
Abstract
Studies of chick–quail chimeras have reported that avian ultimobranchial C cells originate from the neural crest. It has consequently been assumed, without much supporting evidence, that mammalian thyroid C cells also originate from the neural crest. To test this notion, we employed both Connexin43-lacZ and Wnt1-Cre/R26R transgenic mice, because their neural crest cells can be marked. We also examined the immunohistochemical expression of a number of markers that identify migratory or postmigratory neural crest cells, namely, TuJ1, neurofilament 160, nestin, P75NTR, and Sox10. Moreover, we examined the expression of E-cadherin, an epithelial cell marker. At embryonic day (E)10.5, the neural crest cells densely populated the pharyngeal arches but were not distributed in the pharyngeal pouches, including the fourth pouch. At E11.5, the ultimobranchial rudiment formed from the fourth pouch and was located close to the fourth arch artery. At E13.0, this organ came into contact with the thyroid lobe, and at E13.5, it fused with this lobe. However, the ultimobranchial body was not colonized by neural crest–derived cells at any of these developmental stages. Instead, all ultimobranchial cells, as well as the epithelium of the fourth pharyngeal pouch, were intensely immunoreactive for E-cadherin. Furthermore, confocal microscopy of newborn mouse thyroid glands revealed colocalization of calcitonin and E-cadherin in the C cells. The cells, however, were not marked in the Wnt-Cre/R26R mice. These results indicated that murine thyroid C cells are derived from the endodermal epithelial cells of the fourth pharyngeal pouch and do not originate from neural crest cells.
Keywords
C
Neural crest cells are multipotent progenitor cells that are derived from the dorsal aspect of the neural tube. The cells migrate extensively during early stages of embryogenesis and give rise to various cell types, i.e., most of the peripheral nervous system, the skin melanocytes, and the ectomesenchymal derivatives such as smooth muscle cells, and bone and cartilage in the head and neck (Le Douarin and Kalcheim 1999). The migration and differentiation of neural crest cells have been extensively documented in avian species. Recently, mice have also been subjected to fate-mapping techniques using neural crest–specific transgenes (Waldo et al. 1999; Jiang et al. 2000). One of these is Connexin (Cx)43, a member of the family of proteins that make up gap junctions. Cx43 is highly expressed by neural crest cells and their derivatives (Lo et al. 1997). Thus, to label neural crest–derived cells, transgenic mice in which the Cx43 promoter sequence is linked to an lacZ reporter were constructed (Lo et al. 1997). Analysis of these mice has revealed that the lacZ expression pattern resembles the distribution of neural crest cells in chick–quail chimeras (Waldo et al. 1999). Moreover, we demonstrated that derivatives of the neural crest cells in Cx43-lacZ transgenic mice can be readily identified by immunohistochemistry using a monoclonal anti-β-galactosidase antibody (Kameda et al. 2004; Chisaka and Kameda 2005). This technique is thus useful for analyzing the neural crest cell lineages of mammals. In addition, Wnt1-Cre/R26R mice (double transgenic) are available to define the long-term fate of the neural crest lineages. The progeny of the neural crest cells are marked indelibly in the Wnt1-Cre/R26R mice (Chai et al. 2000; Jiang et al. 2000).
In the mouse, the ultimobranchial primordium buds off from the fourth pharyngeal pouch at embryonic day (E)11.5, moves close to the fourth arch artery during E11.5–12.5, and then moves to the final position of the thyroid gland at E13.0 (Kameda et al. 2007). Thus, active migration is required for the development of the murine ultimobranchial body. Subsequently, the ultimobranchial cells begin to disperse as C cells in the thyroid parenchyma at E14.5. Notably, at E14.5, the C cells begin to express neuronal markers, i.e., TuJ1, CGRP, and somatostatin. Moreover, we have demonstrated that Mash1 is expressed in the ultimobranchial cells of the wild-type murine embryos at E12.5–E13.5, and that Mash1-null mutant mice lack thyroid C cells (Kameda et al. 2007). Mash1 is a basic helix-loop-helix (bHLH) transcription factor that is a mammalian homolog of the Drosophila achaete-scute complex (asc) gene and that plays a key role in the differentiation of autonomic neurons from uncommitted neural crest cells (Johnson et al. 1990; Guillemot et al. 1993). Thus, Mash1 is essential for murine C-cell development. At face value, these latter observations could support the notion that C cells originate from neural crest cells. However, our observations in the present report suggest instead that Mash1 may actually be responsible for endowing the endoderm-derived C cells in the mouse with neuronal properties.
Cadherins are a family of transmembrane glycoproteins that are responsible for Ca2+-dependent, cell–cell adhesion (Takeichi 1988). E-cadherin is distributed mainly in epithelia of both ectodermal and endodermal origin. It plays a role in the establishment of cell polarity and maintenance of the epithelial phenotype, and is essential for the normal development of epithelia (Vleminckx and Kemler 1999). Thyroid follicular cells, which are derived from the endodermal epithelium of the ventral pharyngeal floor, express high levels of E-cadherin at all developmental stages (Fagman et al. 2003). Similarly, all epithelial cells of the pancreatic buds and gut, i.e., endoderm derivatives, express E-cadherin at early stages of development and onwards (Esni et al. 2001). Furthermore, E-cadherin is required for the formation and differentiation of mammary alveolar epithelium (Delmas et al. 1999). Whether the ultimobranchial body expresses E-cadherin during its organogenesis has not yet been tested.
In both mouse and chick embryos, the cardiac neural crest, a subpopulation of the cranial neural crest, populates the caudal three pharyngeal arches, i.e., the third, fourth, and sixth arches (Waldo et al. 1999; Jiang et al. 2000). To clarify the origin of mammalian thyroid C cells, we here examined the fourth pharyngeal pouch and ultimobranchial body of Cx43-lacZ transgenic mice, in which the ectomesenchymal neural crest cells colonizing the pharyngeal arches can be identified. We also examined the ultimobranchial body and thyroid gland of Wnt1-Cre/R26R transgenic mice, in which derivatives of the neural crest cells are marked indelibly. Furthermore, we subjected early mouse embryos to immunohistochemical analysis using various antibodies that label migratory or postmigratory neural crest cells. These antibodies include TuJ1, which recognizes neuronal progenitors (Kameda 1995); neurofilament (NF)160, a marker of neuronal differentiation in the ontogeny of the sympathoadrenal lineage (Sommer et al. 1995); nestin, a marker of neural stem cells and uncommitted neural crest cells (Rauch et al. 2006); and P75NTR and Sox10, which are both markers of differentiating neural crest cells and glial cells (Kim et al. 2003; Young et al. 2003). In addition, to elucidate whether C cells bear characteristics of endodermal epithelium, we examined the expression pattern of the epithelial marker E-cadherin in the fourth pharyngeal pouch, ultimobranchial body, and thyroid C cells. The results indicate that E-cadherin is expressed at continuously high levels throughout the entire development of murine C cells. In contrast, neither the ultimobranchial body nor the fourth pharyngeal pouch in mouse embryos is colonized by ectomesenchymal neural crest cells or neuronal progenitors derived from ganglia. Moreover, the thyroid C cells are not marked in the Wnt1-Cre/R26R mice.
