
Abstract
Amelogenins and tuftelins are highly specialized proteins secreted into the developing enamel matrix during mammalian enamel formation. Both tuftelins and amelogenins have been associated with various functions during nucleation and maturation of the developing enamel matrix. In this study we conducted experiments to investigate whether tuftelins and portions of the amelogenin molecule were deposited and processed in spatially distinguished portions of the developing enamel matrix, using antibodies specific against tuftelin or amelogenins. The amelogenin antibodies were raised against recombinant and native amelogenins and also included an antibody against a polypeptide encoded by amelogenin exon 4. To compare spatial expression patterns of enamel protein epitopes, 3-day postnatal mouse molar tooth organs were processed for paraffin histology and cut into serial sections. Adjacent sections were exposed to antibodies against either tuftelin or various amelogenin epitopes. To investigate age-related changes of enamel protein expression, amelogenin and tuftelin antibodies were applied to tooth organs of developmental stages E19 and 1, 3, 5, 7, 9 and 11 postnatal days. Tuftelin was detected within the odontoblast processes during earlier stages of development (E19 and 1 day postnatal), whereas during later stages (3–11 days) it was recognized in a portion of the enamel layer adjacent to the dentine–enamel junction. In contrast, all four antibodies against amelogenins reacted with parts of the ameloblast cytoplasm and the entire enamel layer. Using immunohistochemistry, we were not able to detect any differences in the spatial distribution of the four amelogenin epitopes investigated. The spatial differences in the distribution of amelogenin and tuftelin as observed in this study may be intepreted as an indication of functional differences between both proteins during early enamel biomineralization.
S
The identification of several different proteins within developing enamel as well as the discovery of alternatively spliced amelogenin gene products and post-translational modifications (Simmer 1995; Simmer et al. 1994a; Fincham et al. 1991) suggests that enamel formation may be an intricate microprocess in which all of these components, as well as post-translational modifications and splicing mechanisms, play important functional roles during the formation of long and parallel-oriented enamel crystals. However, the presence of various proteins and spliced products by itself does not automatically imply their functional importance. Some of them may be relics of evolutionary primitive enamel/enameloid formation, others may be degradation products, and others even may have only been produced in vitro as PCR artifacts. At present, experimental strategies used to investigate the function of various protein components of developing enamel during crystal orientation and growth include anti-sense and transgenic mouse production experiments (Chen et al. 1994; Diekwisch et al. 1993). These experiments provided evidence for a significant role of amelogenins during early enamel biomineralization. Further support of the importance of the entire amelogenin molecule for enamel crystal formation is provided by previous studies (Lench and Winter 1995; Aoba et al. 1987a, b).
Several authors have distinguished between phases of enamel crystal formation, mainly nucleation, elongation, and maturation (Diekwisch et al. 1995; Simmer and Fincham 1995; Sasaki 1990). The apparent differences in crystal form and size between these phases suggest that different molecules or portions of molecules might be associated with different stages of enamel development, e.g., tuftelins might be associated with enamel crystal nucleation and parts of the amelogenin molecule might be associated with enamel crystal maturation (Deutsch 1989). Functional differences between different portions of the amelogenin molecule can be concluded from hydrophobicity plots (Fincham et al. 1992). Therefore, we were interested to compare immunohistochemically the spatial distribution of enamel proteins during tooth development by taking advantage of the recent availability of several antibodies against tuftelin and amelogenin as well as amelogenin cleavage products. We hypothesized that differences in the spatial distribution of amelogenins and tuftelin as well as developmental changes in the spatial distribution of any enamel protein might exist. To test these hypotheses, we designed experiments to compare the distribution of amelogenin and tuftelin at a similar stage of mouse first mandibular molar enamel formation. We further studied changes in antigenic distribution of amelogenin and tuftelin during mouse molar enamel formation. Our studies indicated striking similarities in the spatial distribution of all amelogenin antigens investigated, significant differences between the distribution of amelogenin and tuftelin, and developmental changes in the distribution of amelogenin and tuftelin.
Materials and Methods
Animals
Timed pregnant mice and female Swiss-Webster mice with litters were purchased from Charles River (Wilmington, MA). For developmental studies, females with litters of embroyonal stage E19 as well as young mice 1 day, 3 days, 5 days, 7 days, 9 days, 11 days and 13 days postnatal were sacrificed. For comparative studies, 3-day postnatal mice were used. At each developmental stage, three animals were studied. Mice were sacrificed in a humane manner by cervical decapitation according to Baylor College of Dentistry Animal Care Committee recommendations.
