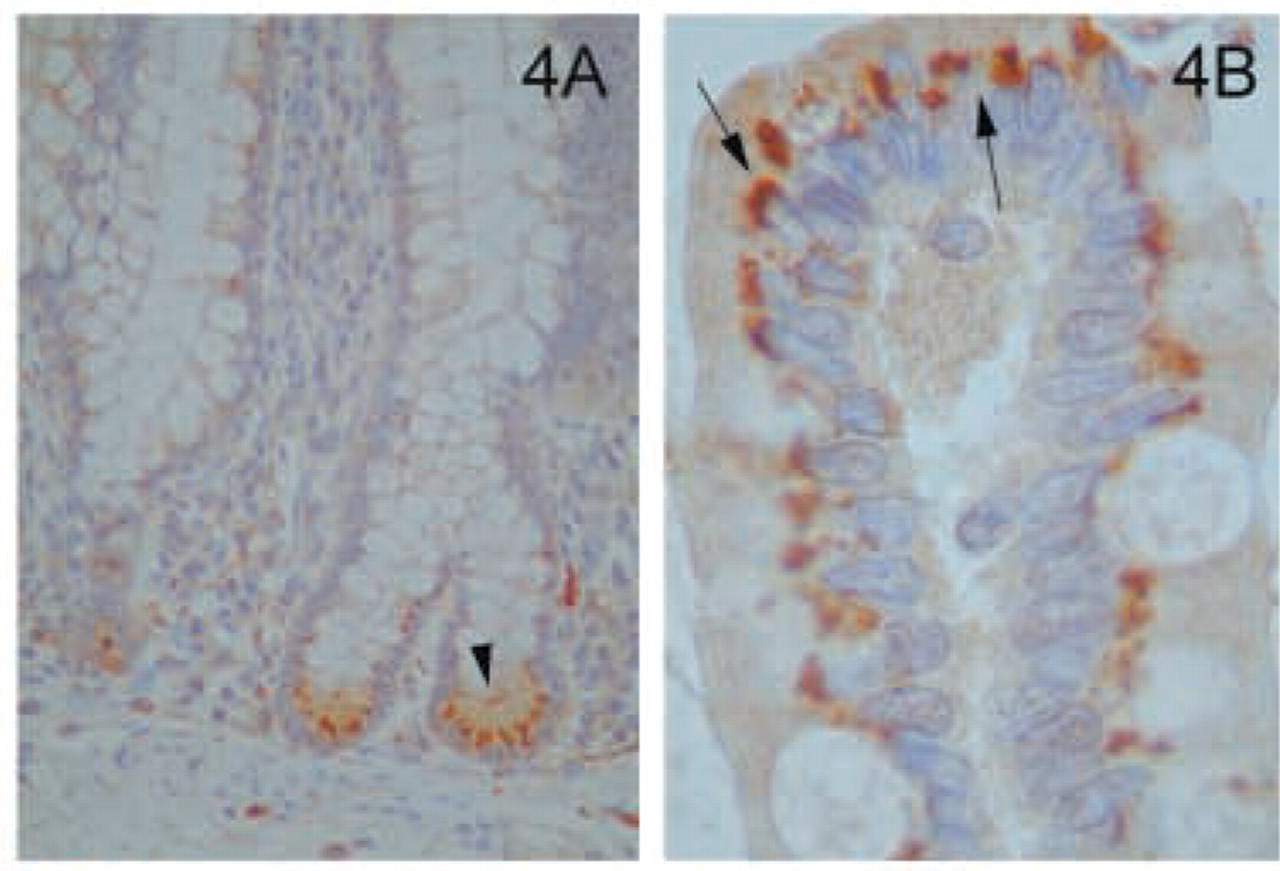
Evaluation of the gastrointestinal tract included examination of esophagus, stomach, ileum, colon, liver, and pancreas. The gastrointestinal epithelium was positive for MAL at all sites examined. In the esophagus, the stratified squamous, non-keratinizing epithelium was strongly positive (Figure 2A). The staining was more pronounced in the apical layer (Figure 2B). When the stratified epithelium changed to the gastric, mucus-secreting simple columnar epithelium of the stomach there was positive staining confined to the apical portion of the surface mucous cell (Figure 3A). In addition, parietal cells, responsible for the secretion of hydrochloric acid and intrinsic factor, were highly immunoreactive, typically in association with punctate cytosolic structures (Figure 3B). In the small intestine, the epithelial cells lining the crypts of Lieberkühn had a very distinctive reaction pattern. Cells at the base of the crypts, including Paneth cells, were strongly positive, with a gradual decrease in staining intensity as the cells approached the surface (Figure 4A). The staining of the cells of the villi showed a characteristic pattern of reactivity similar to that observed for other epithelial cells, i.e., strong supranuclear granular positivity, with no staining at the basal or lateral membranes (Figure 4B). This distribution corresponds to the location of the Golgi apparatus. In the Peyer's patches, only lymphocytes in the paracortical zone were stained (our unpublished results). Throughout the gastrointestinal tract, myenteric and submucosal plexus were strongly positive (our unpublished results). In contrast, the smooth muscle of the muscularis mucosa and the striated muscle of the muscle coat did not stain with the anti-MAL antibody. The mucosal epithelium of the large intestine expressed MAL in a distribution similar to that of the small intestine (our unpublished results).
Summary of the distribution of MAL in different human tissues
Skin and subcutaneous tissues. (
Esophagus. (
Stomach. (
Intestine. (
Liver. (
Pancreas. (
Kidney. (
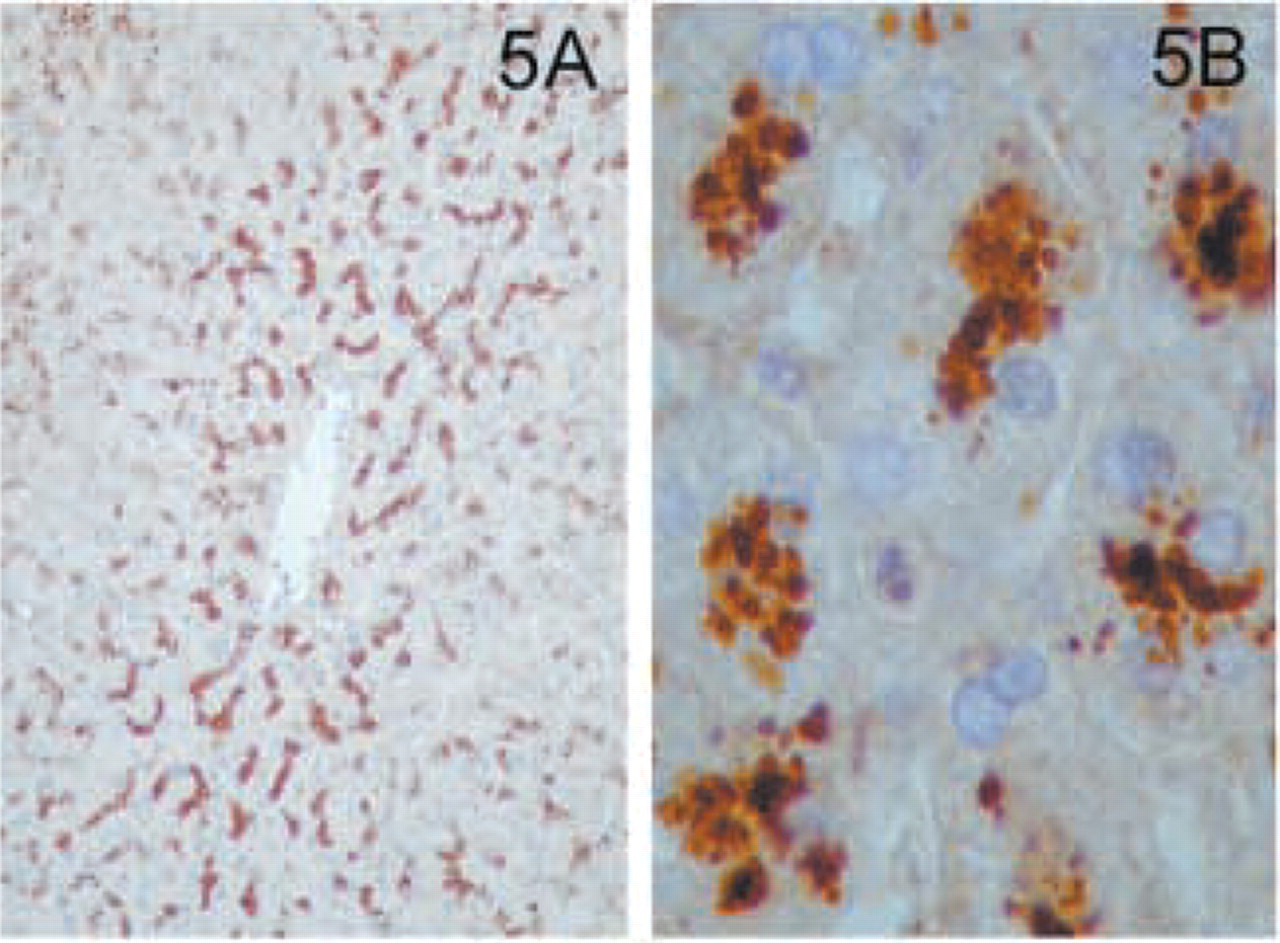
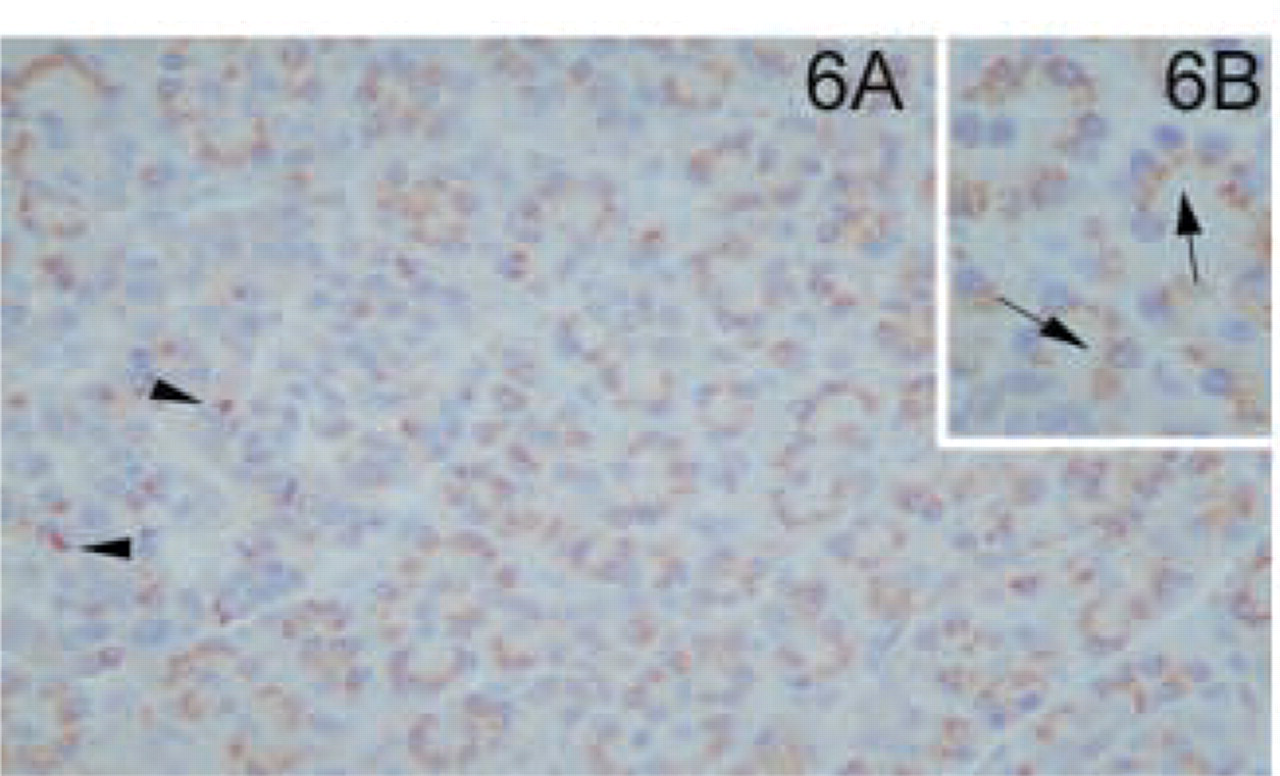
In normal liver, hepatocytes showed granular cytoplasmatic staining for MAL mostly in the perivenular or centrilobular areas (Figures 5A and 5B). Sinusoidal lining cells and Kupffer cells were negative. Bile duct epithelium was mildly positive (our unpublished results). In the pancreas, granular positivity was found in the supranuclear area of acinic cells (Figures 6A and 6B). In the islets of Langerhans, only sporadic cells were stained (Figure 6A).
