
Abstract
Mice with a targeted deletion of the Hoxa3 gene have defects of derivatives of the third branchial arch and pouch. To address the role of the Hoxa3 gene in parathyroid organogenesis, we examined the third pharyngeal pouch development by immunohistochemistry (IHC) using the secretory protein (SP)-1/chromogranin A antiserum, which recognizes the parathyroid from its initial formation onward. At embryonic day (E) 11.5, the SP-1/chromogranin A-immunoreactive primary rudiment of the parathyroid appeared in the cranial region of the third pharyngeal pouch of wild-type embryos. In Hoxa3-null mutants, the third pharyngeal pouch was normally formed but failed to differentiate into the parathyroid rudiment, showing no immmunoreactivity for SP-1/chromogranin A. Classic studies using chick-quail chimeras have demonstrated that the ectomesenchymal neural crest cells are required for proper development of the pharyngeal pouch-derived organs, including the thymus and parathyroid glands. To visualize the migration and development of mesenchymal neural crest cells in Hoxa3 mutants, the heterozygotes were crossed with connexin43–lacZ transgenic mice in which β-galactosidase expression was specific to the neural crest cells. In Hoxa3 homozygotes and in wild types, ectomesenchymal neural crest cells densely populated the pharyngeal arches, including the third one, and surrounded the third pouch epithelium. These results indicate that lack of the Hoxa3 gene affects the intrinsic ability of the third pharyngeal pouch to form the parathyroid rudiment and has no detectable effect on the migration of neural crest cells.
Keywords
T
Chromogranin A, the major secretory protein of adrenal chromaffin cells, is a member of the granin/secretogranin family of acidic glycoproteins that participate in the storage and secretion of peptide hormones and are expressed in many endocrine and neuroendocrine cells (O'Connor et al. 1983; Fischer-Colbrie et al. 1987; Winkler and Fischer-Colbrie 1992). Secretory protein-1 (SP-1) coexists with parathormone (PTH) in secretory granules of the parathyroid chief cells (Cohn et al. 1984). SP-1 and chromogranin A are chemically similar if not identical proteins (Cohn et al. 1982). The present study shows that SP-1 immunore-activity appears in the parathyroid rudiment of the third pharyngeal pouch from the earliest stage of its organogenesis.
Hoxa3 belongs to the Hox family of transcription factors that play multiple roles in the segmental processes of anteroposterior patterning (Krumlauf 1994; Trainor and Krumlauf 2000). In the Hoxa3 homozygous null mutant mouse produced by gene targeting, the thymus and parathyroid glands derived from the third pharyngeal pouch are lacking (Chisaka and Capecchi 1991). The Hoxa3 homozygotes also show a deficiency of the common carotid artery originating from the third arch artery (Kameda et al. 2003). However, the initial formation of the third arch artery is not disturbed; the artery degenerates at embryonic day (E) 11.5 (Kameda et al. 2002). Therefore, the Hoxa3 gene is crucial for the development and differentiation of the third pharyngeal arch and pouch. It is unknown whether or not the initial formation of the parathyroid rudiment is affected by a lack of the Hoxa3 gene.
It has been demonstrated by studies of chick-quail chimeras that the mesenchymal components of thymus lobes and parathyroid glands of avian species are derived from the ectomesenchymal neural crest cells (Le Lièvre and Le Douarin 1975). Ablation of the premigratory cardiac neural crest in chick embryos results in disrupted development of the caudal pharyngeal arches, including thymus and parathyroid hypoplasia or agenesis (Kirby and Waldo 1995). Fate-mapping techniques using neural crest-specific trans-genes have recently become available for mouse embryos (Lo et al. 1997; Waldo et al. 1999; Jiang et al. 2000). In connexin (Cx) 43-lacZ transgenic mice in which the promoter sequence for Cx43 linked to a lacZ reporter, the derivatives of neural crest cells are specified by IHC demonstration of β-galactosidase expression.
The present study first clarified the organogenesis of parathyroid glands in wild-type mouse embryos using the SP-1/chromogranin A antiserum as a specific marker for parathyroid cells. Furthermore, we visualized in Cx43-lacZ transgenic mice whether or not mesenchymal neural crest cells enter the parathyroid and thymus primordia to control the growth of these organs. The main purpose of this study was to examine the etiology of the parathyroid deficiency in the Hoxa3-null mutants. Migrations of neural crest cells into the pharyngeal arches and apoptosis of the third pouch endoderm were estimated in Hoxa3 homozygous mutants in comparison with wild-types.
Materials and Methods
Animals
Targeted disruption of the Hoxa3 gene, production of chimeric mice, and determination of genotype by polymerase chain reaction have been reported previously (Chisaka and Capecchi 1991; Watari et al. 2001). The Cx43-lacZ transgenic mice were kindly provided by Dr. Cecilia W. Lo. A 6.5-kb promoter fragment from the mouse Cx43 gene has been used to drive lacZ expression in neural crest cells (Lo et al. 1997). Hoxa3 heterozygous mice were crossed with Cx43-lacZ transgenic mice. Noon on the day at which a copulation plug was found was designated as embryonic day 0.5 (E 0.5). Pregnant females were sacrificed on days 9.5, 10.5, 11.5, 12.0, 12.5, 13.0, 13.5, 14.5, 15.5, 16.5, or 18.5 after conception. Newborn mice were also used. At least four wild-types and null mutants were examined at each stage. In particular, 40 wild-types and 60 null mutants were analyzed at E 11.5, 15 wild-types and 20 null mutants at E 12.0, and 25 wild-types and 40 null mutants at E 12.5. All animal procedures were approved by the Animal Experimentation and Ethics Committee of Kitasato University School of Medicine or that of the Graduate School of Bio-studies, Kyoto University, and conformed to NIH guidelines. For histological study, the specimens were fixed in Bouin's solution or 8% paraformaldehyde (PFA) in phosphate buffer (PB) for 24–48 hr, embedded in paraffin, and then serially sectioned in the frontal or sagittal plane at a thickness of 5 μm. Selected sections were stained with hematoxylin-eosin to help determine morphological orientation.