Tissue Preparation
Mandibles were dissected and tooth molar organs separated for optimal fixative penetration. Tissues were fixed for 1 day in 10% buffered formalin, decalcified for 2 days in EDTA solution according to Warshawsky and Moore (1967), dehydrated in a graded series of ethanols, and embedded in paraffin. Paraffin blocks were cut into serial sections of 5 μm. For comparative analysis of immunoreactivity with different antibodies, ribbons of six adjacent serial sections were placed on cardboard and separated into single sections. Each single section was stretched on a water bath and mounted on a glass slide coated with poly-
Immunohistochemistry
Immunoreactions were performed following the instructions of the Zymed Histostain SP kit (Zymed; So. San Francisco, CA). All reactions were carried out in a humidified chamber at room temperature. Briefly, sections were treated against endogenous peroxidase using methanol and 3% hydrogen peroxide and then were blocked with 10% goat serum for 10 min. Sections were incubated with primary antibody (amelogenins or tuftelin) for 2 hr. Primary antibodies were diluted in PBS. The dilution of the primary antibody was determined in several preliminary experiments. A dilution was called optimal when a strong signal but no background was detected. As a methodological control, the primary antibody was replaced with PBS. Sections were washed three times in PBS and then incubated for 10 min with biotinylated goat anti-rabbit IgGs as secondary antibodies. After washing in PBS (three times), sections were exposed to the streptavidin–peroxidase conjugate for 10 min and then washed again in PBS (three times). Signals were detected using the AEC Substrate–Chromogen mixture of the Zymed Histostain kit. Sections were counterstained using hematoxylin and mounted with GVA mount.
Primary Antibodies and Dilutions
1. Polyclonal antibody against a recombinant mouse amelogenin (M179), IgG preparation; dilution 1:100 (Simmer et al. 1994b). The antibody stained the enamel and the ameloblast layers in an immunohistochemical study and recognized the 22-kD amelogenin on Western blots (Simmer et al. 1994b).
2. Polyclonal antibody against a mouse exon 4-encoded polypeptide. Exon 4 is expressed only in the largest mouse amelogenin M 194; dilution 1:500 (Simmer et al. 1994a). This antibody reacted against the enamel and the ameloblast layers in an immunohistochemical study and recognized amelogenin extracts on Western blots (Simmer et al. 1994a).
3. Polyclonal antibody against enamel tooth protein extracts separated by gel electrophoresis. This antibody is directed against 10–22-kD tooth proteins excised from slab gels containing mainly amelogenins; dilution 1:1000 (Slavkin et al. 1982). This antibody reacted against ameloblasts and the entire enamel layer on paraffin sections and recognized a 26-kD band on Western blots.
4. Polyclonal antibody against the C-terminal 25 amelogenin amino acids of the pig 173 residue amelogenin (courtesy of Dr. Aoba, Forsyth Dental Center, Boston); dilution 1:100 (Aoba et al. 1992).
5. Polyclonal anti-tuftelin antibody against a synthetic polypeptide derived from the mouse tuftelin sequence. The peptide sequence used for antibody production was QSKDTTIQELKEKIA; dilution 1:50 (Zeichner-David et al., submitted for publication).
6. Antibody control performed by replacing the primary antibody with normal serum at a dilution of 1:50. Further controls were performed using pre-adsorbed primary antibodies for tuftelin and amelogenins as well as controls using normal serum.
Sample Analysis
To guarantee consistency and comparability in the sampling area for all five antibodies investigated, the center of the mesial margin of the mesiobuccal cusp of first mandibular mouse molars was selected for analysis and photodocumentation. Photomicrographs were taken on a Zeiss Axiophot using a X63 Planapochromat objective and DIC Normarski contrast. Figures 1A-1F and 2E-12H were taken on Kodachrome 40 film; Figures 2A-2D were photographed on Fujichrome RTP64T since Kodachrome was no longer available.