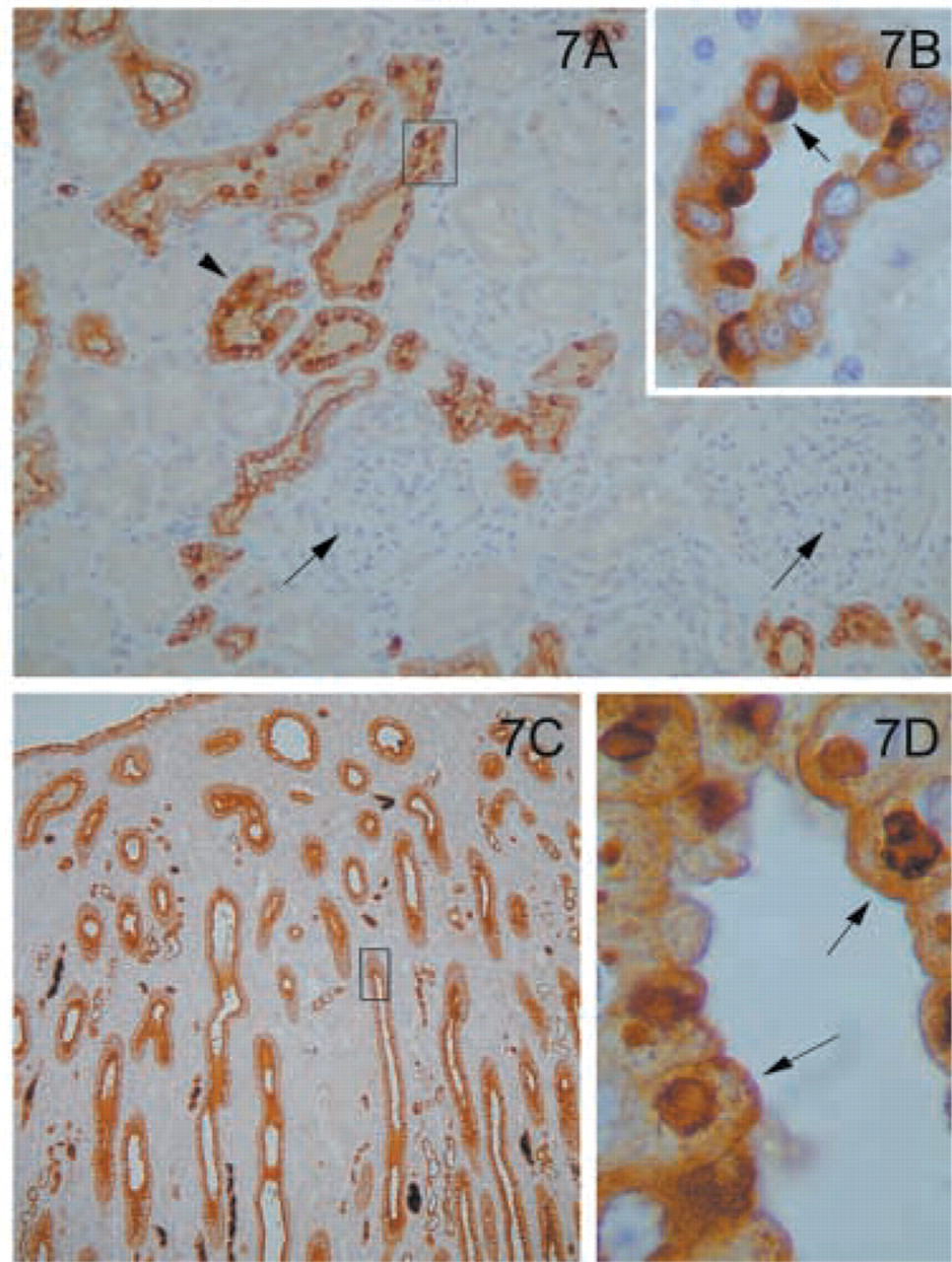
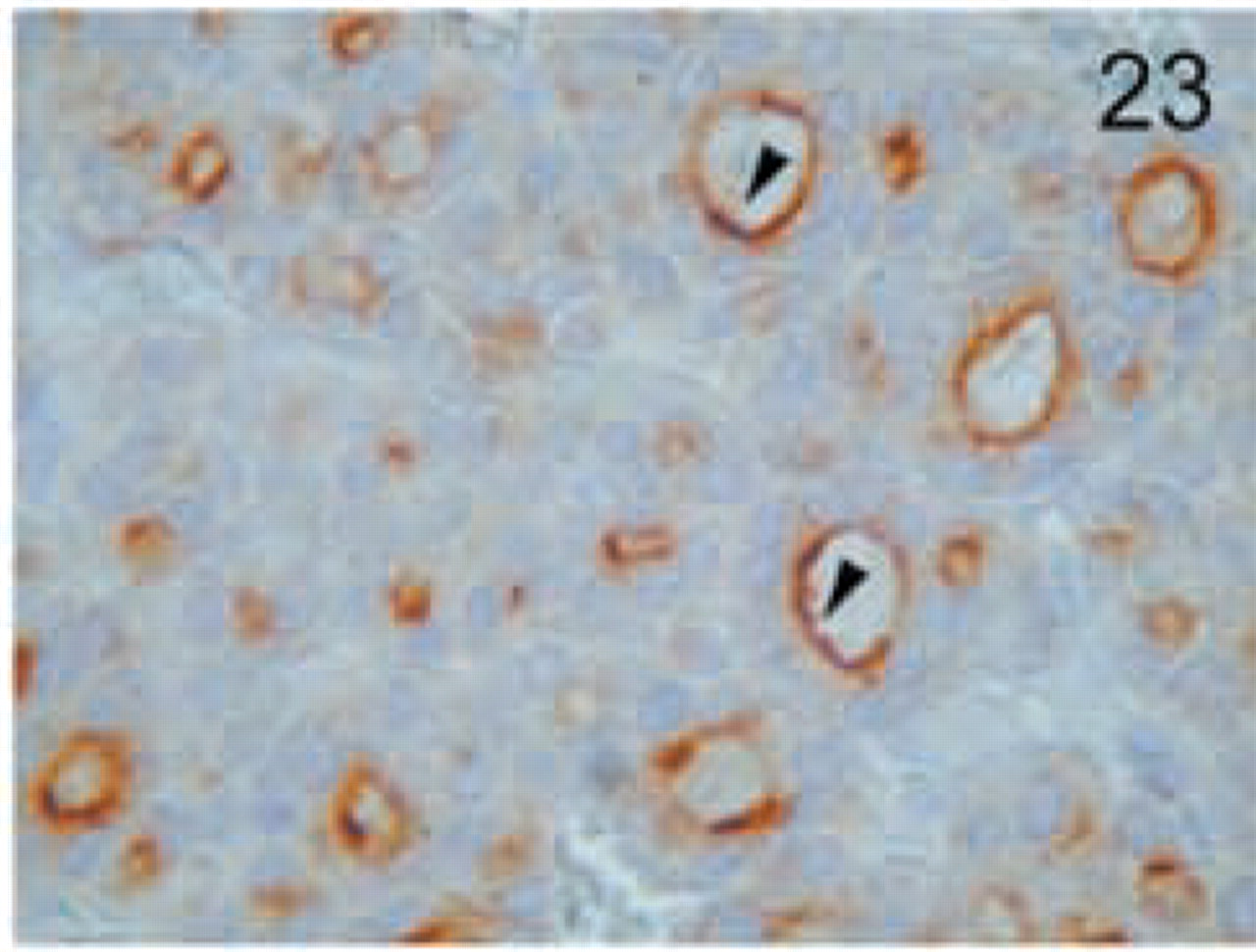
Multiple sites of the genitourinary tract expressed the MAL molecule. In the kidney, MAL was widespread in specific parts of the kidney tubuli. In the kidney cortex, MAL was present in the distal convoluted tubules but not in the proximal tubules (Figure 7A). Staining was more pronounced in some cells of the tubules than in others (Figure 7B). The glomeruli were negative (Figure 7A). Intense labeling was observed in the collecting tubules of the medulla (Figure 7C). The staining also had a characteristic granular apical pattern (Figure 7D). The transitional epithelium of the urothelium was also positive with characteristic granular staining in the superficial layer (Figure 8).