TUNEL Assay
To visualize apoptotic nuclei, sections fixed in 4% or 8% PFA in PB were stained by the terminal transferase dUTP-biotin nick-end labeling (TUNEL) technique according to the manufacturer's instructions (ApopDETEK In Situ Cell Death Assay kit; DAKO, Carpinteria, CA). Specificity controls included the alternative omission of the biotin-dUTP, of the terminal deoxynucleotidyl transferase (TdT), or of the streptavidin-horseradish peroxidase conjugate. No labeling was detected in any control sections.
Immunohistochemistry
IHC staining was carried out by the streptavidin-biotin-per-oxidase method or the peroxidase-antiperoxidase (PAP) method as described previously (Kameda et al. 1998). The following primary antibodies were employed: the monoclonal anti-β-galactosidase antibody and the polyclonal anti-SP-1/chromogranin A and anti-human protein gene product (PGP) 9.5 antisera. The SP-1/chromogranin A antiserum was purchased from Incstar (Stillwater, MN) and used at a dilution of 1:500. The PGP 9.5 antiserum was purchased from UltraClone (Isle of Wight, UK) and used at a dilution of 1:600. The β-galactosidase antibody was purchased from Promega (Madison, WI) and used at a dilution of 1:300. Control reactions included replacing the primary antibodies with normal (non-immune) rabbit or mouse Ig and omission of the primary antibodies. In addition, for β-galactosidase immunoreactivity non-transgenic embryos were examined in parallel as controls. All control reactions were negative.
Results
To determine the etiology of late-term absence of the parathyroid gland in Hoxa3-null mutant mice, we examined the differentiation and development of the third pharyngeal pouch using the SP-1/chromogranin A antiserum as a specific marker for the parathyroid rudiment. The fusion of the endoderm of the third pharyngeal pouch and the ectoderm of the third cleft occurs by E 9.5 (Cordier and Haumont 1980). In addition to nerve cells and fibers, the endodermal epithelial cells of the pharyngeal pouches were immuno-reactive for PGP9.5. At E 10.5, the third pharyngeal pouch was fissure-like in shape and was lined with high columnar epithelial cells immunoreactive for PGP 9.5 (Figure 1A). SP-1/chromogranin A-immunoreactive cells did not yet appear in the third pharyngeal pouch of E 10.5 wild-type mice (Figure 1B). In the Hoxa3 homozygous mutants, the formation of the third pharyngeal pouch was normal until E 10.5 (Figure 1C). At this stage, the second and third pharyngeal clefts were situated closely in the wild-types, whereas they did not approach in the null mutants. Therefore, the shape of the third pharyngeal arch in the mutants differed somewhat from that of wild-types as early as E 10.5.
Consecutive frontal sections of the pharyngeal region in E 10.5 wild-type embryo (+/+) immunostained with PGP 9.5 (
At E 11.5, the third pharyngeal pouch was separated from the pharynx in many wild-type embryos, forming an elongated follicular structure (Figure 2A). A bulge of the primary parathyroid rudiment was detected in the cranial region of the third pharyngeal pouch. The rudiment was composed of multiple layers of cuboidal cells immunoreactive for SP-1/chromogranin A (Figure 2B). On the other hand, in Hoxa3 homozygotes at E 11.5, the third pharyngeal pouch was still attached to the pharynx and retained the fissurelike structure (Figure 2C). The third pouch endoderm consisted of high columnar cells and there was no aggregation of cuboidal cells immunoreactive for SP-1/chromogranin A (Figure 2D).