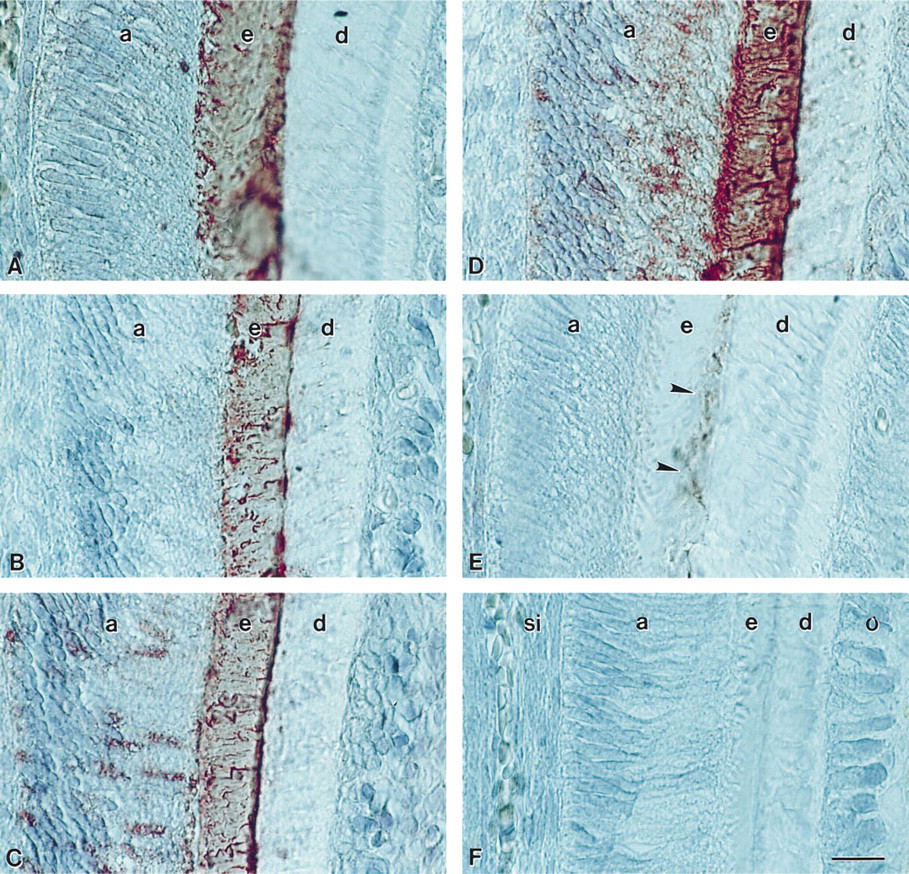
Enamel protein localization in the developing enamel matrix of a 3-day postnatal first mandibular molar mesiobuccal cusp. (
Results
Using conventional immunohistochemistry, we detected significant staining patterns for amelogenins and tuftelin in the developing enamel of first mandibular mouse molars. We compared adjacent sections reacted with various antibodies against amelogenin and tuftelin and we analyzed developmental changes in the spatial distribution of amelogenins and tuftelins.
Amelogenin epitopes were localized in the entire enamel layer and parts of the ameloblast cell body, whereas tuftelin epitopes were detected at the dentine–enamel junction of 3-day postnatal mouse molars. Analysis of adjacent sections of the mesial ridge of the mesiobuccal cusp of 3-day postnatal mouse molars revealed striking similarities in the staining pattern of the four amelogenin antibodies investigated in this study, as well as a different staining pattern for tuftelin. All four antibodies against amelogenin (M 179, exon 4, SDS gel 22-kD extract, C-terminal 25 amelogenin amino acids) reacted with the entire enamel layer (Figures 1A–1D). A weak reaction was also detected in the apical half of the ameloblast cytoplasm. The anti-tuftelin antibody reacted with a small portion of the enamel layer adjacent to the dentine–enamel junction (Figure 1E). No reaction was observed in all controls (Figure 1F).
During postnatal mouse molar enamel formation, tuftelin signals moved from the odontoblast process towards the dentine–enamel junction. At E19, which was the earliest stage we analyzed, tuftelin-immunoreactive products were located at the apical odontoblast pole (Figure 2A). In 1-day postnatal mouse molars, tuftelin immunostaining was documented in the odontoblast process and in the apical pole of the odontoblast cells. There was also a weak reaction in the ameloblast cells (Figure 2B) and in the stratum intermedium (Figures 2B and 2C). In 3-, 5-, 7-, 9-, −11, and 13-day postnatal mouse molars, tuftelins were located at the dentine–enamel junction of the developing enamel layer (Figures 2C, 3 days and 2D, 11 days) (data for 5, 7, 9, and 13 postnatal days not shown). The tuftelin reaction product formed a fine line parallel to the dentine–enamel junction on the side of the enamel layer.