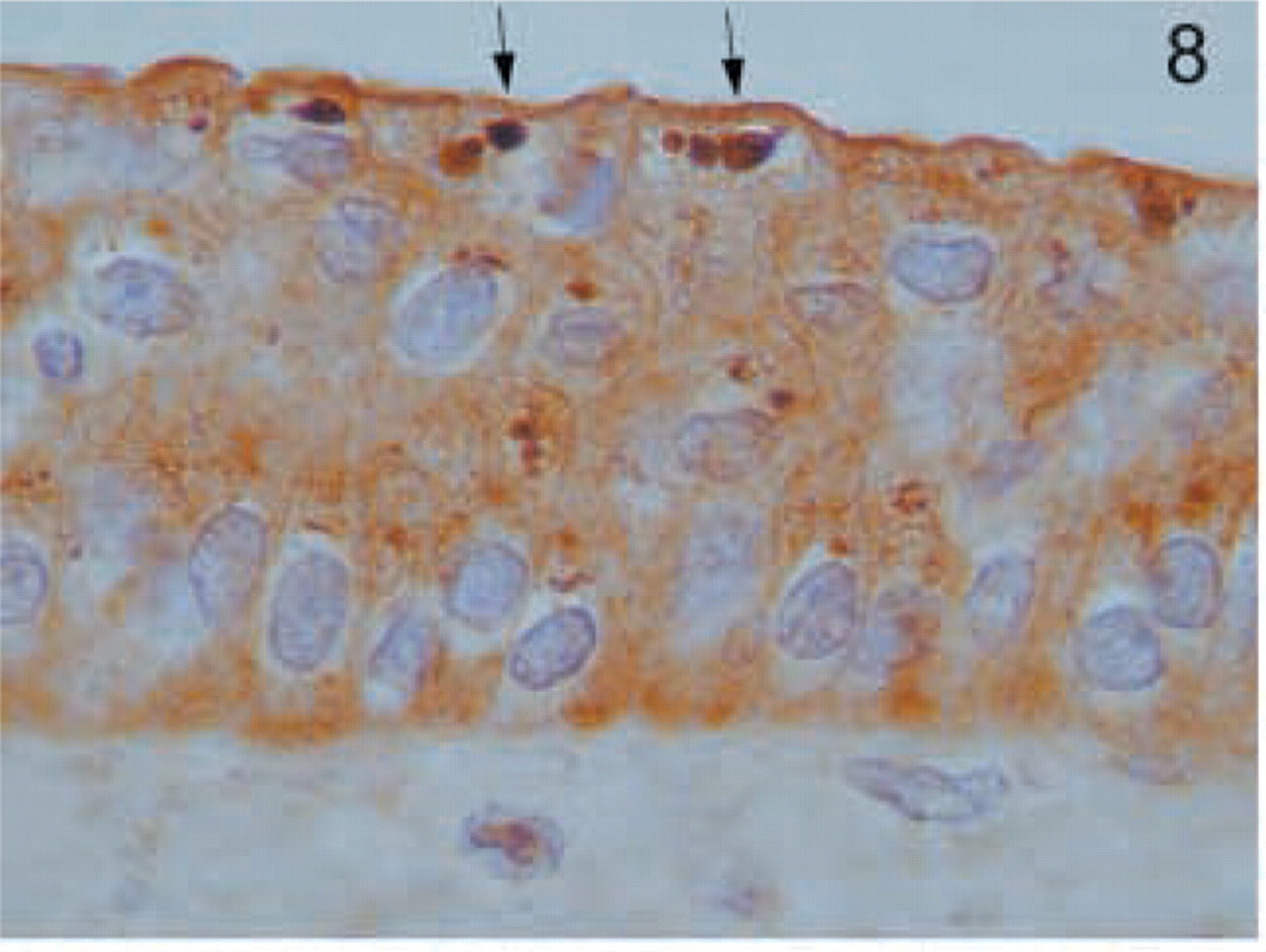
Urothelium. Staining in the transitional epithelium of the urothelium has a characteristic apical distribution, more marked in the superficial layer (arrows). Original magnification X100.
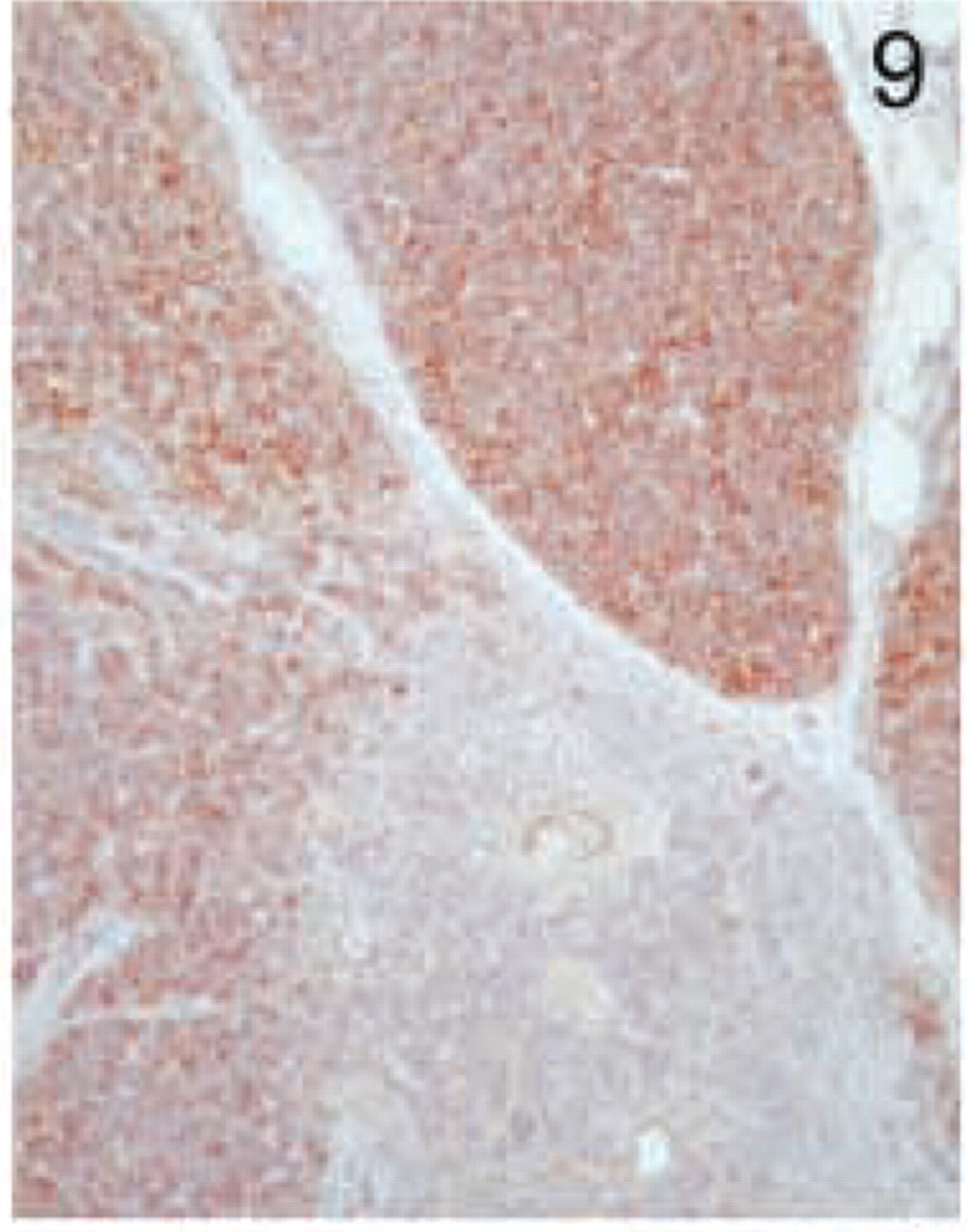
Thymus. Staining is restricted to the thymic cortex, with very few positive cells in the medulla. Original magnification X20.
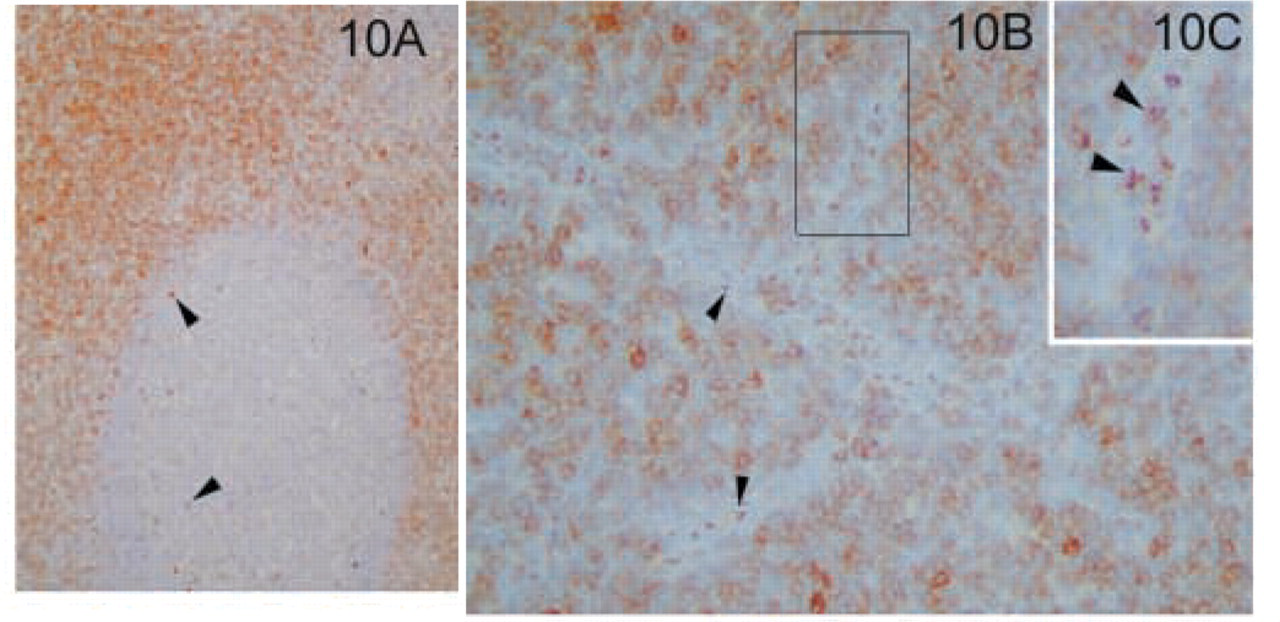
Lymph node. (
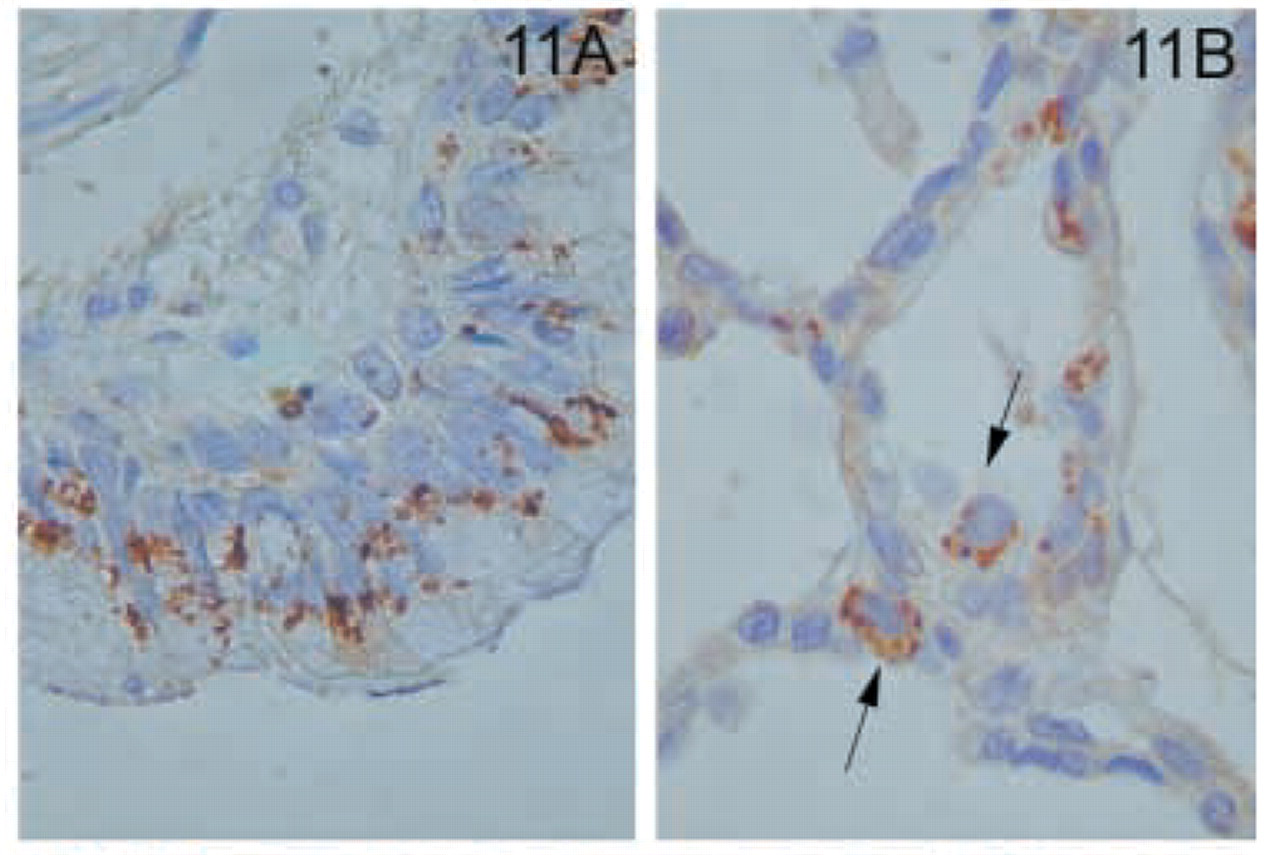
Lung. (
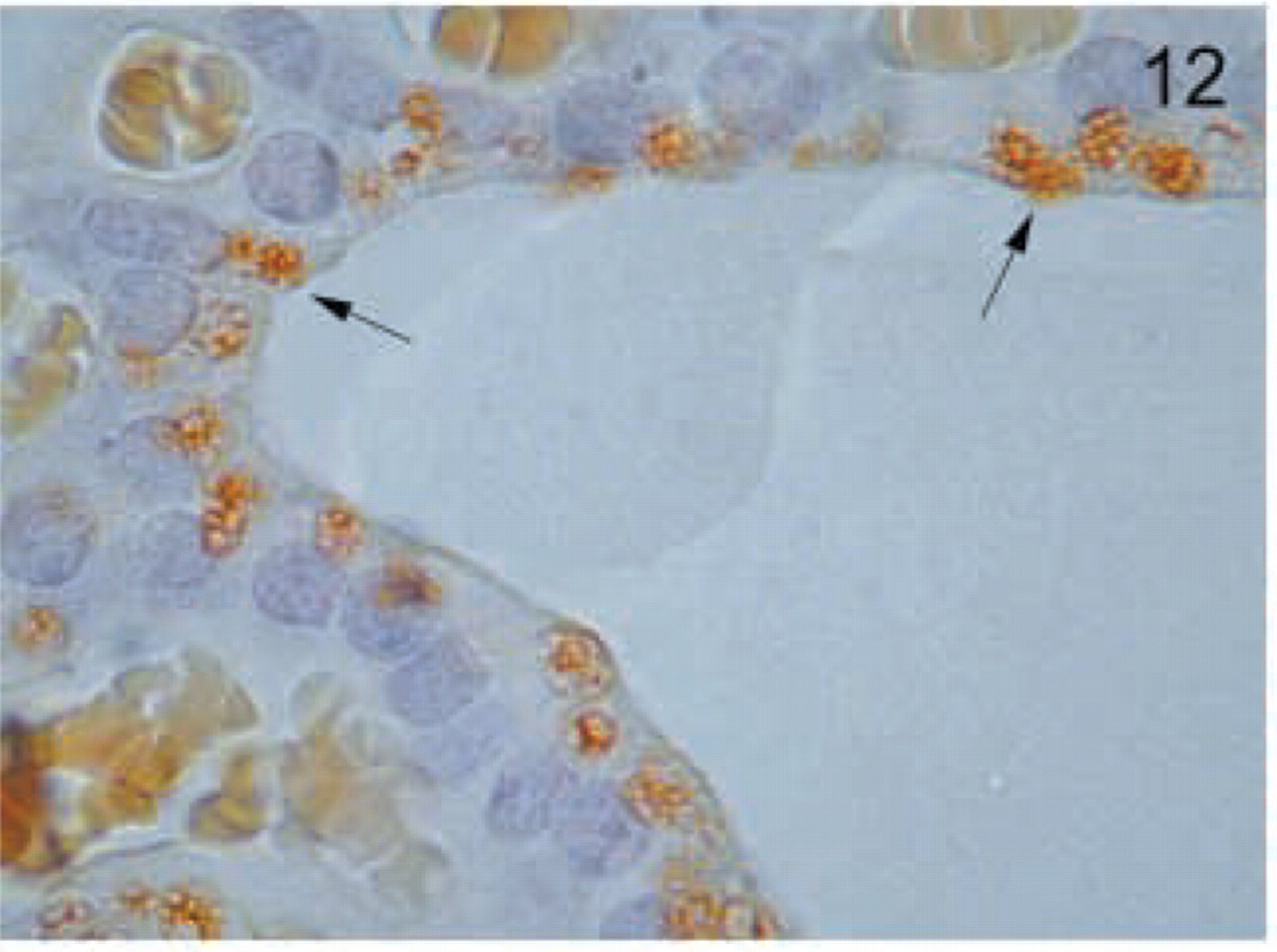