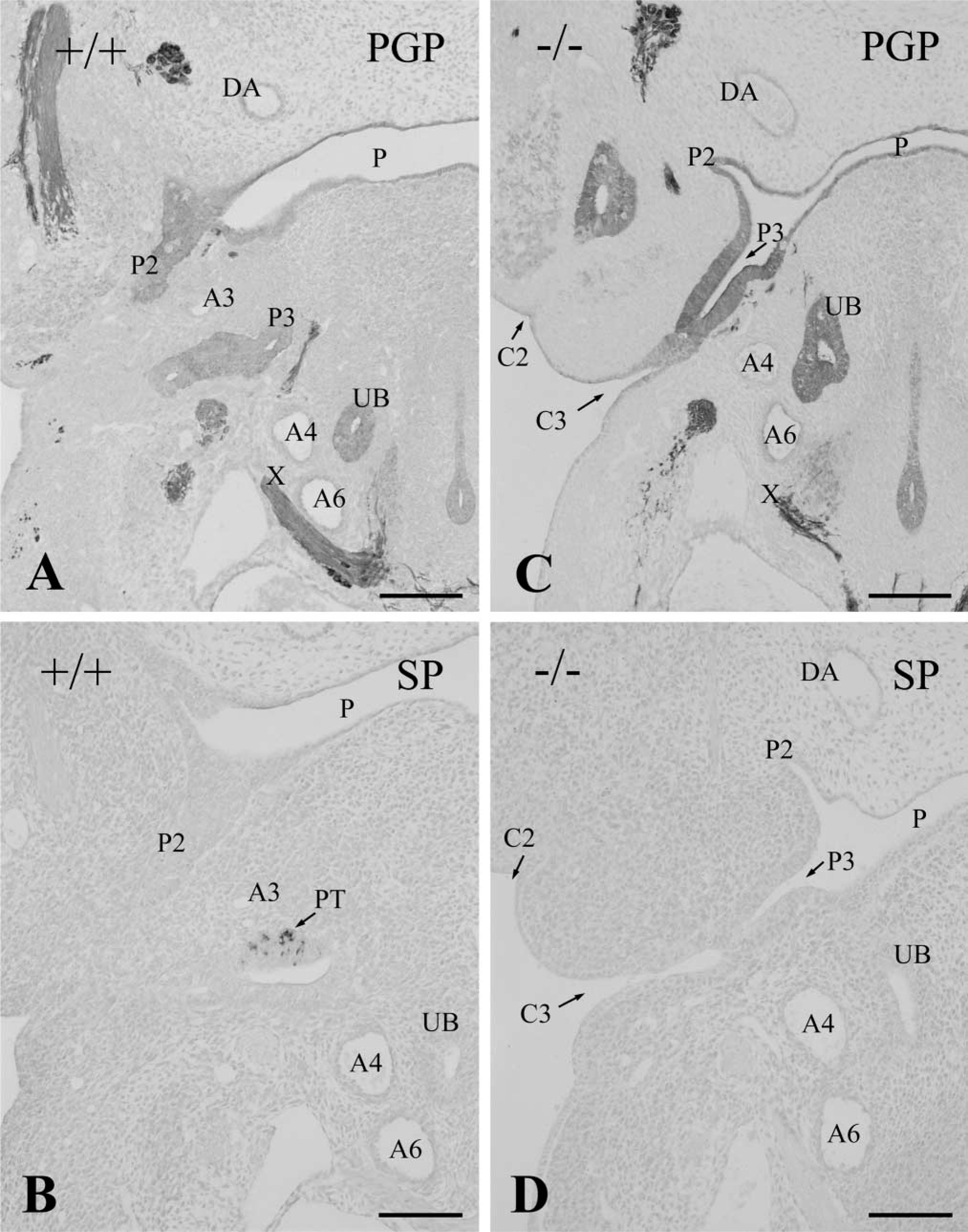
Consecutive frontal sections of E 11.5 littermate wild-type (+/+,
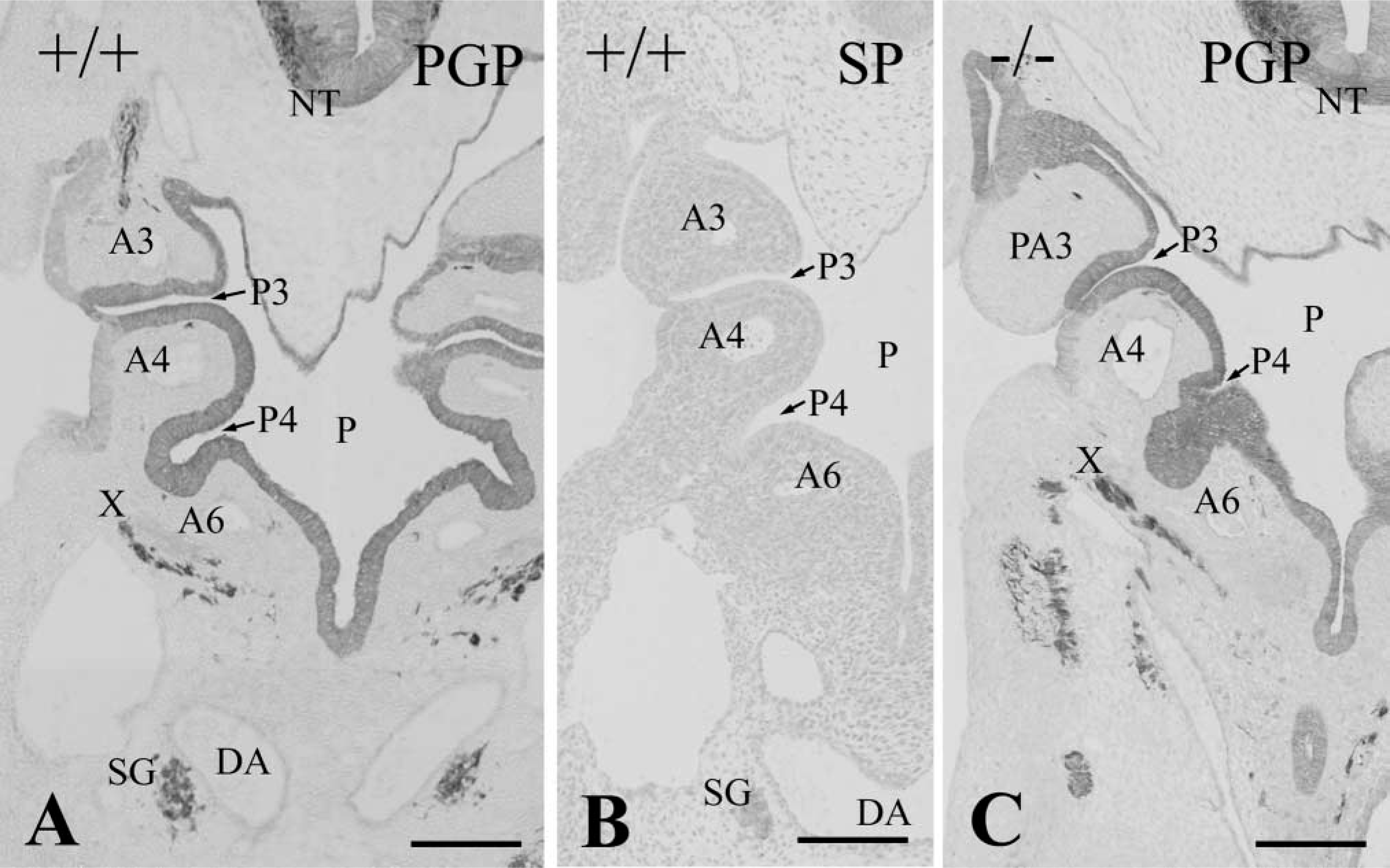
In addition to the differentiation of the third pharyngeal pouch, the Hoxa3 gene appears to play roles in the development of the third pharyngeal arch. In E 11.5 wild-types, the third pharyngeal cleft fused with the second one and the third pharyngeal arch merged into the fourth one (Figure 3A). Therefore, the third pharyngeal arch of wild-types became indistinct at this stage. In contrast, Hoxa3 homozygous mutants at E 11.5 still had the distinct third pharyngeal arch partitioned by the third and second pharyngeal pouches and clefts, showing the delayed differentiation of the third arch (Figure 3C).