Immunohistochemical analysis of β-galactosidase expression by the neural crest derivatives in an embryonic day (E)10.5 Connexin (Cx)43-lacZ mouse using the β-galactosidase (Gal) monoclonal antibody (
Materials and Methods
Animals
Cx43-lacZ transgenic mice were kindly provided by Dr. Cecilia W. Lo. In these mice, a 6.5-kb promoter fragment from the mouse Cx43 gene is used to drive lacZ expression in neural crest cells (Lo et al. 1997). Thus, Cx43-lacZ mice are a stable transgenic line that expresses β-galactosidase in the nuclei of neural crest cells (Waldo et al. 1999). Transgenic males were mated with outbred females of mixed background (CD-1, C57Bl/6, 129Sv) to obtain embryos at various stages of development. Wnt1-Cre/R26R transgenic mice were constructed as described previously (Chai et al. 2000; Jiang et al. 2000). In addition, embryonic and newborn CD-1 mice from an inbred colony were used for histological and immunohistochemical studies. Noon on the day on which a copulation plug was found was designated E0.5.
Cx43-lacZ and Wnt1-Cre/R26R transgenic mice were fixed in 4% paraformaldehyde (PFA) in 0.1 M phosphate buffer (PB), pH 7.4, for 24 hr. Specimens were rinsed in PB containing 10% sucrose for 2–4 hr and then immersed overnight at 4C in PB containing 25% sucrose. Serial sections (6-μm-thick) were cut with a cryostat.
CD-1 mouse embryos were fixed in Bouin's solution or 8% PFA in PB for 24 hr, embedded in paraffin, and then serially sectioned along the frontal, sagittal, or cross planes at a thickness of 5 μm. Selected sections were stained with hematoxylineosin to help determine the morphological orientation.
Immunohistochemistry and X-galactosidase Histochemistry
Immunohistochemical staining was carried out by using the streptavidin-biotin-peroxidase or peroxidase-antiperoxidase method as described previously (Kameda 1995). The following primary antibodies were employed: the monoclonal antibody to β-galactosidase (Promega, Madison, WI; 1:300 dilution); the monoclonal antibody TuJ1 that recognizes the neuron-specific class III β-tubulin isotype, cβ4 (Berkeley Antibody Company, Richmond, CA; 1:500 dilution); the monoclonal antibody to NF160 (clone NN18, Sigma, St. Louis, MO; 1:200 dilution); the monoclonal antibody to E-cadherin (clone 36, BD Biosciences, San Diego, CA; 1:100 dilution); and the polyclonal antisera to PGP9.5 (Dako, Carpinteria, CA; 1:200 dilution), human nestin (Immuno-Biological Lab., Gunma, Japan; 1:20 dilution), P75NTR (Promega; 1:100 dilution), Sox10 (Chemicon, ElSegundo, CA; 1:100 dilution), calcitonin (Dako; prediluted), CGRP (Milab, Malmö, Sweden; 1:1000 dilution), somatostatin (produced in our laboratory as described in Kameda et al. 1982; 1:1000 dilution), bovine tyrosine hydroxylase (Chemicon; 1:500 dilution), and met-enkephalin (UCB-Bioproducts, Brussels, Belgium; 1:1000 dilution).
To examine the overlap between calcitonin and E-cadherin immunoreactivity, the thyroid glands of newborn mice were stained for immunofluorescence and viewed on a Bio-Rad MRC 2000 confocal laser scanning microscope. The secondary antibodies used were rhodamine-conjugated swine anti-rabbit IgG antiserum (Dako; 1:30 dilution) or fluorescein isothiocyanate–conjugated goat anti-mouse IgG antiserum (Chemicon; 1:50 dilution). All incubation steps were performed at 4C. Single optical sections of calcitonin and E-cadherin immunostaining were captured separately. The confocal microscope images were then merged by using LaserSharp 2000 image software.
X-galactosidase (X-gal) staining was carried out according to standard procedures as described previously (Chai et al. 2000; Chisaka and Kameda 2005).
Results
Development of the Murine Ultimobranchial Body and Thyroid C Cells and Their Expression of Neural Crest–derived Cell Markers
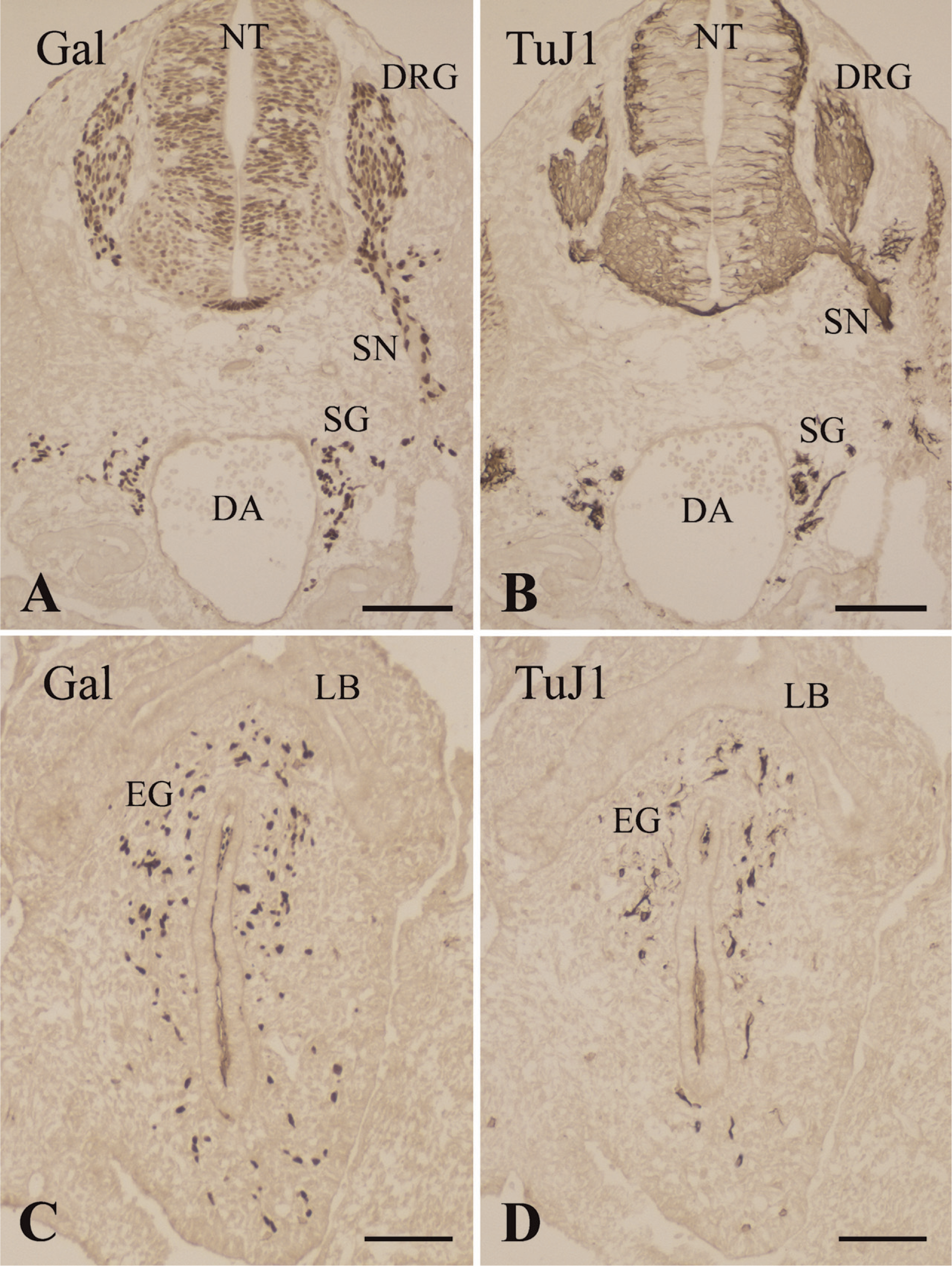
At E10.5, β-galactosidase expression in the Cx43-lacZ transgenic mice was detected in the neural tube and neural crest derivatives, including the dorsal root ganglia, peripheral nerves, sympathetic ganglia, and enteric ganglia (Figures 1A and 1C). At this stage, the β-galactosidase expression pattern highlighted a continuous stream of cell populations migrating from the neural crest. These neuronal neural crest–derived cells were immunoreactive with the TuJ1 monoclonal antibody (Figures 1B and 1D).