Thyroid. Thyroid follicular cells have a characteristic granular apical distribution of MAL (arrows). Original magnification X100.
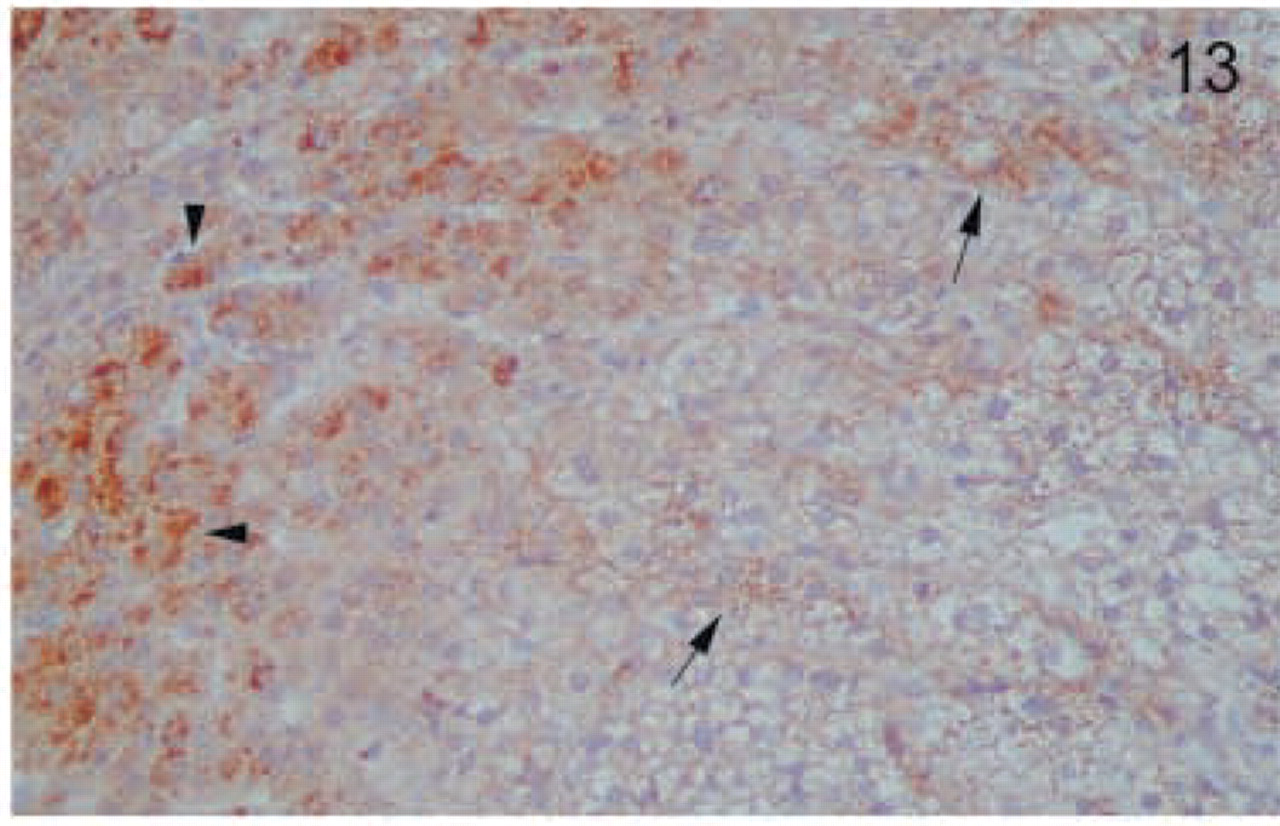
Adrenal gland. Secretory cells of the adrenal medulla show intense granular staining (arrowheads). All layers of the adrenal cortex stained positively, most strongly in the zona reticularis (arrows). Original magnification X40.
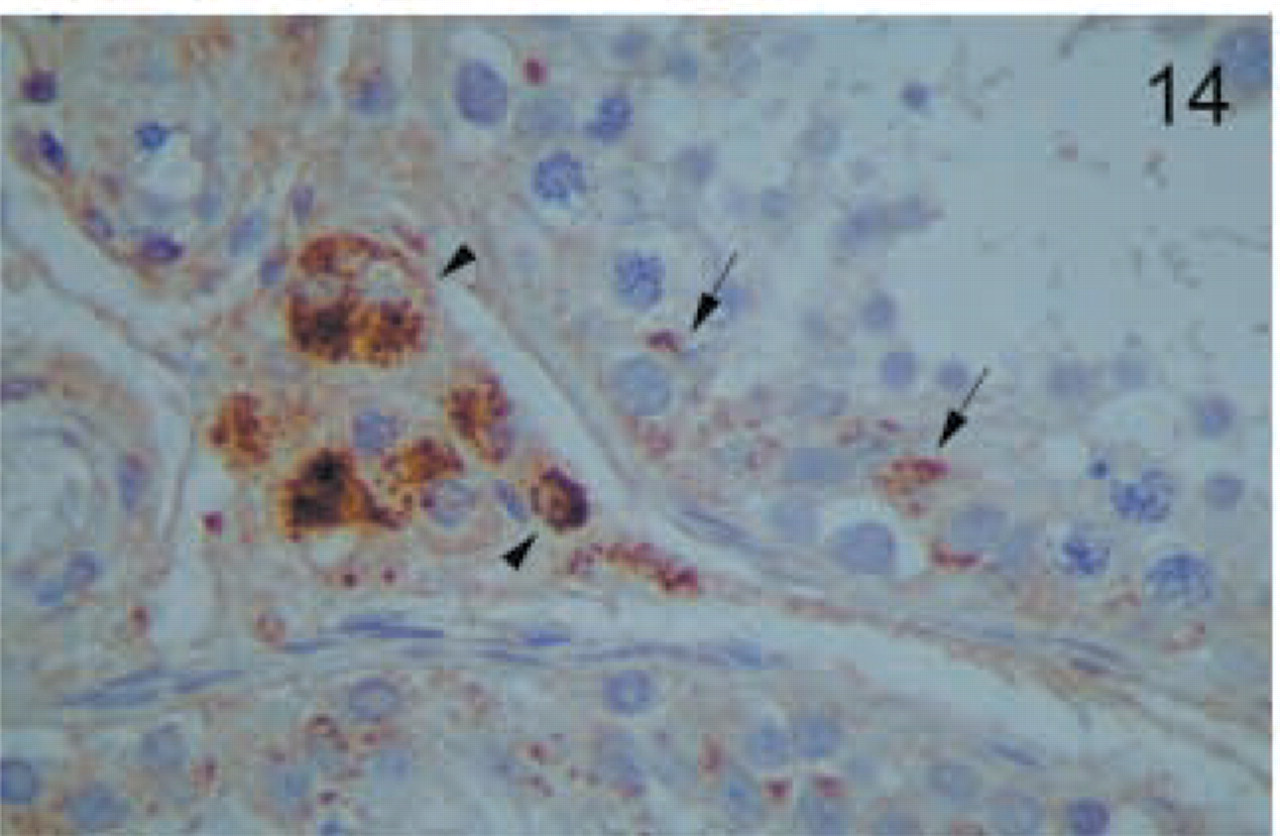
Testis. Strong granular positivity is found in secretory Leydig cells (arrowheads), and faint staining is detected in Sertoli cells (arrows). Germinal cells were negative. Original magnification X100.