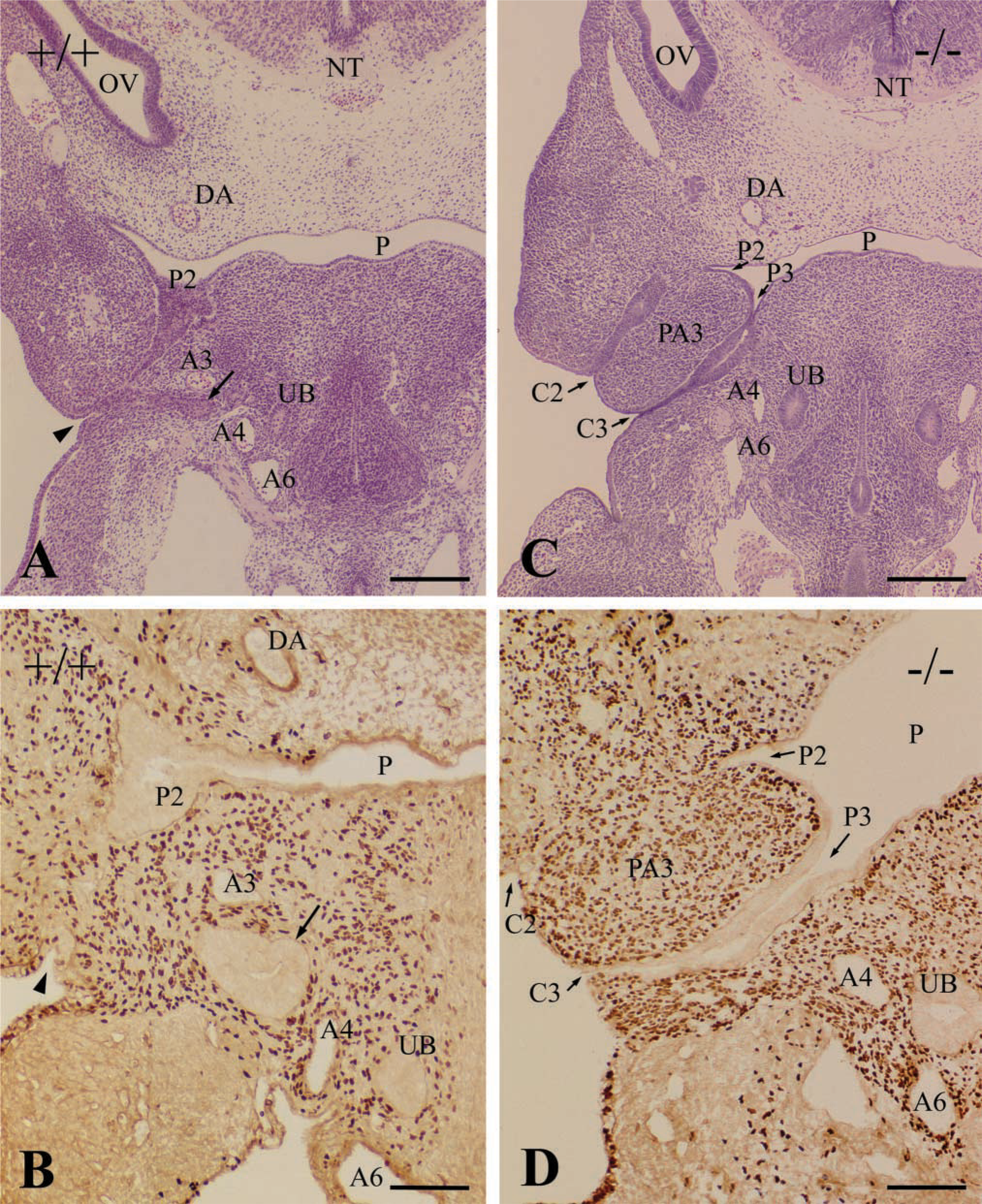
Frontal sections of pharyngeal region in wild-types (+/+,
The migration of ectomesenchymal neural crest cells into the pharyngeal arches was examined by immunostaining with the monoclonal anti-β-galactosidase antibody (Figures 3B and 3D). The β-galactosidase-positive neural crest cells densely populated the third pharyngeal arch in Hoxa3 homozygotes as well as wild-types at E 11.5 (Figures 3B and 3D). In both wild-type and Hoxa3 mutant embryos, the neural crest cells surrounded the parathyroid/thymus primordia or the third pharyngeal pouch but never invaded them.