Consecutive frontal sections of the pharyngeal arches and pouches in an E10.5 mouse immunostained with the protein gene product (PGP)9.5 (
At this point (E10.5), it was possible to precisely identify the fourth pharyngeal pouch for the first time. The pharyngeal pouch epithelium, which consists of high columnar cells, was immunoreactive for PGP9.5 (Figure 2A). Invasion of neuronal cells and nerve fibers immunoreactive for TuJ1, NF160, or nestin into the epithelia of pharyngeal pouches, including the fourth pouch, was not detected (Figures 2B–2D). The pharyngeal arches were colonized by the ectomesenchymal neural crest cells, and the mesenchyme of the pharyngeal arches was filled with β-galactosidase–positive cells (Figure 3A). However, the epithelia of the pharyngeal pouches were free of such β-galactosidase–positive cells.
Immunohistochemical analysis of β-galactosidase expression by neural crest derivatives in Cx43-lacZ mice at E10.5 (
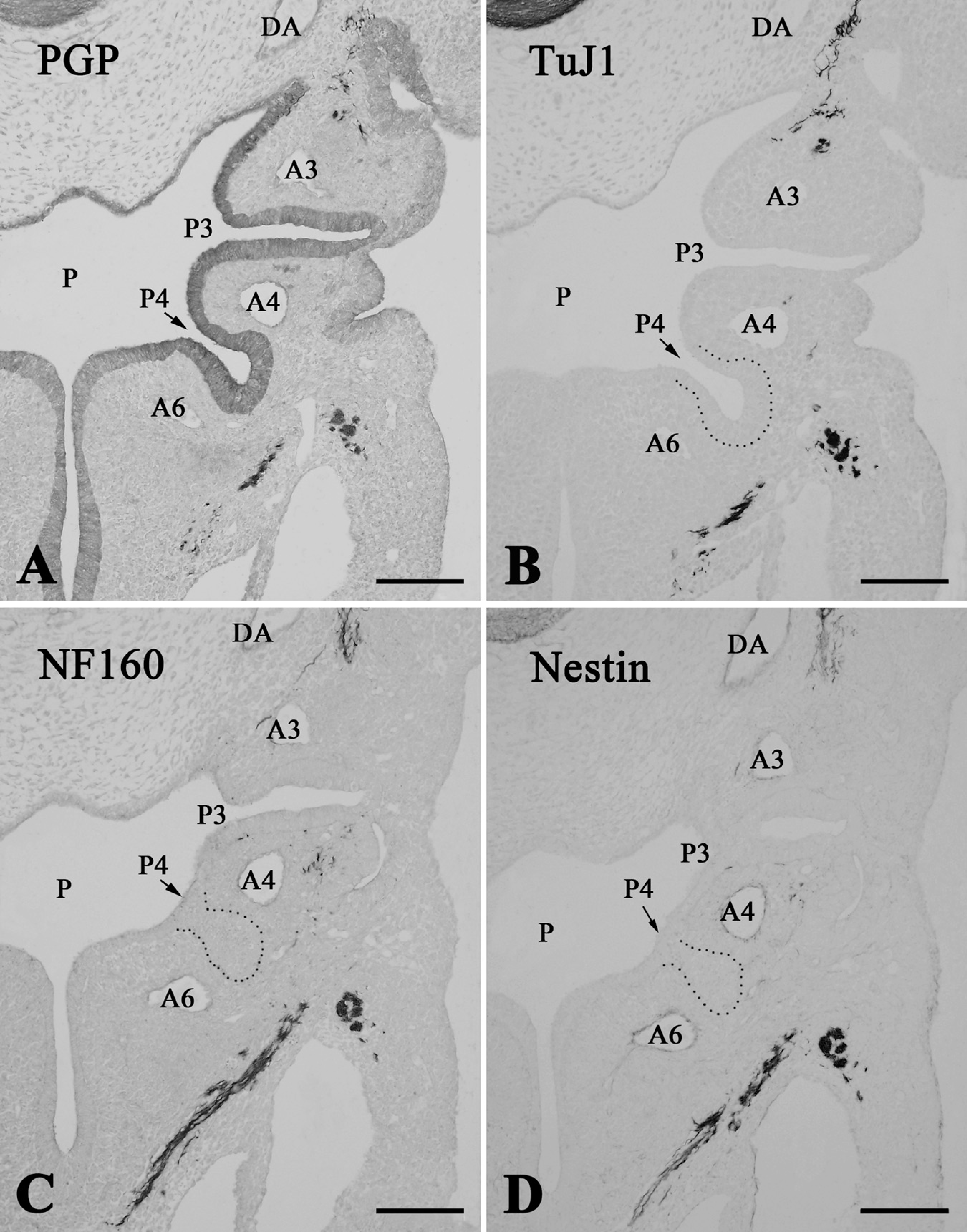