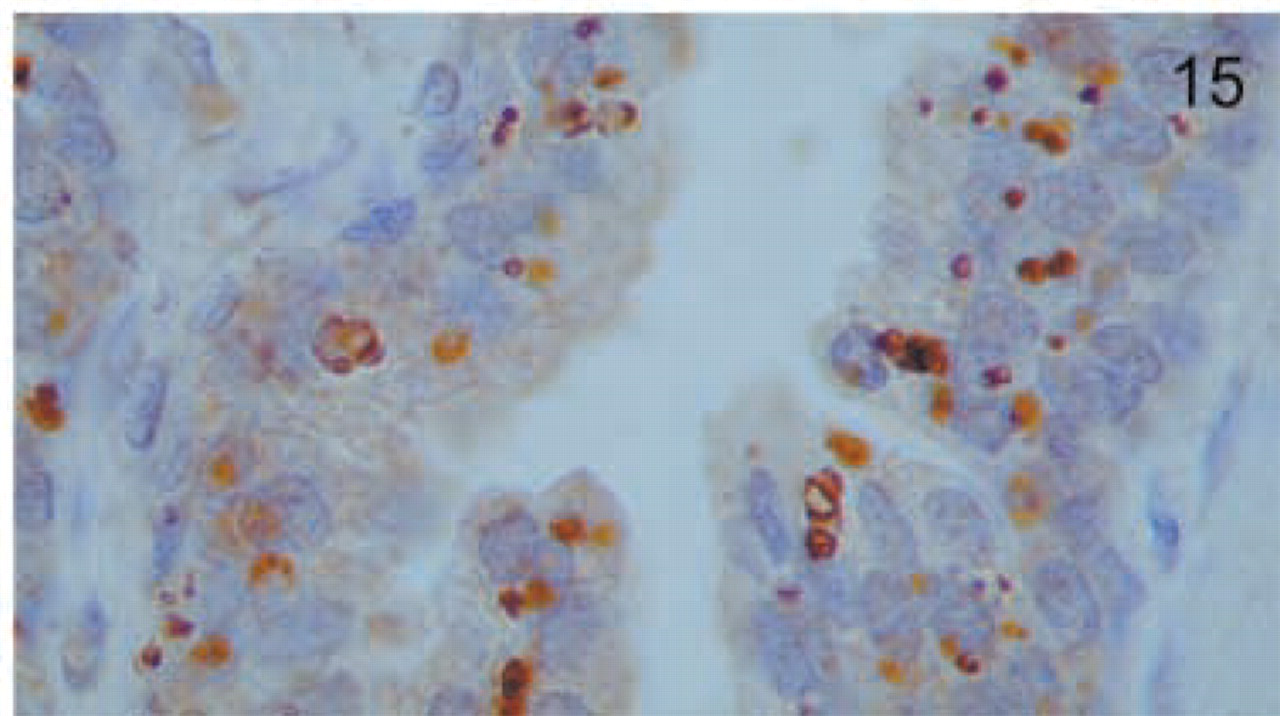
Prostate. The epithelium of the prostate glands shows intense apical staining for MAL. Original magnification X100.
The hematopoietic tissues analyzed included thymus, lymph node, and tonsil. In the thymus, the expression of MAL was restricted to the thymic cortex in the T-cell areas, with very few positive cells in the medulla (Figure 9). In normal lymph node and tonsil there were similar patterns of staining. Staining was confined to paracortical lymphocytes (T-cell zone) and to sporadic T-lymphocytes within the follicles (Figure 10A). In contrast to the lack of expression of the MAL antigen in endothelial cells, high endothelial venules were strongly positive, with a characteristic apical distribution (Figures 10B and 10C). Dendritic cells and follicular dendritic cells were negative (our unpublished results).
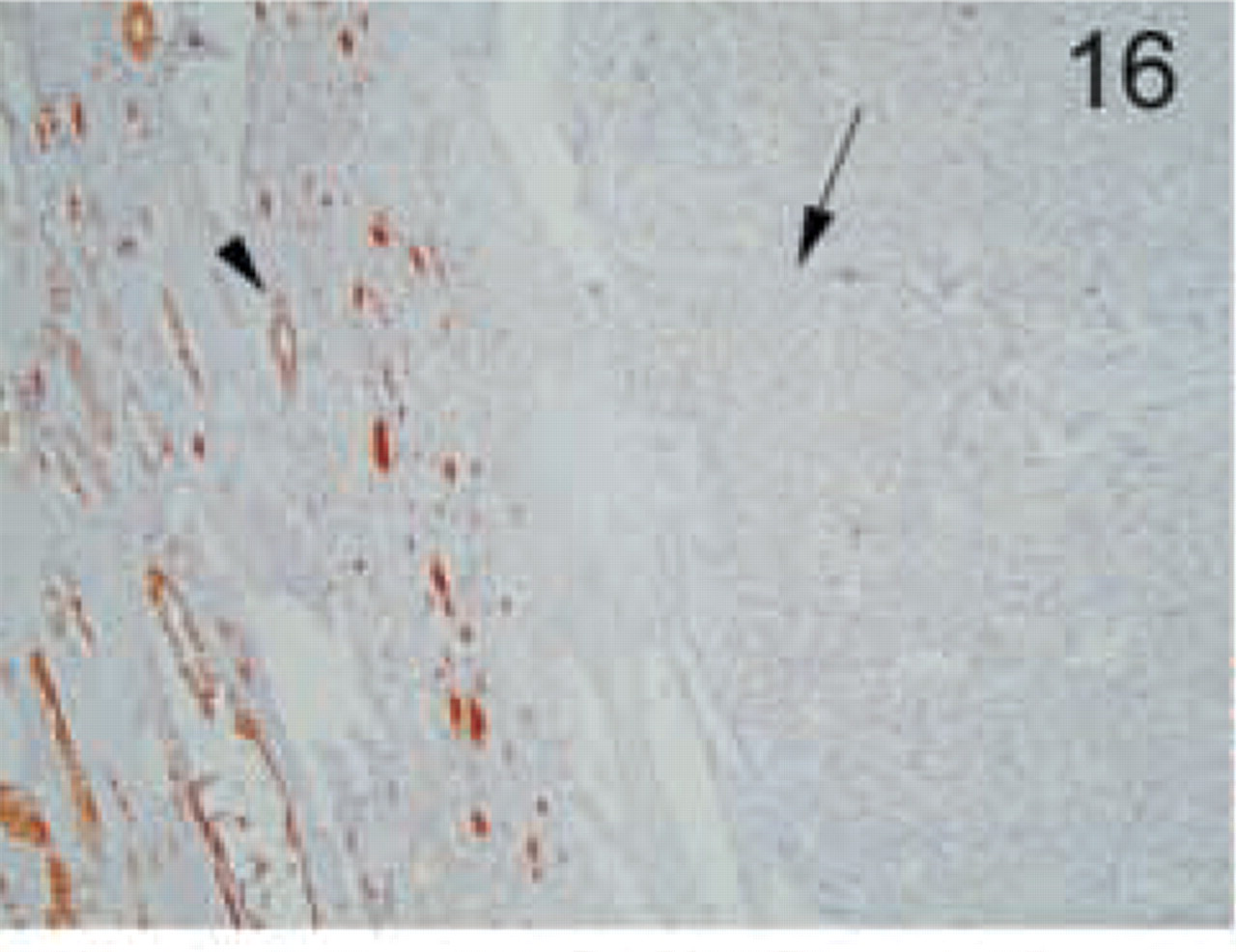
Staining of epithelial renal tumors: clear-cell carcinoma. No staining was found in renal cell carcinoma cells (arrow). Normal staining is found in the adjacent normal kidney cells (arrowhead). Original magnification X10.