Subsequently, the time course of parathyroid/thymus primordia development was determined in wild-type embryos. At E 12.0, the thymus rudiment originating from the third pharyngeal pouch became clear and extended in a caudal direction. The thymus still displayed a follicle structure in which the epithelium had many mitotic figures. The parathyroid rudiment was attached to the cranial part of the thymus (Figure 4A). At E 12.5, the thymus rudiment was significantly increased in size and formed a solid cell cluster. It began to move towards the anterior thoracic cavity. The parathyroid rudiment was localized on the top of the thymus (Figure 4B). At E 13.5, owing to a caudal migration of the thymus, the parathyroid rudiment contiguous with the top of the thymus was situated at the side of the thyroid and ultimobranchial glands (Figure 4C). At E 14.5, the parathyroid joined with the lateral side of the thyroid gland and was completely separated from the thymus, showing a normal position in mature mice (Figure 4D).
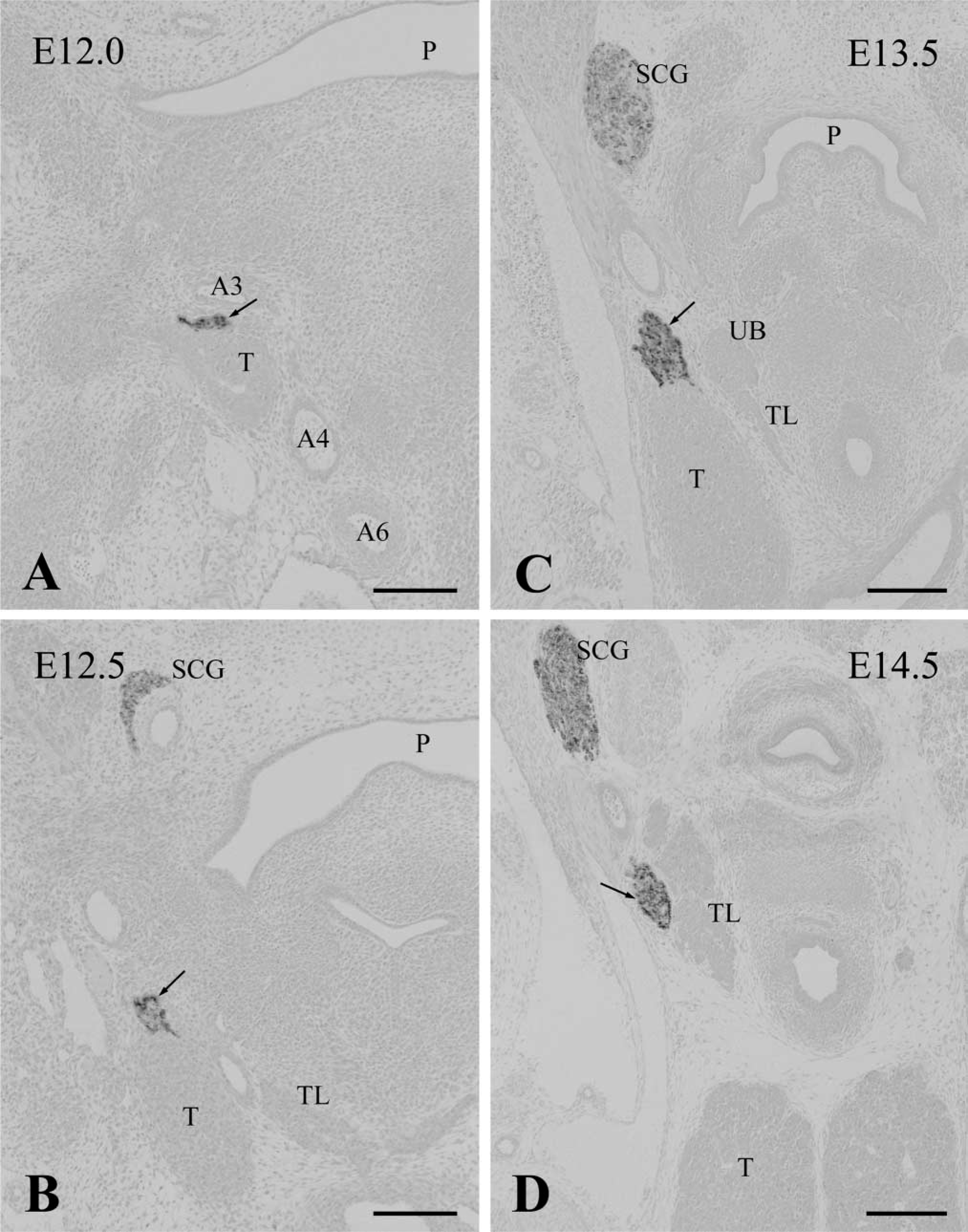
The parathyroid rudiments in wild-type embryos at E 12.0 (
In Hoxa3-null mutants at E 12.0, differentiation of the third pharyngeal pouch did not occur and SP-1/chromogranin A-immunoreactive parathyroid cells were never detected in the third pouch, remaining as rudimentary tissue. Because the separate second and third clefts were still distinct in the mutants, the third pharyngeal arch was discriminated clearly even at E 12.0 (Figure 5A). A small thymus rudiment sometimes appeared in the ectodermal epithelium of the third cleft (Figure 5A). On and after E 12.5, the third pharyngeal pouch disappeared. Therefore, no parathyroid rudiment was formed in the Hoxa3-null mutants (Figure 5B). The thymus lobes were rarely observed in the E 12.5–14.5 null mutants, although their size was far smaller than controls.