During postnatal mouse enamel formation, amelogenin signals were mainly detected in the enamel layer. For the developmental analysis, immunostainings were performed using the antibody against the recombinant mouse amelogenin molecule M179 expressed in E. coli. Signals for amelogenin were detected in granules within the ameloblasts and at the initial enamel layer of E19 embryonal stage mouse molars. Isolated reaction products were also seen in the dentine layer (Figure 2E). In 1-day postnatal mouse molars, amelogenin expression products were located at the apical pole of the ameloblasts and in the enamel layer (Figure 2F). In mouse molars older than 3 postnatal days (3–13 days; data shown only for 3 and 11 days), amelogenins were located in the developing enamel layer (Figures 2G, 3 days and 2H, 11 days) as well as in the cytoplasm of ameloblasts (Figure 2G, 3 days).
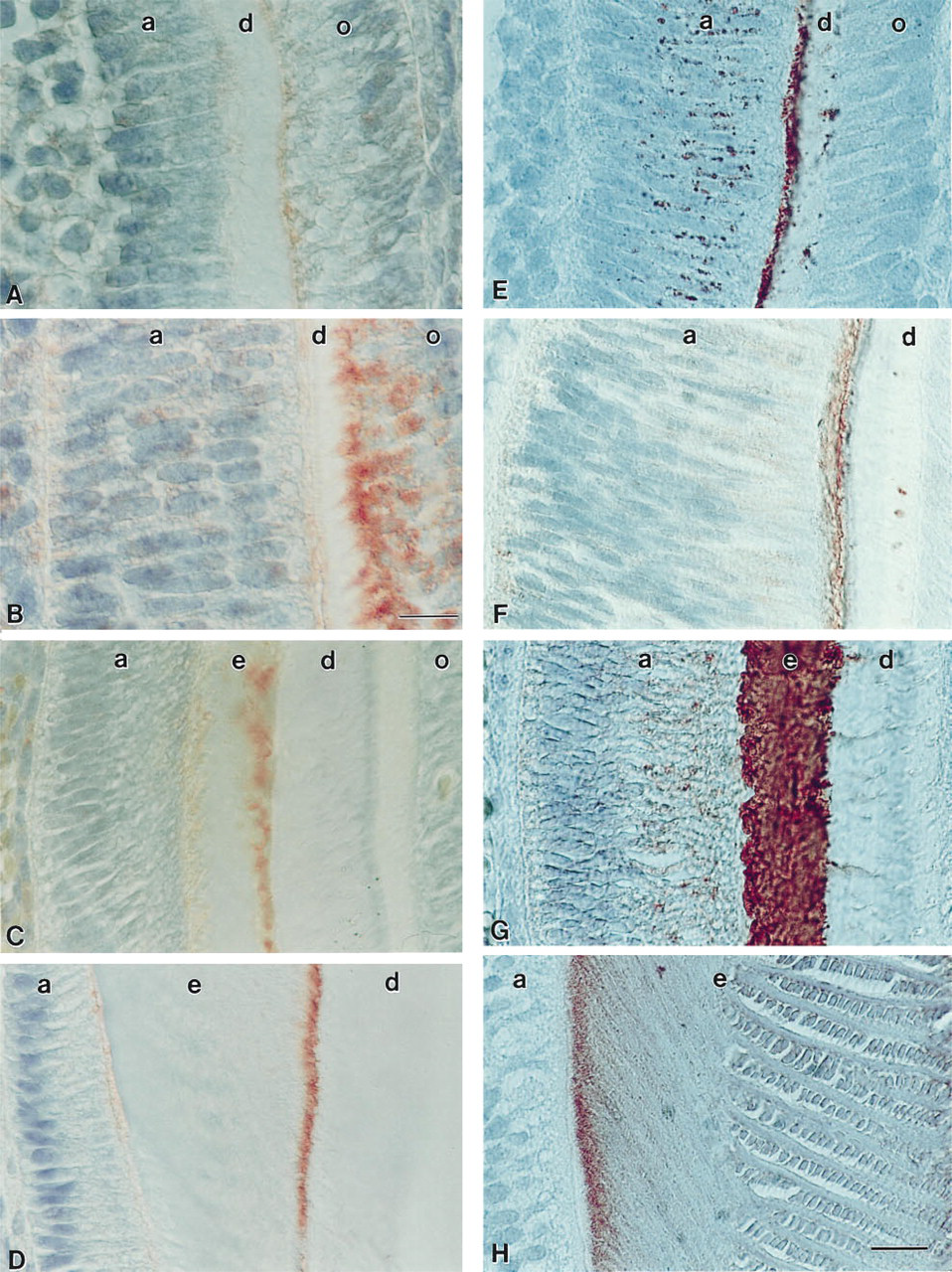
Tuftelin (
Discussion
In the present study we compared the immunohistochemical distribution of amelogenins and tuftelin in 3-day postnatal mouse molars. We also analyzed changes in the distribution in both classes of enamel proteins during postnatal mouse molar development. Our studies indicated striking similarities in the spatial distribution of all amelogenin antigens investigated, significant differences between the distribution of amelogenin and tuftelin, and developmental changes in the distribution of tuftelin and amelogenin. Specifically, tuftelin localization appeared to change from the odontoblast layer of early secretory stage towards the dentin–enamel junction of later developmental stages (Figure 3).