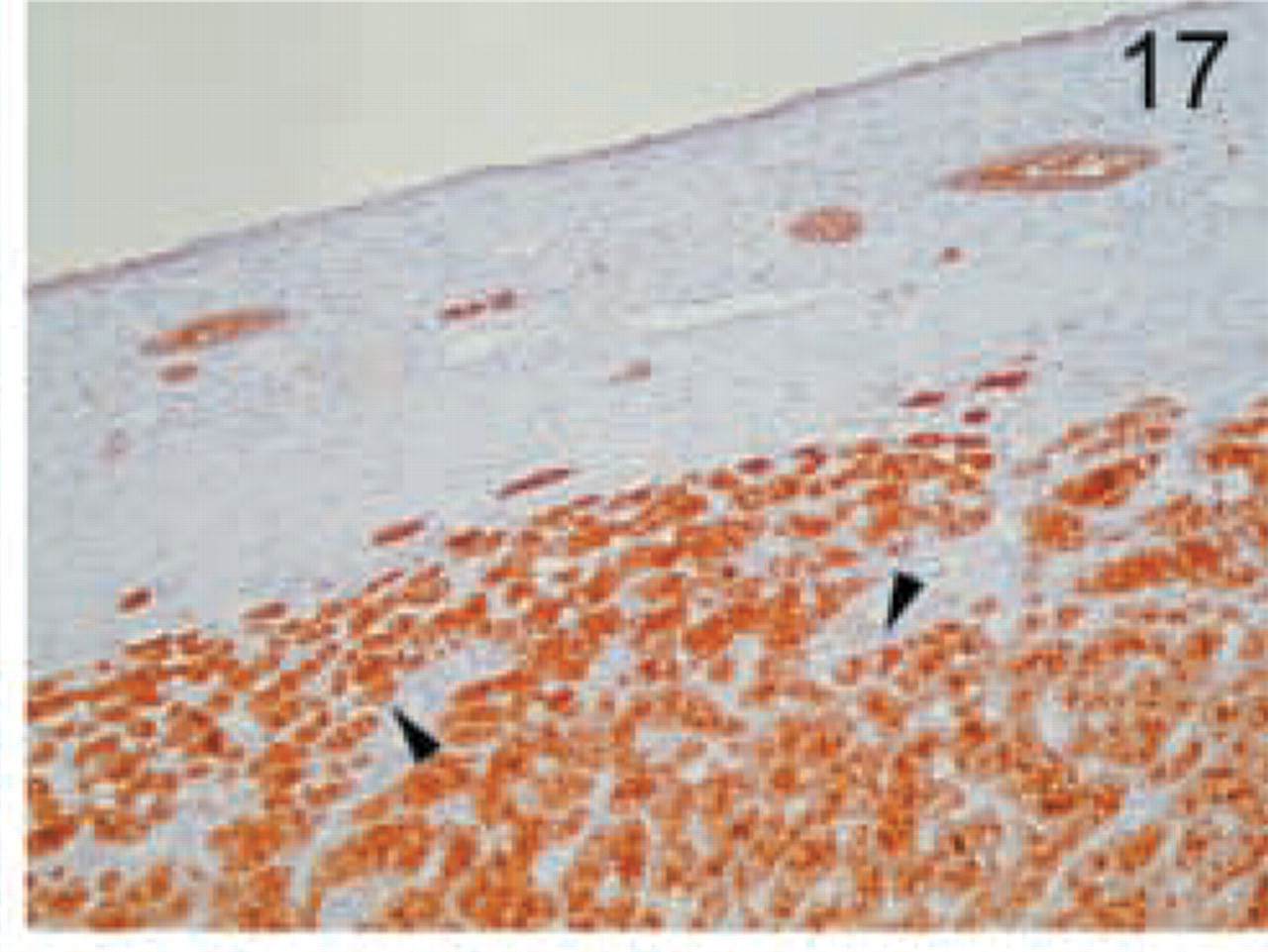
Granular cell carcinoma. Intense granular staining is found in all cells (arrowheads). Original magnification X20.
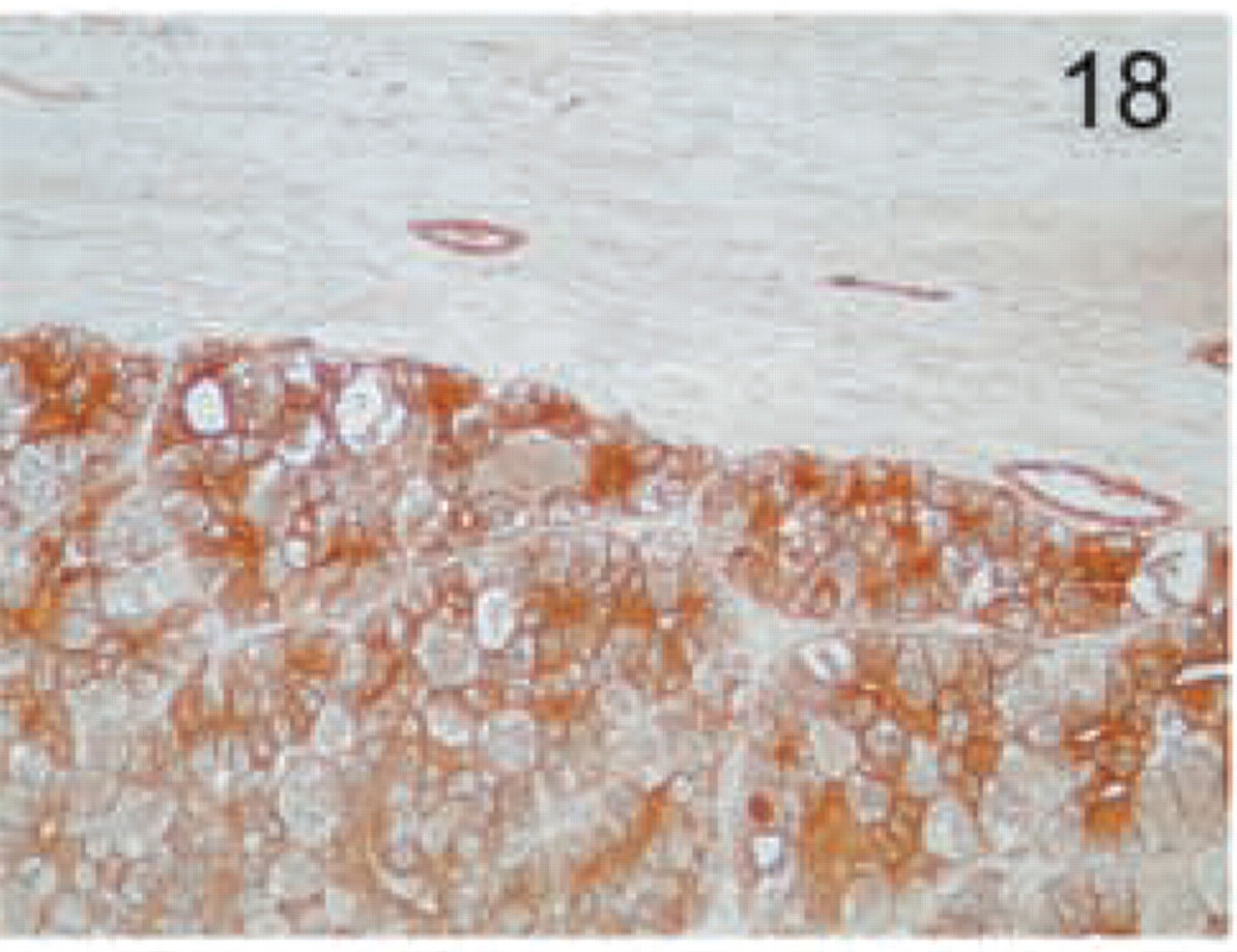
Chromophobe carcinoma. Diffuse pronounced staining is found in all cells, with a variable staining pattern among individual cells. Original magnification X20.
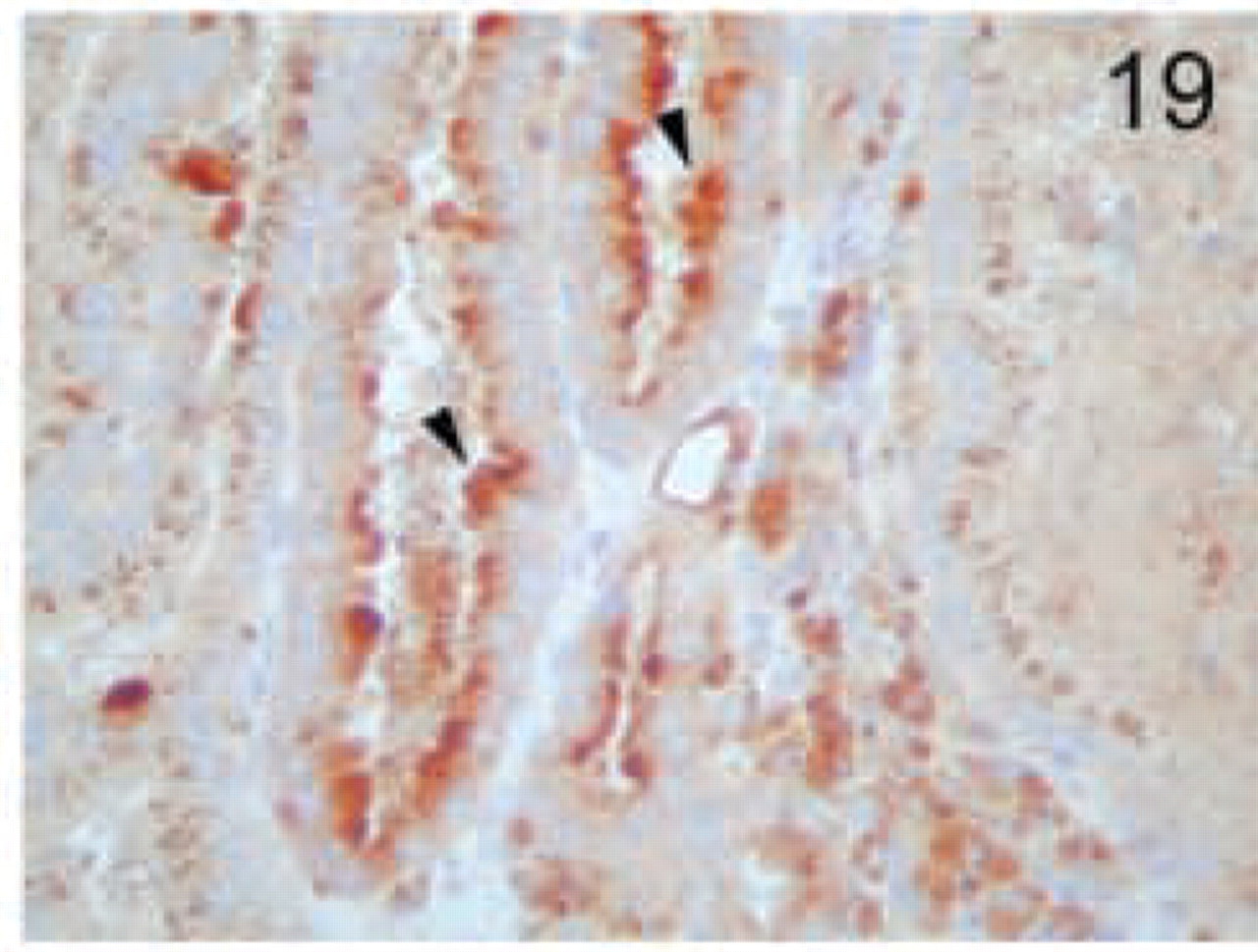
Papillary renal carcinoma. Staining is more pronounced in the apical side, with more pronounced staining in some cells (arrowheads). Original magnification X40.