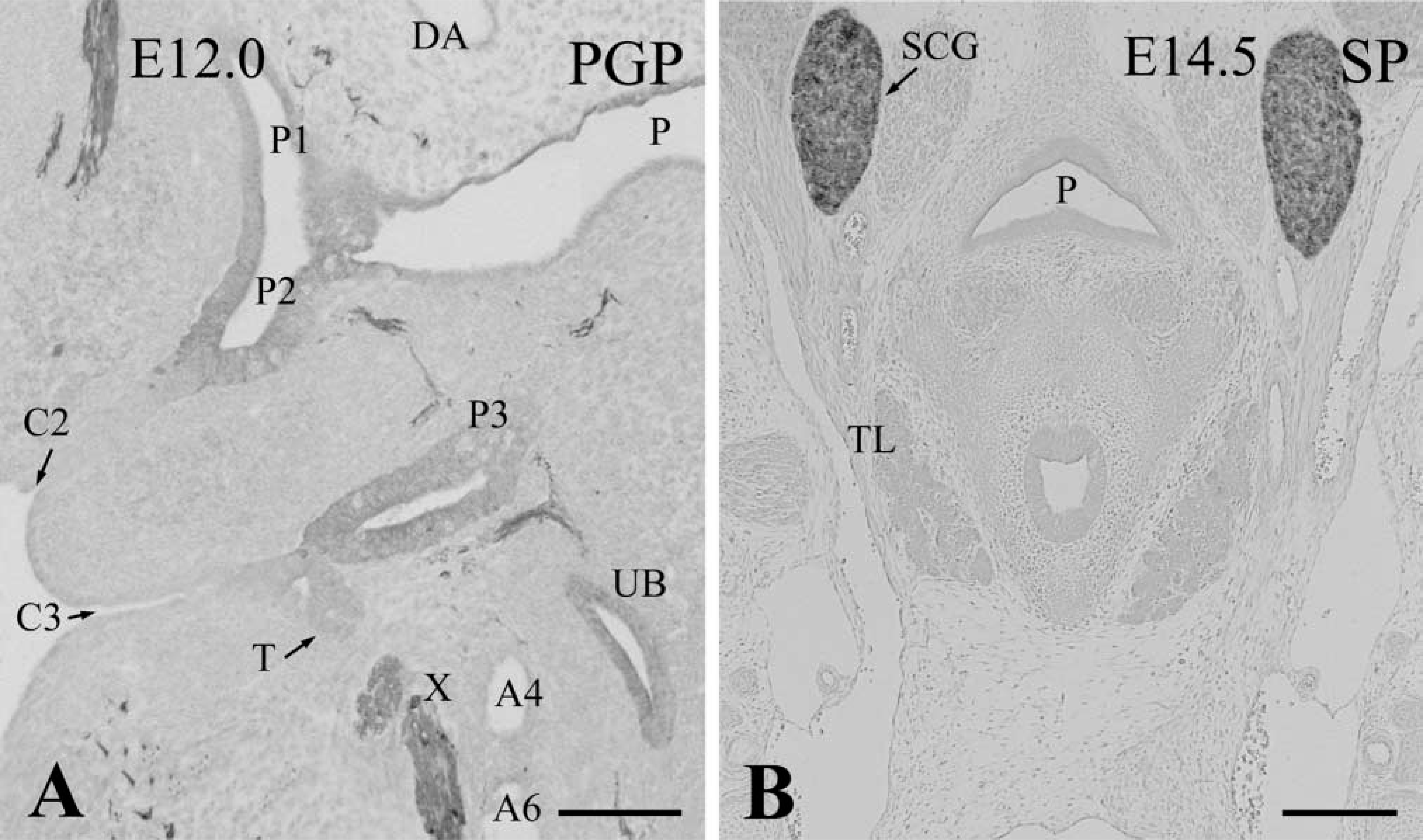
Frontal sections of the pharyngeal region in E 12.0 (
TUNEL assay revealed that high levels of apoptosis occurred in the pharyngeal pouch endoderm, including the second, third, and fourth pouches, in wild-types at E 11.5. In Hoxa3-null mutants at E 11.5–12.0, there was increased apoptosis in the third pouch relative to age-matched control embryos (Figures 6A and 6B). Apoptotic cells frequently aggregated in large clusters in the pouch epithelium (Figures 6C and 6D).