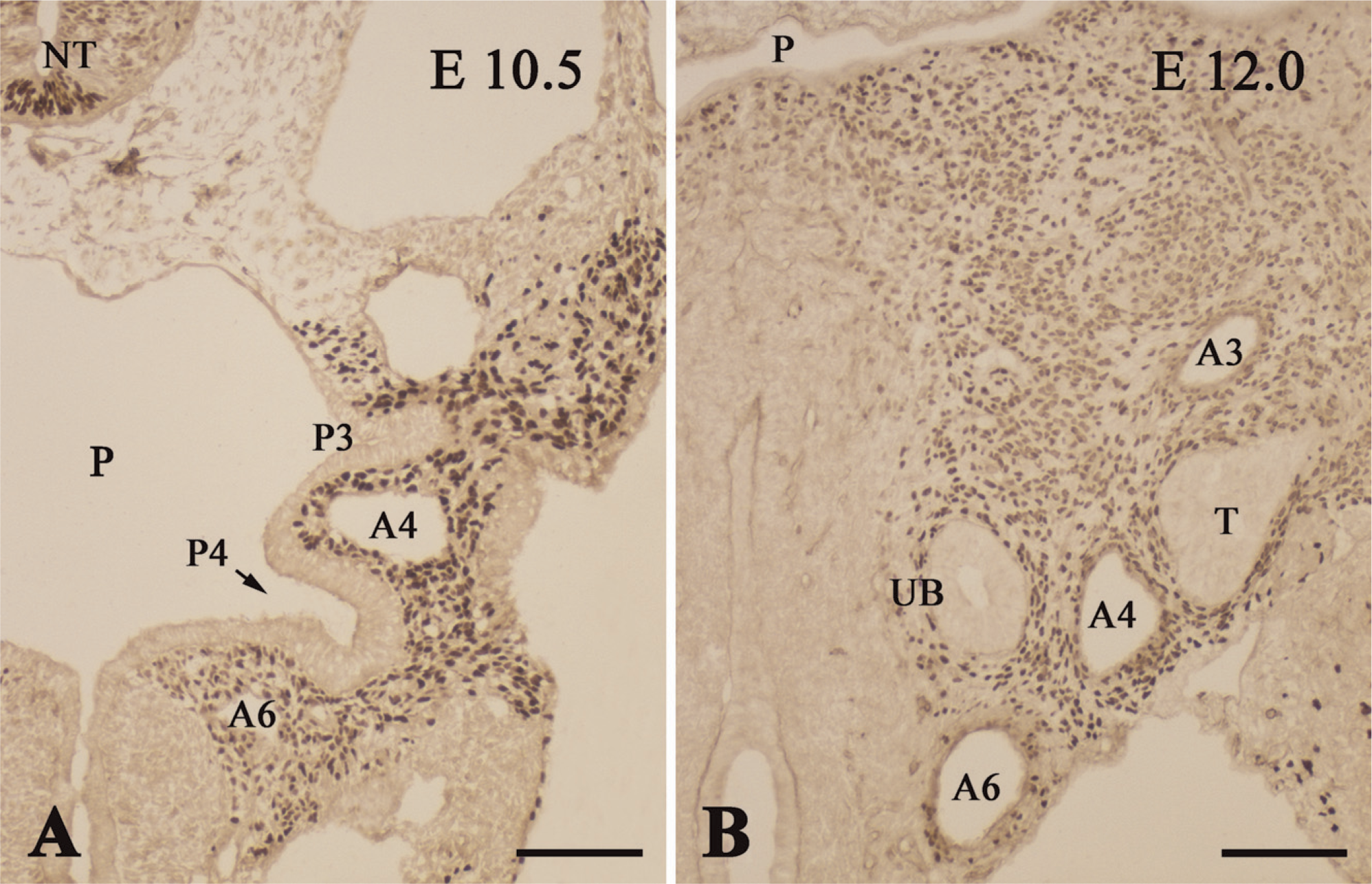
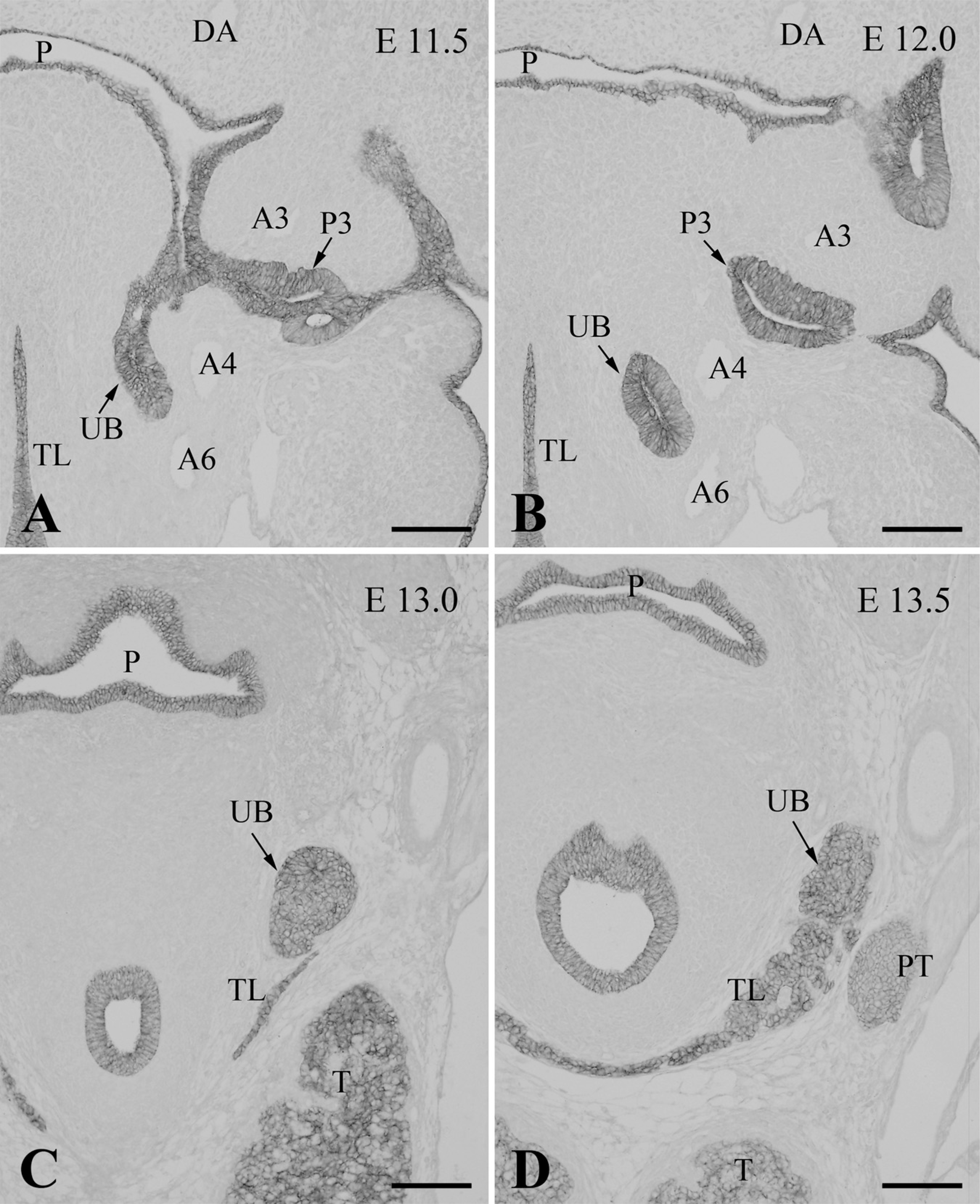
At E11.5, the ultimobranchial rudiment formed from the fourth pharyngeal pouch and became located medially, close to the fourth arch artery (Figure 4A). At this point, the organ consisted of an ovoid follicle with a central cavity that was lined with high columnar epithelium bearing mitotic cells. The ultimobranchial rudiment, along with other pharyngeal derivatives, including the thymus, parathyroid, and thyroid primordial, was immunoreactive for PGP9.5 (Figure 4A). Nerve fibers exhibited intense immunoreactivities for PGP9.5, TuJ1, NF160, and nestin (Figures 4A–4D), and showed moderate immunoreactivity for P75NTR (data not shown). Schwann cell nuclei in the nerve fibers were intensely immunoreactive for Sox10 (see Figure 2 in Kameda 2007). However, TuJ1-, NF160-, nestin-, P75NTR- and Sox10-immunoreactive cells were not detected in the ultimobranchial rudiment at E11.5– E12.5 (Figures 4B–4D; and data not shown).