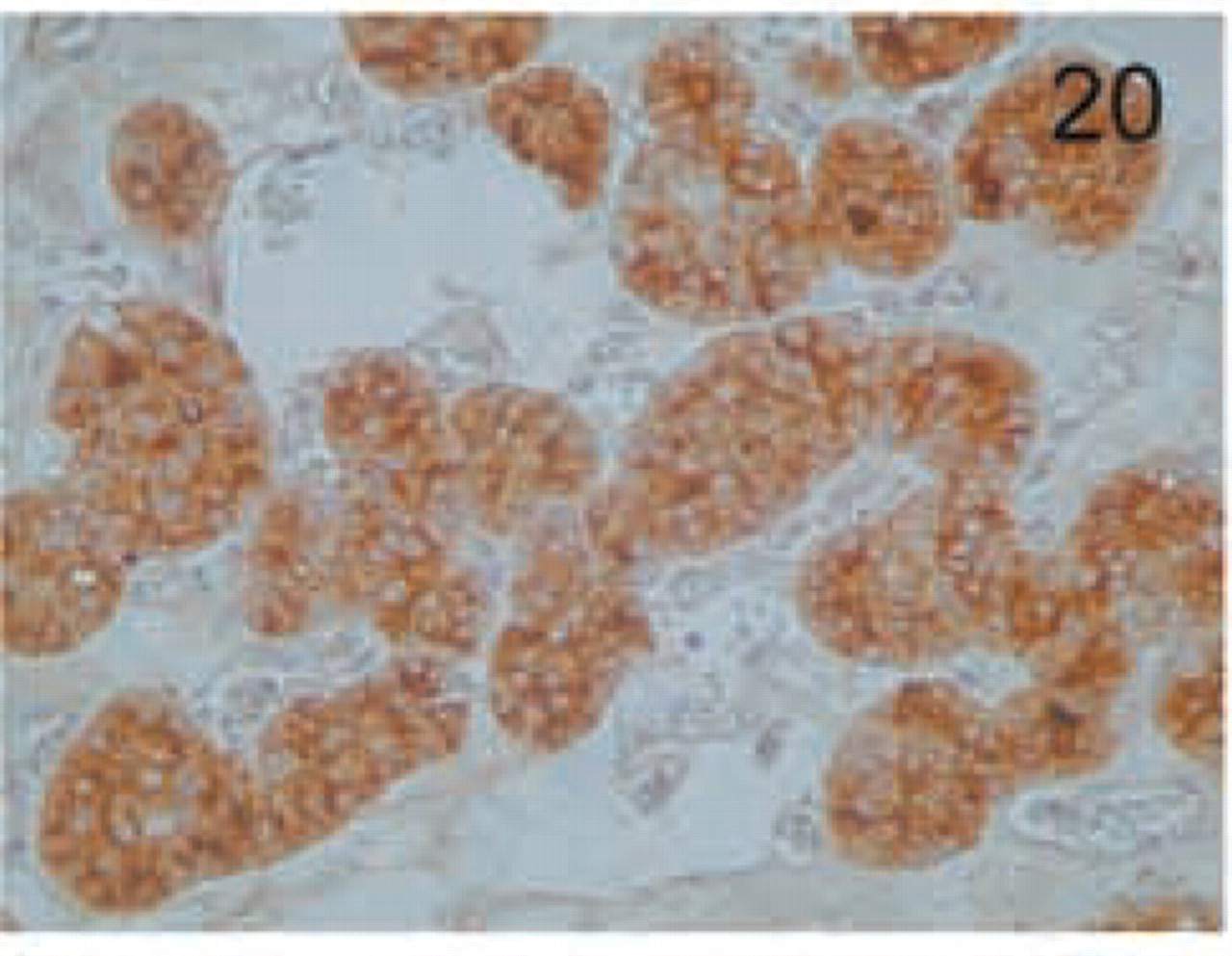
Renal oncocytoma. The epithelium shows intense cytoplasmic granular staining. Original magnification X20.
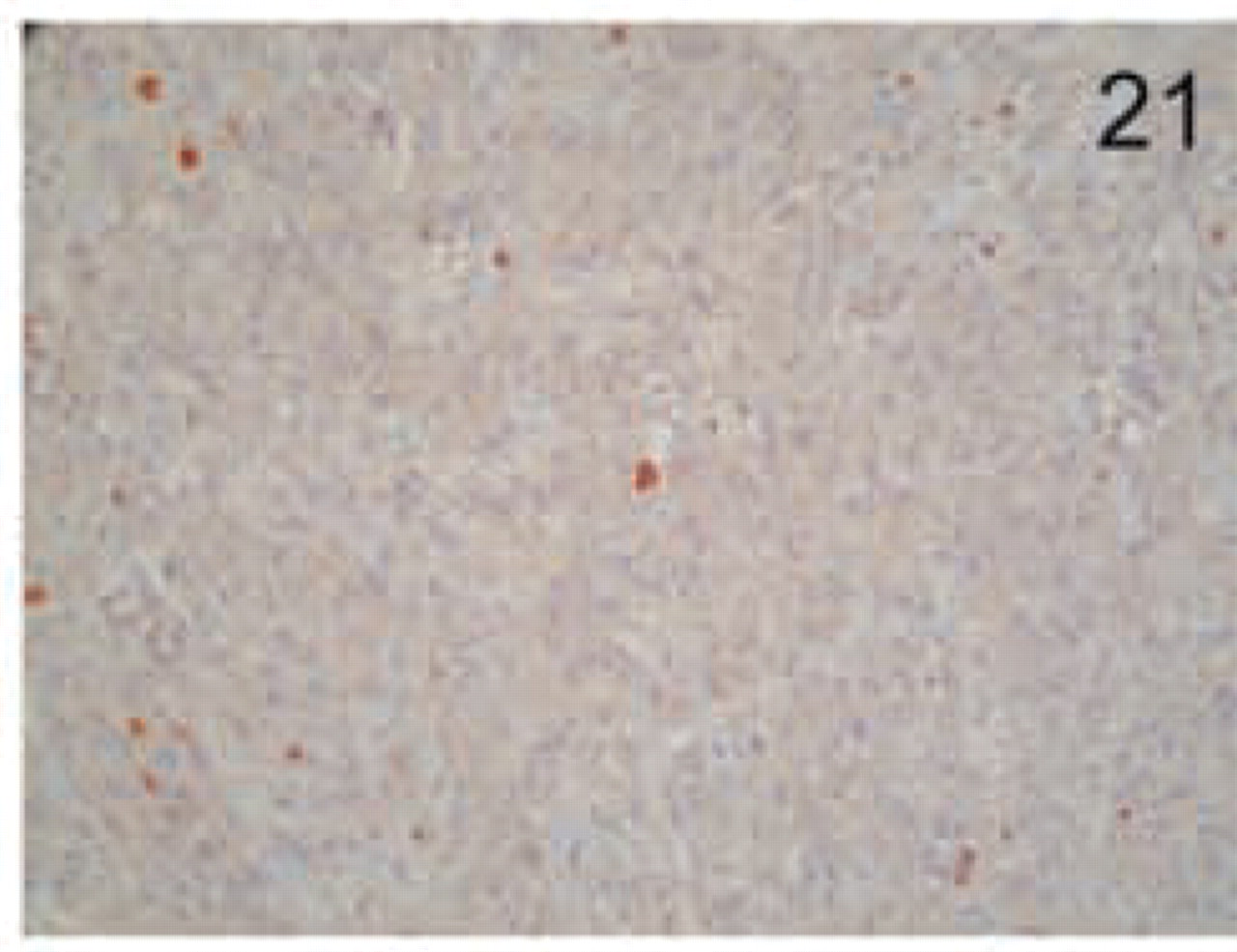
Sarcomatoid carcinoma. No reactivity for MAL is seen. Original magnification X20.
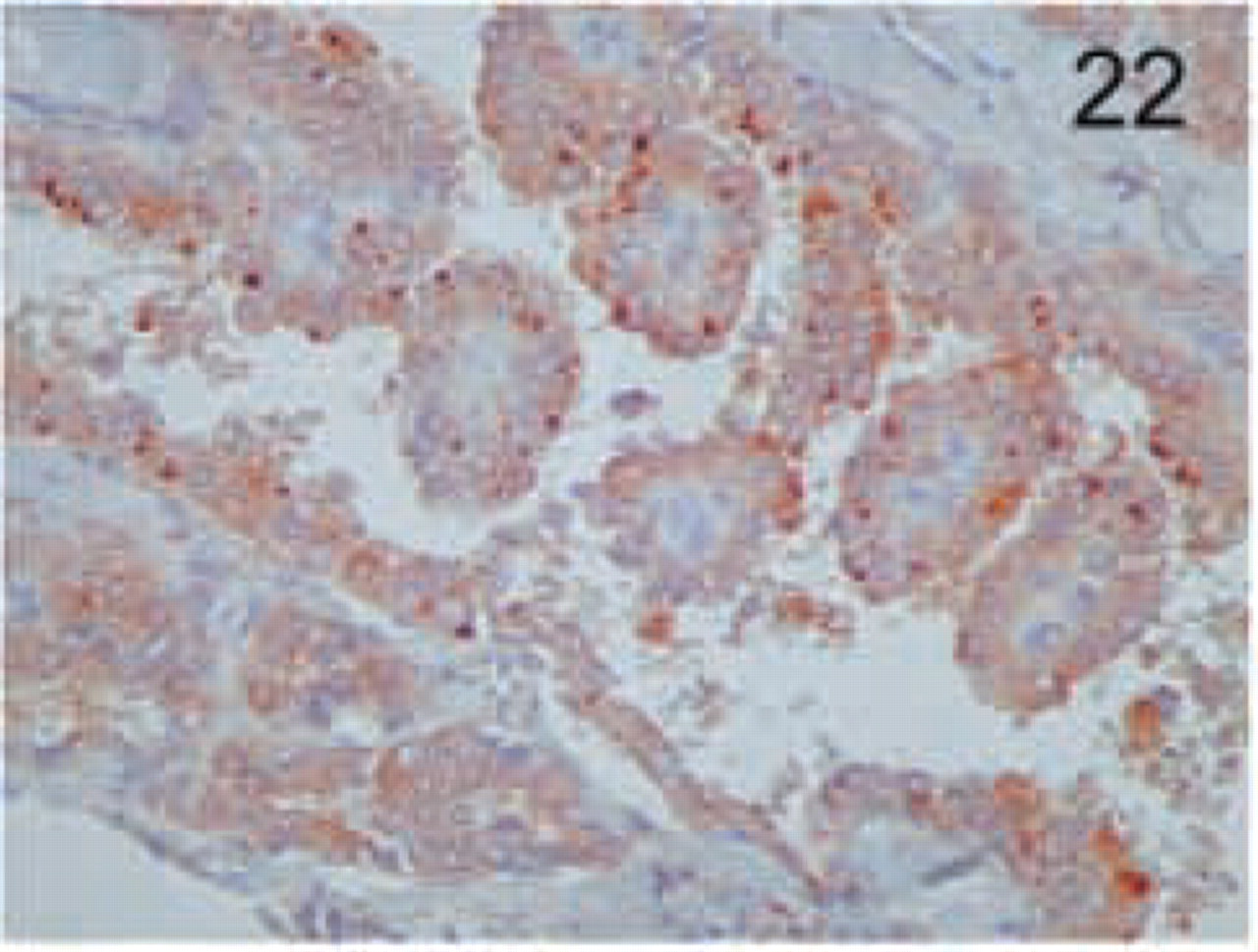
Thyroid tumor of follicular cells: papillary carcinoma. Staining for MAL is found in the apical side, in a granular pattern. Original magnification X20.
Follicular carcinoma. MAL staining is found in the apical side in a diffuse pattern. Original magnification X100.