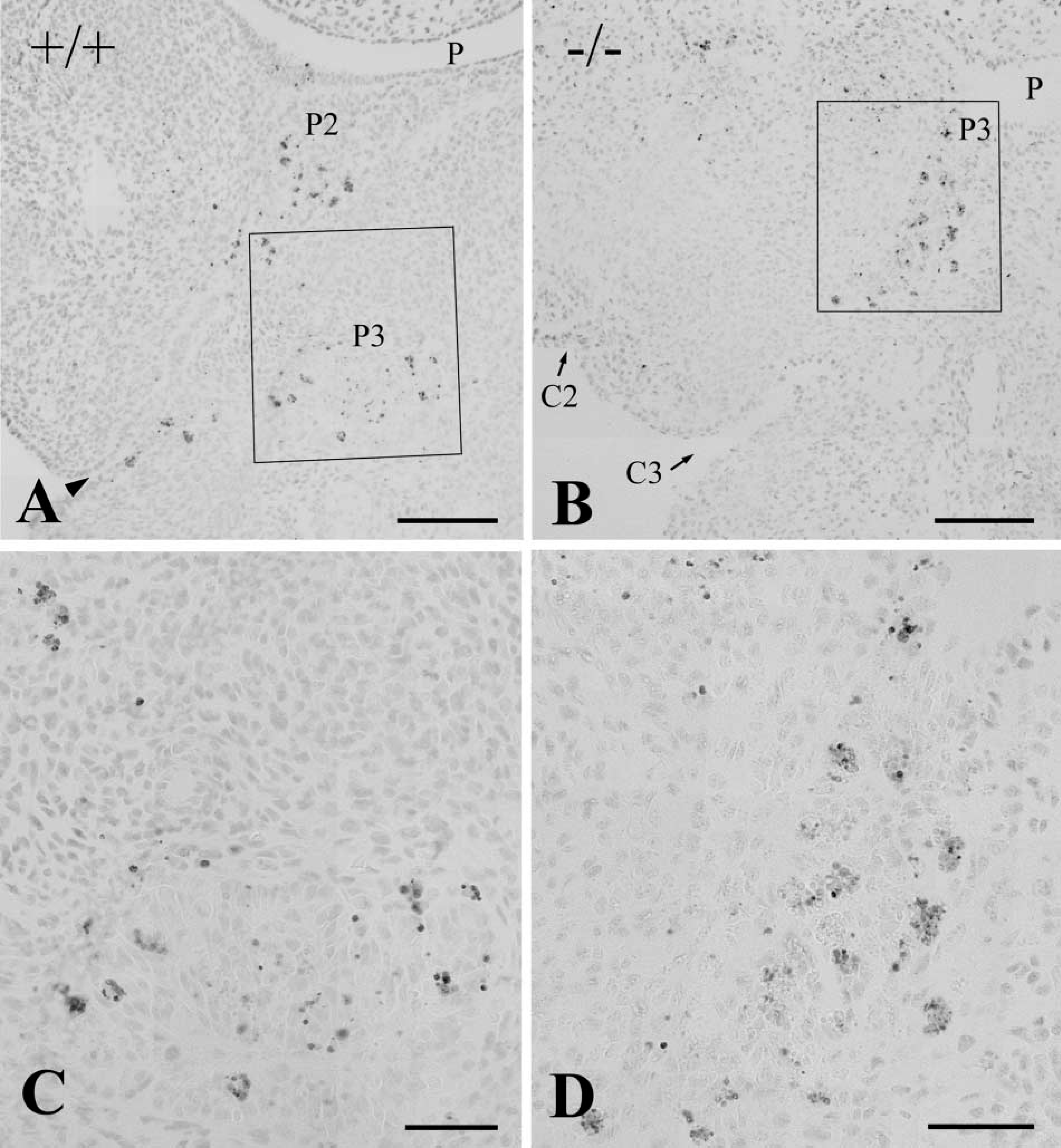
TUNEL analysis of frontal sections through the pharyngeal region in littermate wild-type (+/+,
Discussion
Chromogranin A immunoreactivity is present in virtually all enteroendocrine cells (Portela-Gomes et al. 1997). In human fetuses, abundant chromogranin A-immunoreactive cells are distributed in the earliest (8 weeks of gestation) intestine (Facer et al. 1989). Furthermore, bovine adrenal chromaffin cells have been shown to display immnoreactivity for chromogranin A during their earliest stages of development, at which stage the adrenal medulla has not yet formed and before immunoreactivity for tyrosine hydroxylase appears (Totzauer et al. 1995). Therefore, SP-1/chromogranin A appears to be expressed in progenitors of endocrine cells and to play an important role in the early stages of development. The present study demonstrated that SP-1/chromogranin A immunoreactivity appeared in the parathyroid rudiment at E 11.5, i.e., at the initial stage of parathyroid formation.
Hoxa3 mRNA is expressed in the third pharyngeal pouch endoderm (Manley and Capecchi 1995). The present study demonstrated that the Hoxa3 gene is required for initiation of parathyroid rudiment formation. In wild-type mouse embryos, the differentiation of the third pharyngeal pouch began at E 11.5. The parathyroid rudiment immunoreactive for SP-1/chromogranin A was formed in the cranial dorsal part of the third pouch endoderm and could be discriminated from the thymus at this stage of development. In Hoxa3 homozygous mutants, however, the third pharyngeal pouch still remained in high columnar epithelium and bulges for parathyroid and thymus primordia were not formed at E 11.5. No SP-1/chromogranin A-immunoreactive cells were detected in the third pharyngeal pouch. Therefore, the third pharyngeal pouch failed to differentiate and finally disappeared in the Hoxa3 homozygotes. The intrinsic capability of the third pharyngeal pouch to form the parathyroid glands was affected by the lack of the Hoxa3 gene.