In the pharyngeal region at E11.5–E12.5, β-galactosidase–immunoreactive cells were widely distributed in the distal vagal ganglion, superior cervical ganglion, vagus nerve, sympathetic nerve and muscles, and mesenchymal tissue. However, the ultimobranchial body, thymus, parathyroid, and thyroid primordia, which were all derived from the pharyngeal endoderm, lacked β-galactosidase–immunoreactive cells (Figure 3B). Thus, neither ectomesenchymal nor neuronal neural crest cells invaded the ultimobranchial rudiment at E11.5–E12.5.
Consecutive frontal sections of the pharyngeal region in E11.5 (
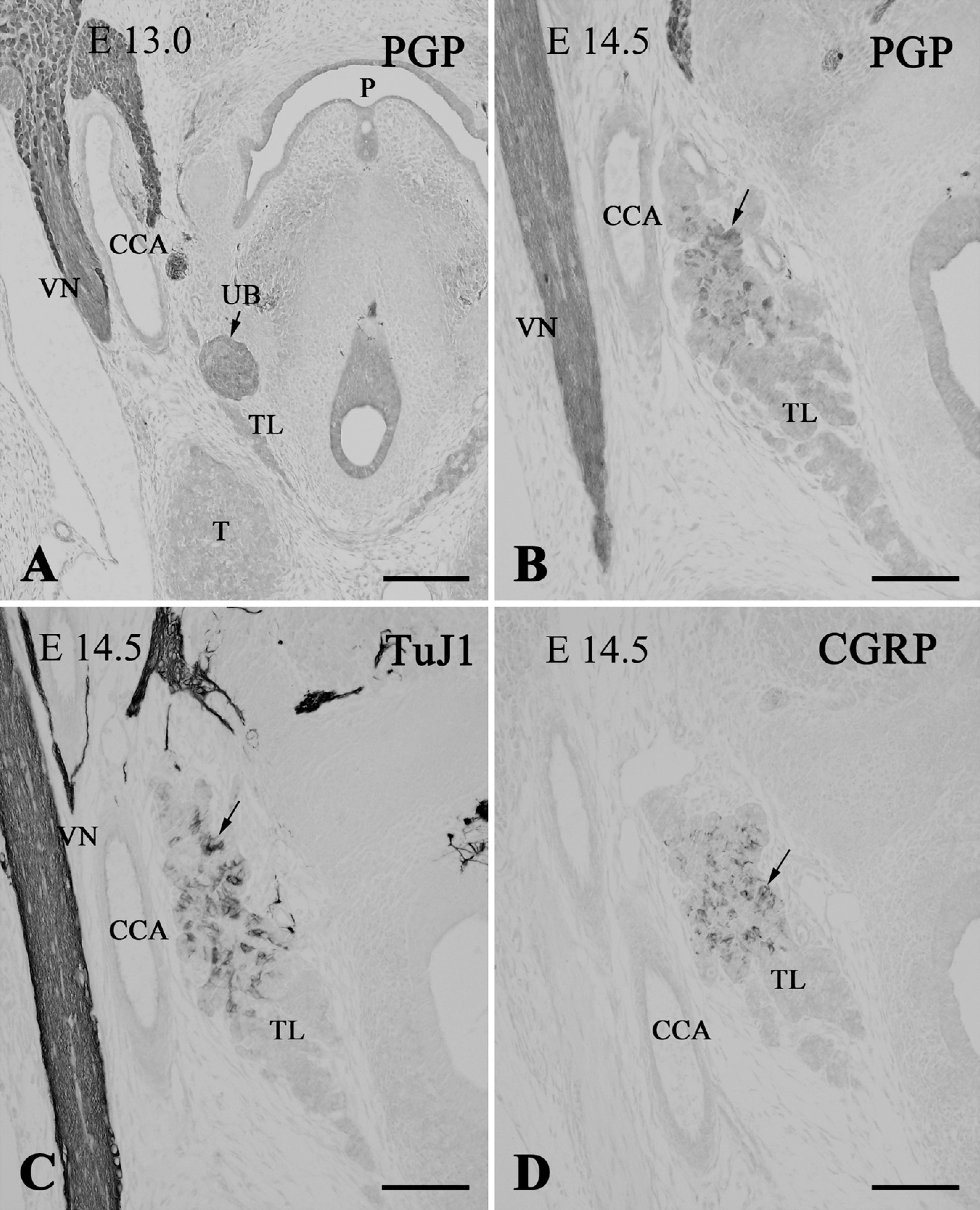
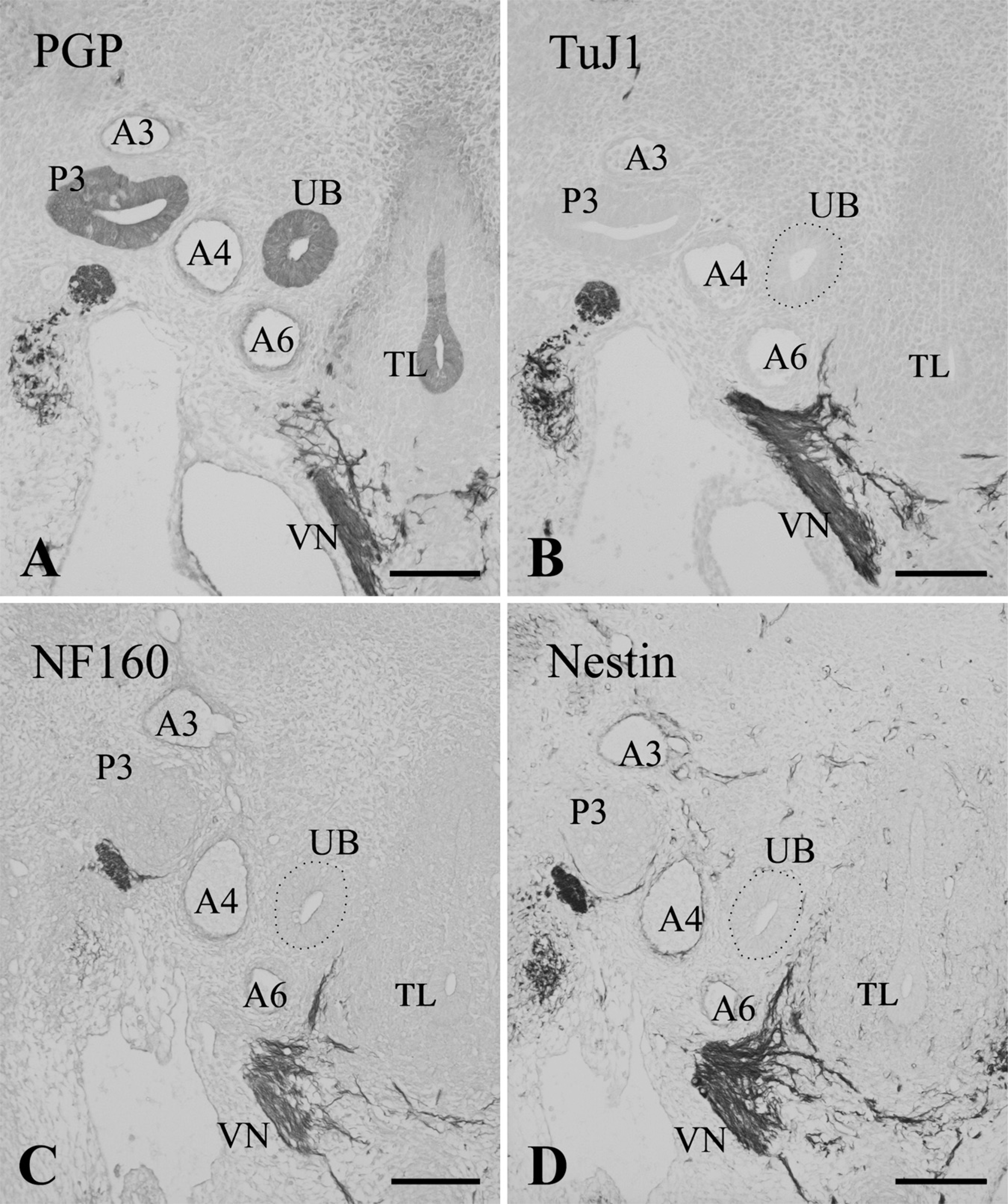
Immunostaining of the thyroid lobes (TL) in E13.0 (
At E13.0, the ultimobranchial body became a solid cluster of cells and joined the thyroid lobe (Figure 5A). The organ was still immunoreactive for PGP9.5. In E13.5 Wnt1-Cre/R26R mice, the ultimobranchial body, as well as thyroid and parathyroid glands, was free of X-gal–positive cells, although