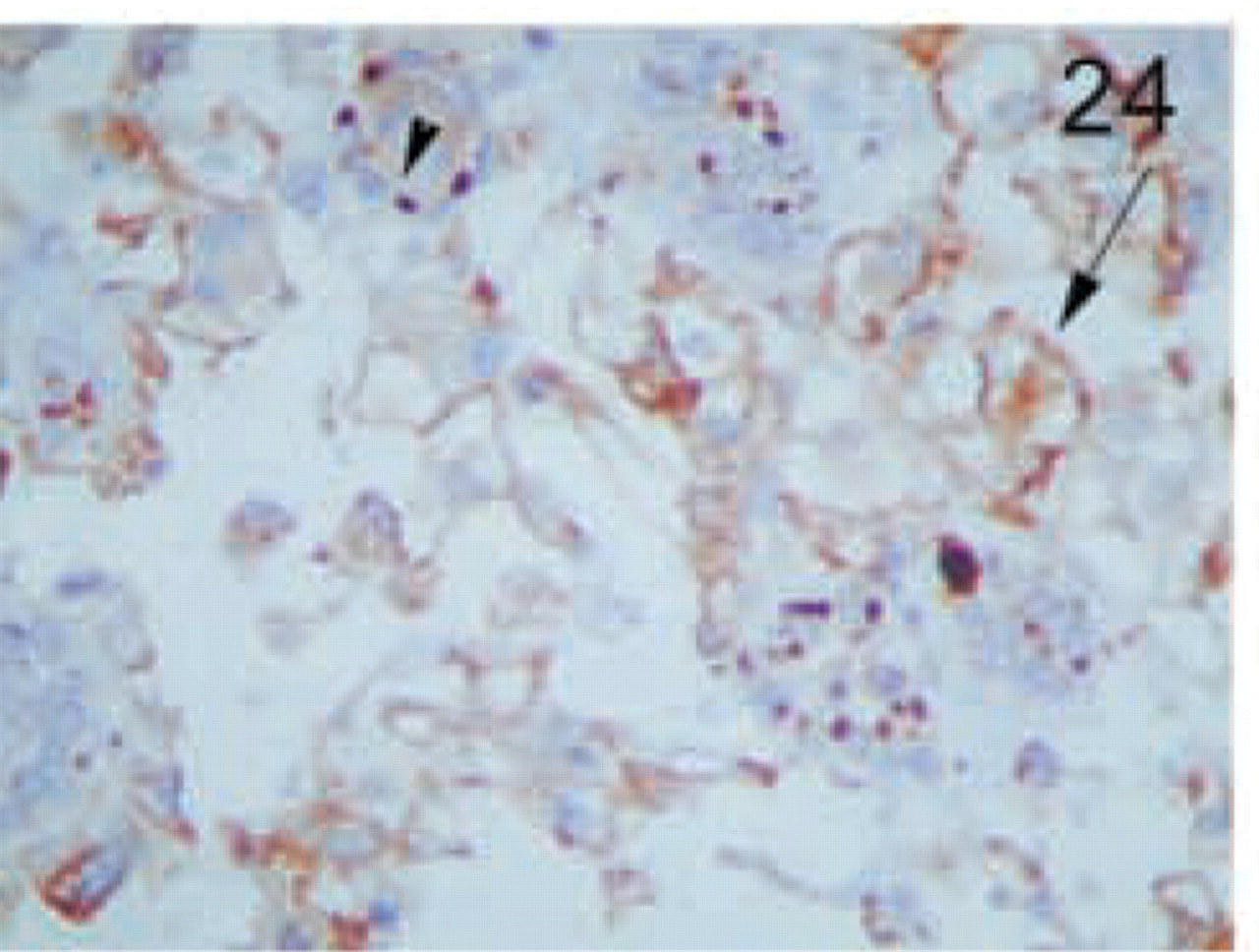
Anaplastic carcinoma. Foci with strong membrane staining are seen (arrow). Compare to the granular apical staining of normal thyroid follicles (arrowhead). Original magnification X100.
In the lung, the ciliated columnar epithelium of bronchi and bronchioles were positive (Figure 11A). Staining was confined to the apical aspect of the cells. In the alveoli, cells lining the alveolar walls were negative (Figure 11B). Nevertheless, large cells with the appearance of type 2 pneumocytes stained strongly with anti-MAL antibody (Figure 11B).
In the endocrine glands, thyroid follicular cells stained strongly and had a characteristic focal granular distribution at the apical side (Figure 12). In the adrenal gland, the medulla showed intense granular staining (Figure 13). Although all the layers of the cortex showed staining, it was stronger in the zona reticularis (Figure 13). In the testis, Leydig cells were strongly positive (Figure 14). In addition, faint staining was found in Sertoli cells but not in spermatogenic cells (Figure 14). The epithelium of the prostate gland showed intense reactivity in its apical aspect (Figure 15).
The distinct types of renal cell carcinoma examined showed different staining patterns. All cases of renal clear-cell carcinoma showed no staining with the MAL antibody (Figure 16). On the contrary, granular cell-type tumors showed intense granular cytoplasmic staining (Figure 17). Cells from chromophobe carcinomas showed intense diffuse staining, which was more pronounced on the apical side and also in some cells compared to others (Figure 18). Papillary renal cell carcinomas showed staining similar to that of normal kidney; staining was more pronounced in some cells of the tubules than in others (Figure 19). Renal oncocytomas showed an intense cytoplasmic staining (Figure 20). Sarcomatoid carcinoma of kidney was negative for MAL expression (Figure 21). Similar to the renal tumors, thyroid follicule-derived carcinomas showed a distinct staining pattern. All cases of papillary carcinoma except one showed staining similar to normal thyroid, more pronounced on the apical side (Figure 22). In follicular carcinomas, staining was found on the apical side in a diffuse pattern compared to the granular staining of normal thyroid cells (Figure 23). In anaplastic carcinoma, only focally stained cells could be seen, with a membrane-staining pattern (Figure 24) different from the granular apical staining of normal thyroid follicles.
Discussion
MAL Expression in Different Types of Human Epithelial Cells
To accomplish their specialized tasks such as diffusion, filtration, absorption, and secretion, epithelial cells adopt different shapes (squamous or flat, cuboidal or columnar) in each tissue and organize as a single (simple) or multiple (stratified) layer of cells. In addition, to protect the body surface and cavities from the harsh external environment, some epithelia are covered by keratin (e.g., skin) or mucus (e.g., respiratory and gastrointestinal tracts). Although we have found representative positive examples of MAL expression in every class of epithelium (Table 1), except for keritanizing stratified squamous epithelia, it is of note that not all other epithelia express MAL. Thus, the simple cuboidal epithelium lining the distal convoluted or collector renal tubules was immunostained for MAL but that of the proximal convoluted tubules showed no detectable staining. One pair of related cell types worth comparing are the flattened endothelial cells of normal blood vessels and the cuboidal endothelium of the high endothelial venules (HEVs). These were found to be negative and positive, respectively, for MAL expression, yet while they have similar roles lining blood vessels and regulating blood coagulation, HEVs in lymphoid organs are the main site for constitutive extravasation during lymphocyte recircularization. Another interesting comparison is that of type 1 and type 2 pneumocytes, the main cellular components of the alveoli. Unlike the thin, flat type 1 cells, which are involved in gas exchange and do not express detectable levels of MAL, round type 2 pneumocytes, which secrete surfactant, a complex mixture of lipid, carbohydrates, and proteins that protects the alveolar surface, were found to express MAL.
MAL as an Element of the Machinery for Lipid Raft-mediated Protein Trafficking in a Restricted Range of Human Cell Types
Specific recruitment of proteins in lipid rafts appears to be a general mechanism for protein segregation and transport used in cells of a wide range of organisms, from mammals (Simons and Ikonen 1997) to yeast (Bagnat et al. 2000). The MAL protein is the first identified member of the integral protein machinery necessary for lipid raft-mediated traffic to the apical surface in the epithelial MDCK and FRT model cell systems (Cheong et al. 1999; Puertollano et al. 1999; Martín-Belmonte et al. 2000,2001). Despite the general use of GEM rafts in protein traffic, we have found that MAL expression is tissue- and cell type-specific. This suggests that the role of MAL in directing membrane trafficking is required in only a restricted range of cells, including many specialized epithelial cell types, secretory cells, T-lymphocytes, and myelin-forming cells. Keeping in mind the demonstrated function of MAL in epithelial MDCK and FRT cells, the obvious role of MAL in the polarized epithelial cells identified as positive for MAL expression is to direct apical transport. Therefore, normal MAL expression might be required for specialized roles (e.g., ion transport by parietal cells, mucous transport by goblet cells, surfactant release by type 2 pneumocytes, expression of specific adhesion molecules in HEVs) that depend on correct protein transport.
The expression of MAL in T-lymphocytes, mast cells, and myelin-forming cells is consistent with the possible existence in these cells of MAL-mediated pathways reminiscent of that of apical trafficking in polarized epithelia. These putative pathways might be related to specialized transport processes that employ GEM rafts, such as myelin biogenesis in myelin-forming cells (Schaeren-Wiemers et al. 1995; Simons et al. 2000) and protein sorting to the uropod protrusion in polarized T-lymphocytes (Millán et al. 2002). In addition, because rafts are also used by T-lymphocytes and mast cells for the assembly of signaling machinery triggered by occupancy of the receptors for the antigen (Montixi et al. 1998) or IgE (Stauffer and Meyer 1997), respectively, the possible involvement of MAL in the activation process in these cells is still an open question (Alonso and Millán 2001).
Our study highlights the knowledge of how apical transport takes place in polarized epithelia that lack MAL expression. MAL was the first identified member of an extended family of proteins (the “MAL proteolipid family”) with structural and biochemical similarities (Pérez et al. 1997; de Marco et al. 2001; de Marco et al. 2002). It is plausible that some of the members of the MAL family play a role similar to that of MAL, acting as machinery in raft-mediated apical traffic, whereas other members possibly participate in novel processes involving lipid rafts.
Potential Applications of the Anti-MAL MAb 6D9
In this study we have shown that the MAL proteolipid is expressed in key cells throughout the human body. The essential role of MAL in raft-mediated traffic and its specific expression in many specialized epithelial cell types, secretory cells, T-lymphocytes, and myelin-forming cells implies that alterations in the expression and/or distribution of MAL would probably be reflected in abnormal functioning of the cells. For example, it has long been known that tumor formation in epithelia leads to disruption of normal cell polarity (Schoenenberger and Matlin 1991). It is plausible that this alteration is accompanied, at least in some cases, by changes in the machinery for protein sorting, the result being that the analysis of MAL expression could aid in detection of incipient neoplasms and improve our undestanding of tumor transformation. The absence of MAL protein in clear-cell renal carcinoma, a histological variant of renal carcinoma related to cytoplasmic accumulation of lipids whose pathogenesis is still unclear, is very interesting. The availability of the human anti-MAL MAb used in this study and its use in both paraffin-embedded section and cryosections may well allow the expression of MAL to be used as a novel tool to characterize a wide variety of human pathologies. The specific immunodetection of MAL in primary mediastinal B-cell lymphoma (Copie-Bergman et al. 1999) offers the first example of a potential diagnostic/prognostic use of the anti-MAL MAb.
Footnotes
Acknowledgements
Supported by grants from the Ministerio de Educación y Cultura (PM99–0092), the Comunidad de Madrid (08.3/ 0025/2000), and Fondo de Investigación Sanitaria (98/109, 01/0085-01, and 01/0085-02). An institutional grant from the Fundación Ramón Areces to CBMSO is also acknowledged.