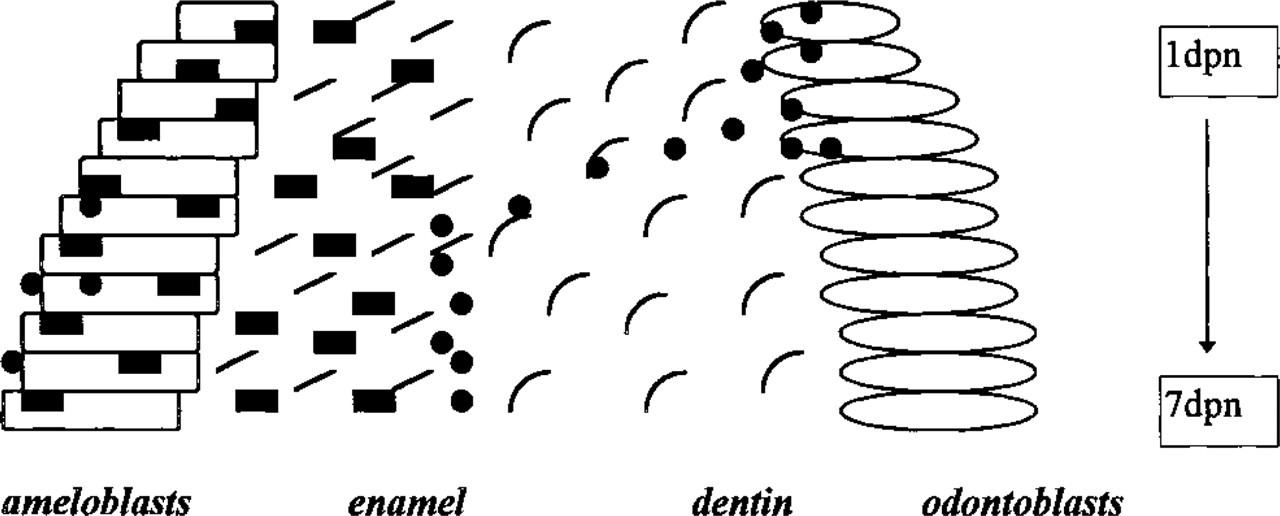
Schematic view of changes in the distribution of tuftelin and amelogenin during tooth mineralization. Amelogenins (▪) were distributed in the ameloblast and enamel layer throughout enamel formation. Tuftelin distribution (•) changed from the odontoblast cell layer to the dentine-enamel junction. Vertical scale indicates days of postnatal development (dpn).
All antibodies for amelogenins reacted with the entire enamel layer at 3 postnatal days. This finding confirms previous observations (Simmer et al. 1994a; Nanci et al. 1985; Slavkin et al. 1982) in which amelogenin was located at the onset of enamel biomineralization. Staining patterns of the antibodies raised against recombinant and those raised against native amelogenin proteins were very similar, indicating similarities between recombinant and native amelogenins at the immunohistochemical level. One might have expected that the C25 antibody would react only to the newly formed enamel, because this sequence is removed early during amelogenin post-translational processing. However, we did not detect any differences in staining intensity throughout the enamel layer. The reaction product in the enamel layer was significantly stronger than the signal we detected in the ameloblast cell bodies. The differences in intensity may be explained by the use of a relatively sensitive avidin-biotin detection system.