these organs were densely surrounded by the positive cells (Figures 6A and 6B). At E14.5, PGP9.5-immunoreactive cells began to scatter within the thyroid parenchyma as C cells (Figure 5B). At this stage, the C cells started expressing the neuronal markers TuJ1, CGRP, and somatostatin (Figures 5C and 5D; and data not shown). Calcitonin immunoreactivity appeared in the C cells at E15.5 (see Kameda et al. 2007). Immunoreactivity for enkephalin and tyrosine hydroxylase was not detected in the C cells of the mouse thyroid gland at any stage of development (data not shown).
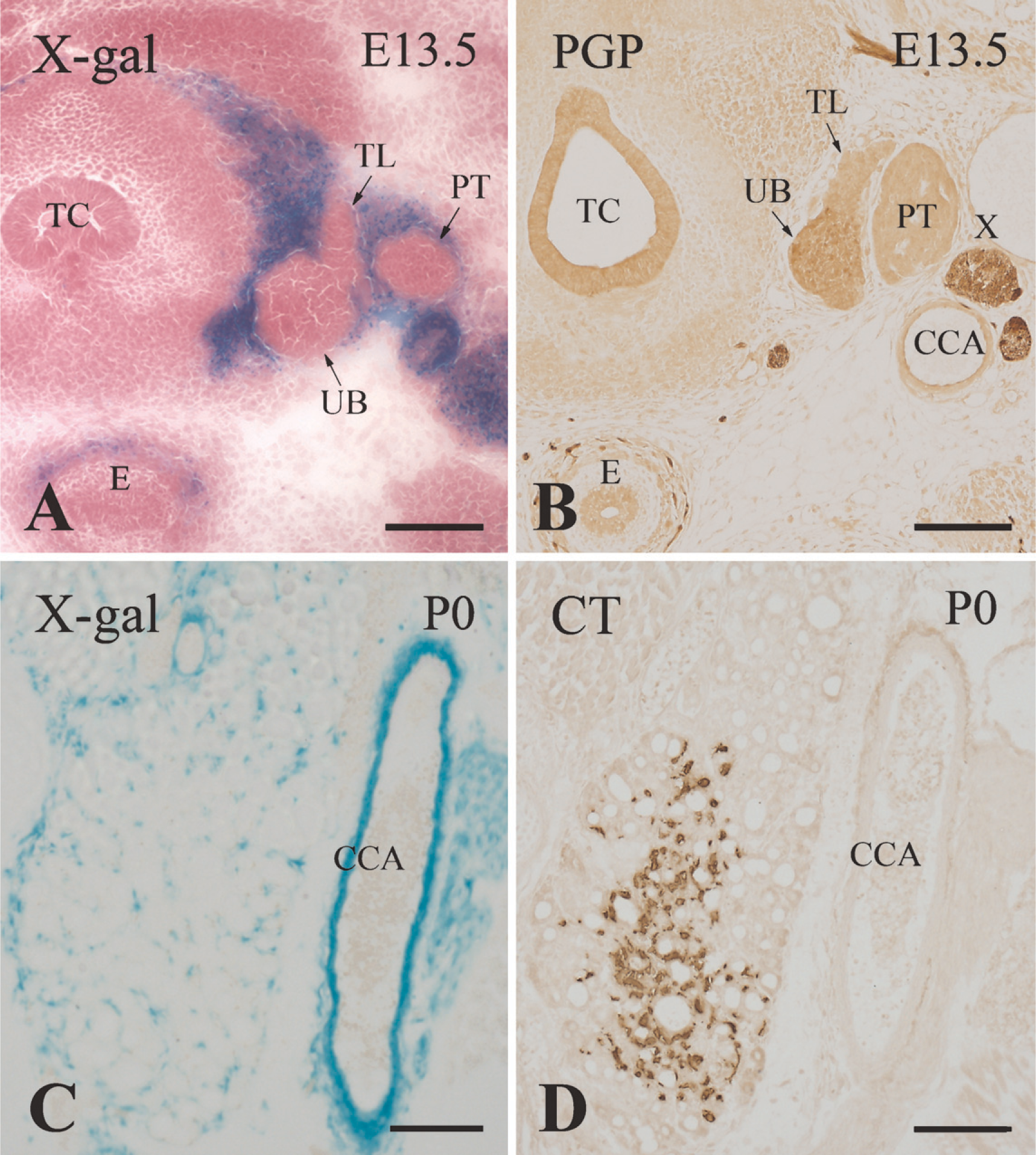
X-galactosidase (X-gal) staining of Wnt1-Cre/R26R transgenic mice. Transverse sections of the ultimobranchial bodies at E13.5 (
In newborn Wnt1-Cre/R26R mice, the wall of the common carotid artery and connective tissue distributed in the thyroid parenchyma were labeled by X-gal staining (Figure 6C). However, no X-gal labeling was localized on the C cells immunoreactive for calcitonin (Figures 6C and 6D).
