
Abstract
An intermediate population has been identified among prostate glands called transiently amplifying (TA) cells, which are characterized by coexpression of basal and luminal cytokeratins (CKs), high proliferation, and lack of p27 expression. These cells are rare in the normal adult prostate and increase in pretumoral conditions, but their importance in the developing gland remains unknown. We analyzed fetal prostates for the expression of CKs (5/6, 18, 19) and factors involved in proliferation and apoptosis: p63, Ki67, p27, epidermal growth factor (EGFR), Bcl2, androgen receptor (AR). Immunostaining was performed on a tissue microarray, including 40 prostates from fetuses aged 13-42 weeks and normal prostate tissue from 10 adults. In both solid buds and the basal compartment of canalized glands, cells expressed p63, CK5/6, CK19, CK18, BCL2, EGFR and were p27 negative. Luminal cells of fetal canalized glands continue to express CK19, EGFR, and BCL2, without p27 expression. In contrast, adult epithelial luminal cells showed diffuse AR and p27 expression, without CK19, BCL2, and EGFR staining. Proliferation was high and diffuse in fetal glands and rare and restricted to basal cells in adult glands. These results indicate that most fetal epithelial prostatic cells exhibit the phenotype of TA cells, suggesting their regulatory function in prostate development.
T
The prostate develops from the endodermal urogenital sinus (UGS) following a branching morphogenesis. Prostatic buds emerge at week 10 of gestation from the wall of the UGS and grow in the surrounding mesoderm. During branching morphogenesis of the solid epithelial buds, ductal lumina appear first in the proximal ductal areas near the urethra, then canalization progresses distally toward the ductal tips (Cunha et al. 2004). Two studies of the developing prostrate in both rodents and humans indicate that the majority of epithelial cells coexpress basal and luminal CKs as well as CK19 (Xue et al. 1998; Wang et al. 2001). Other studies observe that numerous epithelial cells in the fetal prostate of humans express both the antiapoptotic factor Bcl2 and EGFR (Leav et al. 1998; Xue et al. 2001b). Taken together, these results suggest that TA cells could represent a predominant population in the developing prostate. We therefore undertook an immunohistochemical study on both fetal and adult human prostates, using CKs markers together with factors involved in cell proliferation and apoptosis, including p27, to further clarify the phenotype of the developing prostate in humans.
Materials and Methods
Tissues
This study was performed after information of the Poitiers University Institutional Review Board on Human Subjects and in accordance with the Helsinki Declaration.
Fetal tissues included 40 prostates taken during autopsy from human fetuses aged 13-42 weeks, without urogenital disorders. Tissue was fixated in 10% formalin <30 min after it was received, and the fixation was continued overnight. Tissue preservation was confirmed by hematoxylin-eosin staining of all samples before immunohistochemistry. Macerated or poorly preserved material was discarded.
For comparative purposes, normal adult prostate tissue was obtained from 10 men aged 62-69 years who were undergoing cystoprostatectomy for urothelial bladder carcinoma and did not have prostate cancer.
After fixation in formalin, tissues were embedded in paraffin, cut in serial 5 μm sections, stained with hematoxylin-eosin, and mounted on slides.
Tissue Microarray
The original slides from the 40 fetal prostates were reviewed by a pathologist to locate the epithelial bud (distal) and canalized glands (proximal). For each case, four cores of tissue (0.6 mm in diameter) were transferred from the selected areas to the recipient block. Serial sections of the tissue microarray (TMA) block were cut 5-μm thick and stained with hematoxylin-eosin to verify that the cores adequately represented the selected areas.
Immunohistochemistry
Immunostaining was performed on the TMA sections and the 10 adult prostate tissue samples. In addition, whole mounted sections from five fetal prostates were analyzed to compare the staining profile and to test the representation of the TMA tissue.