Immunoreactions against total amelogenins were compared to immunoreactions against recombinant exon 4-encoded peptides. Exon 4 is the coding region for a 14-amino-acid segment that is part of M194 amelogenin and is not found in other mouse amelogenins. Unexpectedly, staining patterns between exon 4-encoded peptides and total amelogenins were very similar. These similarities in immunostaining between exon 4-encoded and total amelogenins were also observed in other areas of the tooth organ and at other developmental stages (data not shown). Therefore, we suggest that the entire developing enamel matrix contains both M194- and M180-encoded amelogenins.
We observed developmental changes in M179 recombinant amelogenin staining patterns. Vesicles within the ameloblasts were heavily stained only at embryonal stage E19. These well-contrasted spheres represent secretory vesicles during early enamel secretion. Whereas strong signals for amelogenin were detected in the enamel layer from 5 to 7 postnatal days, at later stages (9–13 postnatal days) amelogenin reaction products were accumulated at the interface between ameloblasts and enamel. This may indicate the presence of amelogenin in the most recently secreted enamel and removal of amelogenin from the remaining matrix, as suggested by Deutsch (1989).
In comparison to the amelogenins which reacted within the entire enamel layer or in the most recent enamel, tuftelin was localized in the developing enamel matrix as a fine line parallel to the dentine–enamel junction. Previous studies suggested that the first product secreted by ameloblasts is enriched with enamelins (Farge et al. 1987; Deutsch et al. 1984; Robinson et al. 1977). According to this suggestion, enamelins would be expected at the interface between dentine and enamel, as we report in the present study. In this location, enamelin could act as a nucleator for enamel crystal growth as hypothesized in several publications (Deutsch 1989; Deutsch et al. 1984).
Interestingly, our study indicated that the position of tuftelin signals changed from the odontoblast process towards the dentine–enamel junction during first mandibular mouse molar development. Whereas at embryonal stage E19 and in 1-day postnatal molars tuftelin was localized in the mesenchymal component of tooth biomineralization, from Day 3 onwards tuftelin was located in the enamel layer as a fine line at the interface between dentine and enamel. We speculate that tuftelin may have been originally produced in odontoblasts and then secreted via odontoblast processes into the newly formed enamel matrix. Whereas Inai et al. (1991) and Nakamura et al. (1994) suggested amelogenin translocation from the inner enamel epithelium to odontoblasts, we speculate that tuftelin may be secreted from the odontoblasts into the newly formed enamel protein matrix to form an admixture with amelogenins to support enamel crystal nucleation. This mechanism would resemble protein secretion into the developing teleost fish enameloid matrix via odontoblast processes, as documented via autoradiography (Shellis and Miles 1974). Another possibility is that tuftelin was originally secreted by ameloblasts and was subsequently translocated into the mesenchymal portion of the tooth.
Immunohistochemistry of mineralized tissues poses several technical challenges. Undecalcified frozen sections provide traps for antibodies on the rough mineral surface. Immunfluorescent images are difficult to interpret because of the autofluorescence observed in mineralized tissues. Therefore, we chose to use paraffin sections and immunoperoxidase techniques. In preliminary studies we observed that antigen preservation also depended on the type of fixation and decalcification. Throughout the present study these parameters were kept consistent. Our studies further indicated that the detection of signals depended on the concentration of the primary antibody and on the type of the detection system. The Zymed avidin–biotin staining kit applied in the present study featured well-contrasted slides with low background if an optimal primary antibody concentration was selected. Different detection kits or concentrations of the primary antibody resulted in slightly different data, especially regarding staining of the ameloblast cell bodies. From a technical standpoint, it was important to keep experimental procedures as comparable and consistent as possible. Further studies using immunogold electron microscopy (Nanci et al. 1985) may provide insights into crystal–protein relationships of amelogenins and tuftelin.
In summary, we used immunohistochemistry as a technique to compare the spatial distribution between amelogenin and tuftelin epitopes in the developing enamel matrix. Our studies revealed similarities in the spatial distribution of all amelogenin antigens investigated, significant differences between the distribution of amelogenin and tuftelin, and developmental changes in the distribution of amelogenin and tuftelin. The spatial differences in the distribution of amelogenin and tuftelin, as documented in this study, may be intepreted as an indication of functional differences between the two proteins during early enamel biomineralization.
Footnotes
Acknowledgements
Supported in part by intramural research funds from Baylor College of Dentistry (F9404) and NIH Traineeship DE07270-01.