Expression of E-Cadherin by the C Cells and Their Precursors
E-cadherin is a marker of epithelial cells, and E-cadherin immunoreactivity is known to localize mainly to the lateral cell membrane, which is the area of cell–cell contact. The endodermal lining of the pharyngeal pouches at E11.5–E12.0 was immunoreactive for E-cadherin (Figures 7A and 7B). Moreover, the ultimobranchial cells exhibited intense immunoreactivity for E-cadherin at all stages of development. Thus, the ultimobranchial primordium still attached to the pharyngeal endoderm at E11.5 (Figure 7A), the ultimobranchial follicle located close to the fourth arch artery at E12.0 (Figure 7B), and the ultimobranchial body forming a solid cluster of cells at E13.0 (Figure 7C) all strongly expressed E-cadherin. Moreover, when the ultimobranchial body entered and fused with the thyroid lobe at E13.5, the whole thyroid lobe, including the ultimobranchial body, was immunoreactive for E-cadherin (Figure 7D). Indeed, the thyroid follicular cells displayed immunoreactivity for E-cadherin throughout thyroid organogenesis.
To clarify the relationship between calcitonin and E-cadherin in the developing C cells, confocal microscopic analysis was performed (Figures 8A–8C). In the newborn mouse, the same C cells were immuoreactive for both calcitonin and E-cadherin (Figure 8C). However, although E-cadherin was primarily localized on the C-cell membrane, calcitonin immunoreactivity was distributed in the C-cell cytoplasm (Figures 8A and 8B).
Discussion
The Cx43-lacZ transgenic mouse is a good model for studying the migration and development of neural crest cells (Lo et al. 1997; Waldo et al. 1999). This mouse allows us to trace the movements of neural crest–derived cells through successive stages during the formation and development of the ultimobranchial rudiment at E10.5–E12.5. Thus, to clarify the origin of mammalian thyroid C cells, we examined the movement of neural crest–derived cells in the pharyngeal region of Cx43-lacZ embryos by immunohistochemical detection of their β-galactosidase expression. Furthermore, we employed Wnt1-Cre/R16R double-transgenic mice to identify the derivatives of neural crest cells. In this double-transgenic mouse, all neural crest cell progeny are efficiently and stably marked, even in the postnatal animals (Chai et al. 2000; Jiang et al. 2000). In addition, because many markers that identify developing neural crest–derived cells in mouse embryos have been discovered to date (Sommer et al. 1995; Kim et al. 2003; Young et al. 2003; Rauch et al. 2006), we also analyzed mouse embryo immunoreactivity with the TuJ1, NF160, nestin, P75NTR, and Sox10 antibodies.
In the Cx43-lacZ embryos, the ectomesenchymal neural crest cells that were distributed in the pharyngeal arches were intensely immunoreactive for β-galactosidase. However, these β-galactosidase–immunoreactive cells never entered the fourth pharyngeal pouch or the ultimobranchial rudiment at E10.5–E12.5. In addition, cells expressing the neural crest markers TuJ1, NF160, nestin, P75NTR, and Sox10 were never detected in either tissue. In the Wnt1-Cre/R16R mice at E13.5 and onwards, neither the ultimobranchial body nor thyroid C cells were labeled by X-gal staining. Thus, the mouse ultimobranchial body does not contain neural crest–derived cells at any stage of its development. Moreover, the thyroid C cells did not express the neural crest marker.
In contrast, the ultimobranchial cells exhibited intense immunoreactivity for E-cadherin, an epithelial marker, at all developmental stages, namely, at E11.5–E12.5, when the ultimobranchial rudiment that had formed from the fourth pharyngeal pouch had a follicle structure and was lined by high columnar cells; at E13.0, when the organ became a solid cluster of cells and had connected with the thyroid lobe; and at E14.5, when the ultimobranchial body began to disperse as C-cell groups within thyroid parenchyma. Moreover, confocal microscopic analysis of neonatal mice revealed that C cells not only express calcitonin, which is a reliable marker of C cells (Kameda et al. 2007), they also express E-cadherin. It has been shown that loss of the epithelial phenotype involves downregulation of E-cadherin (Savagner 2001). Thus, the ultimobranchial body maintains its original epithelial phenotype and E-cadherin–mediated homotypic cell–cell adhesion during all steps of its formation, differentiation, and migration. These observations together indicate that the endodermally derived ultimobranchial body differentiates into C cells and that mouse thyroid C cells originate from the endoderm.