Five-μm tissue sections were placed on charged slides, which were then deparaffinized, rehydrated, and pretreated by either microwave or proteinase K (for EGFR staining) for antigen retrieval. Immunohistochemistry was performed using primary antibodies (Table 1). Briefly, the staining procedure was based on an indirect biotin-avidin system with a universal biotinylated immunoglobulin secondary antibody, diaminobenzidin substrate, and hematoxylin counterstain. A negative control slide was obtained after either omitting the primary antibody or incubating with an irrelevant antibody (mouse monoclonal immunoglobulin).
Scoring of Antibody Staining
For all antibodies except Ki67, epithelial staining was assessed as either diffuse or focal with weak or high intensity. Focal staining is defined as rare scattered positive cells among negative cells within the same epithelial compartment. For Ki67, positive cells were expressed as a percentage of total epithelial cells. The staining profile was assessed for both buds and canalized ducts. For canalized glands, staining was analyzed in both the basal and luminal compartments. In whole mounted sections of fetal prostates (n=5) containing in the same section buds and canalized glands, a distinction was made between the staining pattern of proximal canalized glands and distal ducts (close to buds).
Statistical Analysis
Comparison of Ki67 staining between groups was analyzed using the Student's t-test.
Results
The epithelial staining profile was identical in whole mounted sections of fetal prostates and TMA cores from the same fetuses. The staining pattern was also identical in all solid buds, and in all canalized glands, whatever the gestational age. However, tiny differences of staining were observed between distal and proximal canalized glands.
Characteristics of antibodies used
CK, cytokeratin; EGFR, epidermal growth factor receptor; PSA, prostate-specific antigen; AR, androgen receptor.
Staining results except for Ki67 are summarized in Table 2.
Expression of CK5/6 and p63 was found in solid buds and all basal cells in both fetal and adult glands. CK5/6 and p63 were not expressed in luminal cells. CK19 staining was observed in all epithelial fetal cells, but was restricted to the basal compartment in adult glands (Figure 1). CK18 staining was intense in all cells of both buds and distal fetal glands, but the intensity decreased in the basal compartment of proximal fetal glands and adult glands, whereas the staining remained intense in the luminal compartment. AR positive cells in fetal prostate were found only focally in the luminal compartment of the proximal glands, whereas the staining was diffuse in adult luminal cells. EGFR and Bcl2 positively displayed the same profile: positive staining was observed in solid buds and basal cells of fetal and adult glands. Luminal cells were diffusely positive in distal fetal ducts, only focally positive in proximal fetal ducts, and negative in adult glands (Figure 2). Nuclear p27 positive staining was absent in all epithelial cells of fetal prostates, with a positive internal control on neural ganglion cells adjacent to the prostate. Moreover, p27 was also expressed in other fetal tissues present on the same TMA. In adult epithelial cells p27 expression was observed diffusely in the luminal compartment, and focally in the basal compartment (Figure 3).
In solid buds, Ki67 staining ranged from 10% to 45%, with a median of 21% (Figure 4). In canalized glands, the median Ki67 percentage of positive cells was 8% (6% to 17%), with a decrease in proliferation when compared with solid buds (p<0.0004). Proliferative cells in canalized glands were found in both basal and luminal compartments (Figure 4). In adult prostatic glands, the epithelial proliferation rate was always <5%, and proliferative cells were restricted to the basal compartment. In solid buds, the median Ki67 staining was 28.5% in fetuses <23 weeks, and 20% in older fetuses (>23 weeks), but the difference was not significant (p = 0.07). The median Ki67 staining in canalized glands was 10% in fetuses <23 weeks of age, and 7% in older fetuses (p = 0.3).
Epithelial expression profile in developing and adult prostate
CK, cytokeratin; AR, androgen receptor; PSA, prostate-specific antigen; EG FR, epidermal growth factor receptor; +, diffuse intense staining; ±, diffuse weak staining; −, no staining; +f, focal staining.
Discussion
TMA technology allows for the simultaneous examination of numerous tissue samples for in situ gene-expression profiling, with dramatic savings in reagent and technical costs and a gain in quality insurance. Moreover, TMA construction uses only small quantities of tissues, allowing for the study of rare or scarce biological samples with many molecular markers (Packeisen et al. 2003). This technology could therefore be suitable for the study of fetal tissues. However, the main drawback of the technique is the problem of the representation of TMA cores compared with corresponding conventional whole sections. This representation depends on both the type of tissue and the antibodies tested. We demonstrated that when the cores were selected in representative areas (i.e., buds and canalized glands), the expression profile of the antibodies tested was identical in TMA and whole sections. This suggests that TMA could be an interesting tool for high-throughput molecular analysis of fetal prostate.
Using Ki67 staining, we observed that solid buds display a significantly higher proliferative rate than canalized glands, as previously described (Xue et al. 2001b). After 23 weeks of gestation, epithelial proliferation was lower in both buds and canalized glands (20% and 7%, respectively) when compared with younger fetuses (28.5% and 10%, respectively), but the difference was not significant, in contrast with other studies (Xue et al. 2001). In addition, we observed that even after 30 weeks of gestation, the proliferative activity in canalized glands was not restricted to the basal compartment (as it was in adult glands) but was also present in the luminal compartment. This supports the hypothesis that numerous TA cells are present in the fetal prostate, which are also highly proliferative (De Marzo et al. 1998).
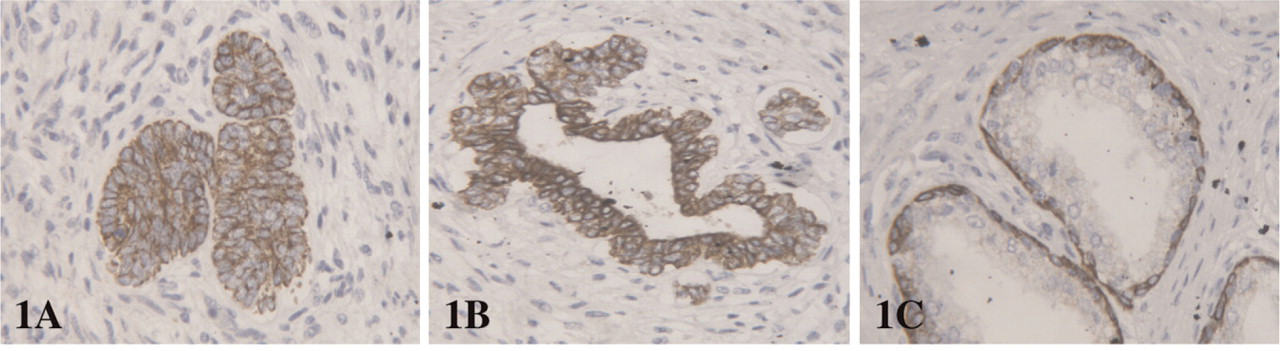
CK 19 expression in (
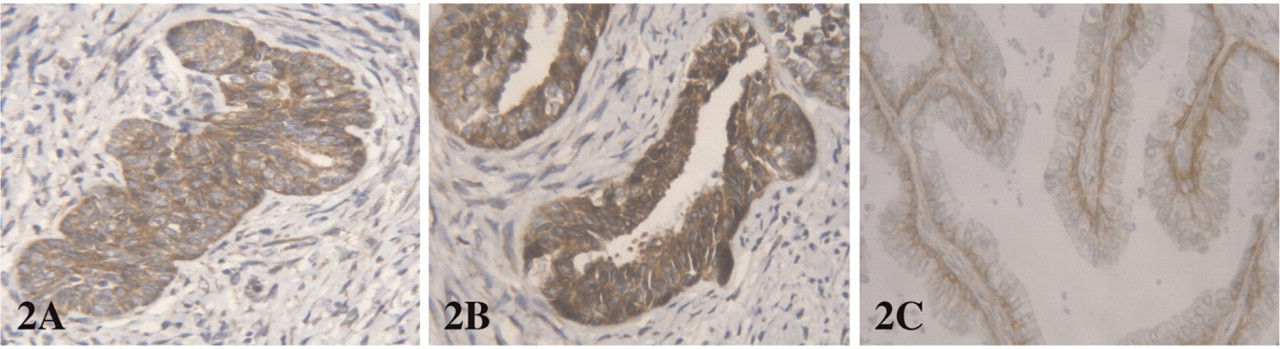
Bcl2 expression in (
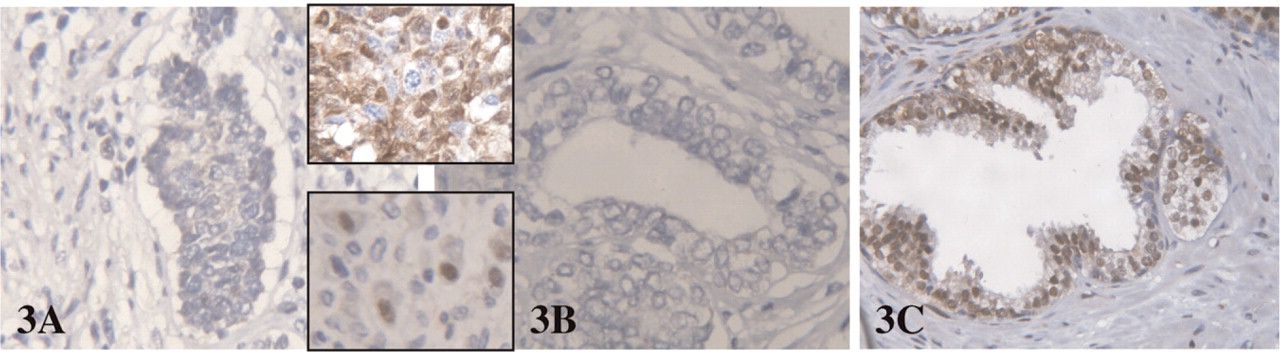
Lack of p27 expression in buds and canalized fetal gland (
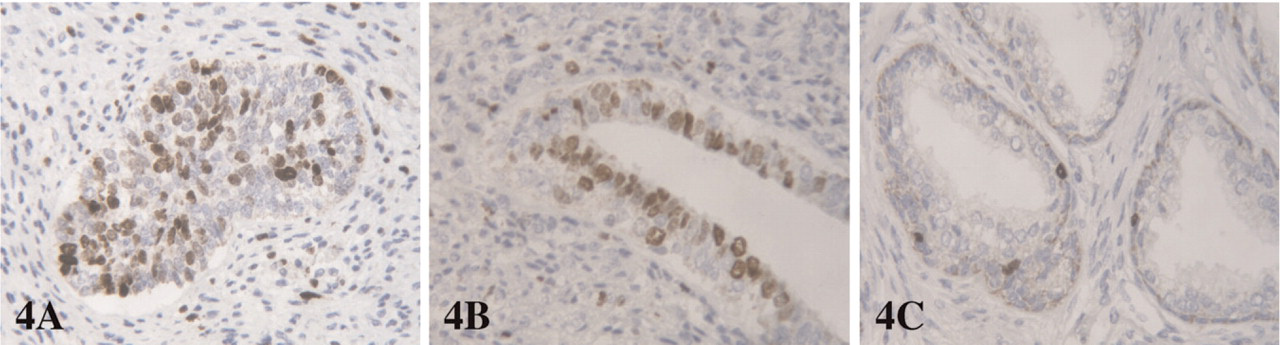
Ki67 staining in (
Very few studies have reported the in situ expression of differentiation markers in the fetal prostate, probably because material of developing human prostate is rarely available. We tested 40 fetuses simultaneously for several differentiation markers, including CKs, together with factors involved in proliferation and apoptosis. We found that, for all antibodies, the expression profile was identical in all buds, irrespective of gestational age. In the same way, the only small differential expression that was observed in canalized glands was between distal and proximal glands, whatever the level of fetal development. Our finding demonstrates that the degree of differentiation in human epithelial prostatic cells depends upon the phase of canalization and not upon the gestational age.