Immunohistochemical analysis of E-cadherin expression in the ultimobranchial body by using the E-cadherin antibody. At E11.5, the ultimobranchial rudiment was attached to the pharynx (
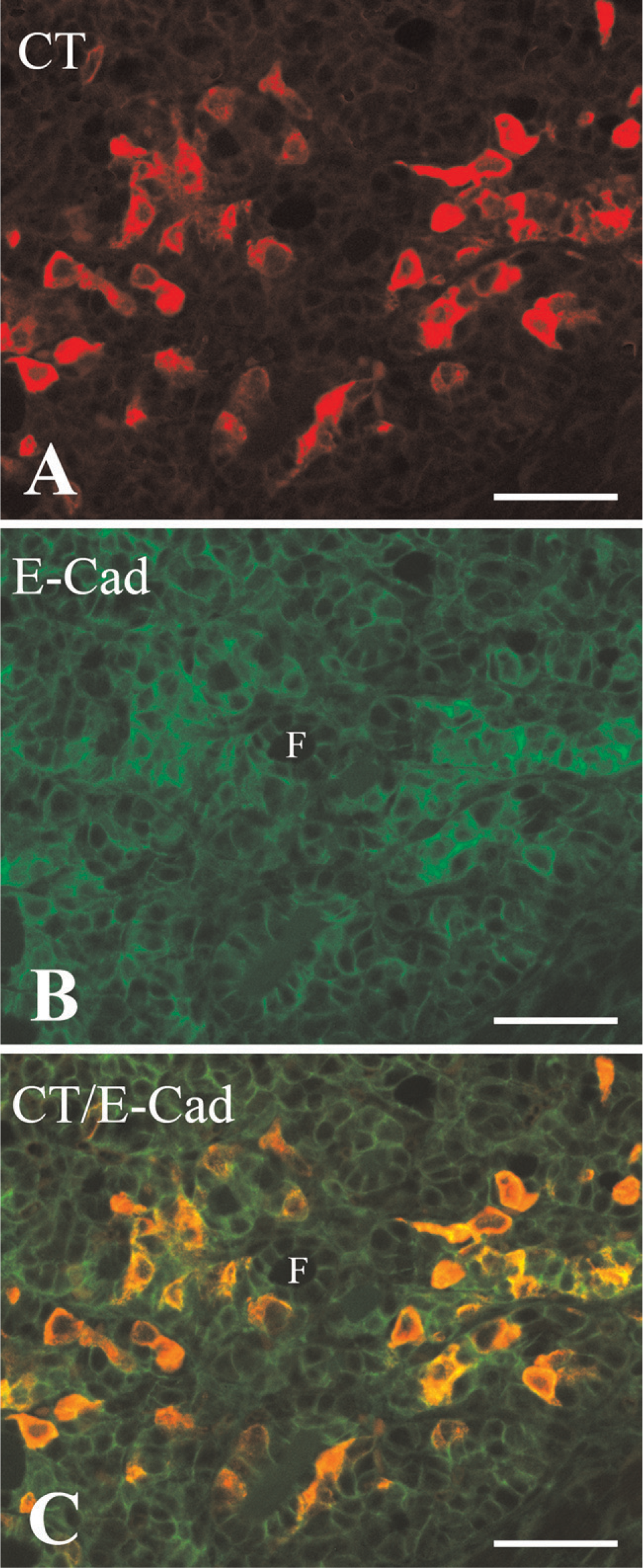
Confocal microscopy of the newborn mouse thyroid gland after double-immunofluorescence labeling for calcitonin (CT) and E-cadherin (E-Cad). Immunostaining for calcitonin (
Pancreatic endocrine cells and pulmonary neuroendocrine cells share several characteristics with neurons; this led initially to the idea that they may originate from the neuroectoderm (see Le Douarin 1988; Van Lommel et al. 1999, for reviews). However, it has now been concluded that these cells are actually derived from the endoderm. Notably, the endocrine differentiation in the developing pancreas requires the bHLH transcription factors neurogenin3 and neuroD1, which generally play roles in neuronal differentiation (Wilson et al. 2003). Furthermore, pulmonary neuroendocrine cells express Mash1, which plays a key role in the differentiation of autonomic neurons from uncommitted neural crest cells (Johnson et al. 1990; Guillemot et al. 1993). Indeed, the formation of pulmonary neuroendocrine cells is impaired when Mash1 is defective (Borges et al. 1997). These observations together suggest that transcription factors that are generally expressed in nervous tissue also endow endocrine cells with neuronal cell traits. Interestingly, we showed previously that the mouse ultimobranchial body begins to express Mash1 at E11.5, and that this expression becomes intense at E12.5, when the organ moves close to the aortic arch or the base of the common carotid artery (Kameda et al. 2007). This Mash1 expression pattern then weakens markedly at E14.5, which is when the ultimobranchial body has dispersed within the thyroid lobe. At this stage, neuronal markers such as TuJ1, CGRP, and somatostatin begin to appear in the C cells. These observations suggest that Mash1 may also promote C-cell development and that it is responsible for the neuronal properties of these cells. Thus, as with pancreatic endocrine cells and pulmonary neuroendocrine cells, the neuronal features of C cells may be driven by the expression of factors whose primary roles are in neuronal differentiation.
Our previous study of the chick ultimobranchial rudiment revealed that it is penetrated by TuJ1- and HNK-1-immunoreactive neuronal cell streams that originate from the distal vagal ganglion and continue to migrate around the esophagus (Kameda 1995). In mouse embryos, however, neither the TuJ1-immunoreactive cells nor the nerve fibers invaded the ultimobranchial rudiment. Notably, the mouse and chick ultimobranchial rudiments localize differently. In chick embryos, the rudiment is located close to the esophagus, which is surrounded by the enteric ganglia, whereas in mouse embryos, the rudiment is located in the pharyngeal region. Thus, the murine ultimobranchial body is not in direct contact with ganglia or nerve bundles. In addition, the ultimobranchial C cells in chick embryos are immunoreactive for enkephalin and tyrosine hydroxylase (Kameda 1991), whereas the mouse thyroid C cells are not immunoreactive for these substances. Tyrosine hydroxylase, which is a rate-limiting enzyme for catecholamine synthesis, is expressed by some neural crest–derived cells (Fauquet and Ziller 1989). Instead, in the mouse, the cells that express tyrosine hydroxylase and enkephalins are the adrenal chromaffin cells, which are derived from the neural crest cells (Bornstein et al. 2000; Huber et al. 2002).
It has been reported that the neural crest cells in avian species contribute connective tissue to pharyngeal derivatives such as the thymus, parathyroid, and thyroid glands, as well to the ultimobranchial gland (Le Lièvre and Le Douarin 1975). Indeed, the ablation of the premigratory cardiac neural crest cells in chick embryos results in hypoplastic deficiencies in the thymus, parathyroid, and thyroid glands (Bockman and Kirby 1984). It has been suggested that the development of these organs may depend on direct interactions of mesenchymal derivatives of the neural crest with the pharyngeal epithelium. As far as we know, the effect of ablation of the neural crest on the development of the avian ultimobranchial gland has not been investigated to date. It has been demonstrated in Wnt1-Cre/R26R transgenic mouse embryos that although the ectomesenchymal neural crest cells densely invest the thymus, few of these cells actually enter the organ during its migration (Jiang et al. 2000). The adult thymus, parathyroid, thyroid, and ultimobranchial glands are located well away from the sites at which they started to develop, and their morphogenic processes involve profound cell migration. It is quite possible that the X-gal–positive neural crest cells that surround these organs during their migration may aid this migratory process in some way. In addition, in the mouse, as well as avian species, ectomesenchymal neural crest cells may contribute connective tissue to the pharyngeal derivatives, inasmuch as X-gal–positive connective tissues were detected in the thyroid gland of the newborn Wnt1-Cre/R26R mice.