In our study, the basal cell markers CK5 and p63 were expressed in buds and in the basal compartment of both fetus and adult glands and were lost in the luminal compartment even at the early phase of canalization. In contrast, other studies found focal persistence of CK5 and p63 in the luminal fetal cells of distally canalized glands (Xue et al. 1998; Wang et al. 2001). This discrepancy could be due to a difference in the antibodies tested, insofar as the previous studies used a cocktail of high molecular mass CKs instead of CK5/6. Moreover, the staining pattern of p63 was analyzed by immunofluorescence, and the authors did not distinguish between basal and luminal compartments in early canalized glands (Wang et al. 2001).
We observed that the luminal cell marker CK18 was strongly positive in all cells of both buds and distal glands, as previously described (Hudson et al. 2000). However, in contrast to a previous study (Sherwood et al. 1990) but in accordance with others (van Leenders et al. 2000), we found, using a commercially available monoclonal antibody, that CK18 expression remains positive, although less intense, in the basal compartment of both proximal fetal glands and adult glands. This data suggests that CK18 could not be a highly specific marker of terminally differentiated luminal cells.
CK19 is thought to represent a population in the process of differentiation into luminal cells and has been shown to be lost in terminally differentiated cells (Hudson et al. 2000, 2001). In fact, we found that CK19 expression in adult glands was restricted to the basal compartment. This staining pattern was not identical in the fetal prostate, because all cells, even in the luminal compartment of proximal glands, were strongly positive for CK19. This expression profile suggests that no complete luminal epithelial differentiation is achieved at birth in the developing prostate. This hypothesis is supported by the fact that androgen receptor expression, which was diffuse in adult luminal cells, was only focal in the luminal fetal compartment.
The anti-apoptotic factor Bcl2 and the EGFR have been shown to be present in basal cells and down-regulated during differentiation, without expression in normal luminal adult cells (McDonnell et al. 1992; Ibrahim et al. 1993). In fetal tissues, despite a decreased and focal staining in proximal glands, the great majority of epithelial cells expressed both Bcl2 and EGFR, which can confer them a resistance to apoptosis and promote proliferation.
The cyclin-dependant kinase inhibitor p27 has been implicated in cell cycle arrest, and increased level of p27 is thought to signal an exit from the cell cycle (Roberts et al. 1994). In adult prostatic glands, p27 was expressed by all cells in the luminal compartment and by a majoritary subpopulation of basal cells (De Marzo et al. 1998). P27 downregulation occurs not only in most prostate cancers (Tsihlias et al. 1998) but also in high grade PIN (De Marzo et al. 1998) and in putative pretumoral conditions such as proliferative inflammatory atrophy (van Leenders et al. 2003). The lack of p27 expression in prostate epithelium has been suggested to be a specific indicator for TA cells in the non-tumoral prostate (De Marzo et al. 1998). We also found p27 expression in all luminal cells and numerous basal cells from adult prostatic glands. In contrast, no p27 staining was observed in epithelial cells of the fetal prostate, whereas expression was found in fetal neural ganglion cells adjacent to the prostate and in other fetal tissues present on the same TMA.
In conclusion, our study demonstrates that a majority of epithelial cells in developing prostates showed coexpression of CK18 and 19, basal cell markers such as EGFR and Bcl2, a high proliferative rate, and a lack of p27 staining. This expression profile suggests that the epithelial population in fetal prostates is made up mostly of TA. Because TA could be potential targets for neoplasic transformation but are rare in the normal adult glands, further analysis of fetal TA cells may provide a better understanding of prostate carcinogenesis.