Some candidates for signaling molecules involved in the parathyroid organogenesis have been reported. Glial cells missing (Gcm) 2, a mouse homologue of Drosophila Gcm, is the transcription factor demonstrated in the parathyroid gland in late stages of fetal development (Kim et al. 1998). Gcm2-deficient mice lack parathyroid glands but show normal thymus development (Günther et al. 2000). Gcm2 expression begins at E 9.5 in the caudal pharyngeal pouches and is progressively restricted to the third pharyngeal pouch endoderm (Gordon et al. 2001). In contrast to Gcm expression, SP-1/chromogranin A immunoreactivity was localized only in the parathyroid rudiment in the pharyngeal pouches. Pax1 and Pax9 are closely related members of the paired box gene family. Pax1 is expressed in the third pharyngeal pouch and also in thymic epithelial cells (Wallin et al. 1996). Loss of Pax1 function results in a hypoplastic thymus due to a deficiency in thymocyte development (Wallin et al. 1996; Su et al. 2001). Although hypoplastic parathyroids have also been reported in Pax1 mutant embryos, it is difficult to precisely identify the degenerative organs by hematoxylin-eosin staining (Su et al. 2001). Pax9 mutant mice lack the derivatives of the third and fourth pouches, i.e., thymus, parathyroid, and ultimobranchial glands (Peters et al. 1998). Eyes absent (Eya) 1 gene is expressed in thymus, parathyroid, and ultimobranchial glands during development, and the organ primordia for these structures fail to form in Eya1 –/– embryos (Xu et al. 2002). In Hoxa3 mutants, Pax1 is downregulated in the third pharyngeal pouch (Manley and Capecchi 1995) and Gcm expression is absent (Ellis S. et al., quoted in Su et al. 2001). In Eya1 –/– pouch endoderm, Hoxa3, Pax1, and Pax9 expression is not affected (Xu et al. 2002). It has been suggested that Gcm2 expression in the parathyroid glands is regulated by the Hoxa3-Pax1/9–Eya1 pathway (Xu et al. 2002). Therefore, Hoxa3 seems to be a key regulator in differentiation of the third pouch endoderm.
In human 22q11 deletion syndromes, hypoplastic or aplastic thymus and parathyroid glands are characteristic phenotypic abnormalities (Scambler 2000). Tbx1 is a T-box transcription factor that lies in the 22q11 locus, and Tbx1-null homozygotes lack the thymus and parathyroid glands (Jerome and Papaioannou 2001). Fgf8 mutant mice display pheno-types seen in human 22q11 deletion syndromes, including thymic and parathyroid aplasia and hypoplasia (Frank et al. 2002). The relationships of these genes to Hoxa3 during third pharyngeal pouch development remain to be elucidated.
In Hoxa3 homozygotes crossed with Cx43–lacZ transgenic mice, ectomesenchymal neural crest cells immunoreactive for β-galactosidase densely populated the pharyngeal arches as in wild-type embryos. The migration of mesenchymal neural crest cells into the third arch was not disturbed in the Hoxa3 homozygotes. In avian species, neural crest cells invade the thymus and parathyroid glands and give rise to glandular connective tissue (Le Lièvre and Le Douarin 1975). On the other hand, the present study demonstrated in mouse embryos that neither the endodermal epithelium of the third pharyngeal pouch nor the parathyroid and thymus primordia received the neural crest cells. Furthermore, it has been shown in the mouse by marking the Wnt1-Cre and R26R genes that the mesenchymal neural crest cells surround but do not invade the thymus parenchyma (Jiang et al. 2000). The parathyroid, thymus, and ultimobranchial primordia develop from the pharyngeal pouches and migrate to their final destinations. These organs are densely enclosed by mesenchymal neural crest cells during movement to their locations. The neural crest cells distributed in the pharyngeal arches may be involved in the organ primordia migration in the mouse embryos. In addition, the condensation of mesenchymal neural crest cells during development of pharyngeal pouch-derived organs appears to be accompanied by upregulation of the expression of several growth factors and transcription factors (Clouthier et al. 2000). Appropriate interactions between the pouch epithelium and adjacent neural crest-derived mesenchyme may be necessary for normal development of the parathyroid, thymus, and ultimobranchial glands. Hoxa3 mRNA is expressed in the neural crest cells populating the third pharyngeal arch (Watari et al. 2001). Absence of Hoxa3 signaling appears to affect the proliferation and differentiation of postmigratory neural crest cells in the third arch.
The endodermal epithelia of the pharyngeal pouches in both wild-type and homozygous embryos showed many apoptotic cells. The differentiation and growth of the pharyngeal pouch-derived organs appear to be controlled by apoptosis. In particular, increased apoptotic cells were detected in the third pouch epithelium showing delayed development in the E 11.5–12.0 mutants. Therefore, the third pouch endoderm is unable to differentiate in the absence of the Hoxa3 gene and is eventually lost by apoptosis. Although a parathyroid rudiment was never detected, rudimentary thymus lobes were rarely encountered in the null mutants at E 12.5–14.5. The thymus is formed by the third pouch endoderm fused with the ectodermal cells of the third cleft (Cordier and Haumont 1980). The ectodermal cells may be unaffected in the Hoxa3 mutants and may have the ability to form unstable thymus lobes.
In contrast to the third pharyngeal pouch, the development and differentiation of the fourth pouch were normal in Hoxa3-null mutants. The bilateral ultimobranchial bodies derived from the fourth pharyngeal pouch were always formed and joined with the thyroid glands at E 13.5. At E 14.5, the C-cells immunoreactive for PGP 9.5 began to disperse within the thyroid parenchyma in the Hoxa3 mutants as well as wild-types (Kameda et al. unpublished data). In both wild-type and mutant mice, however, SP-1/chromogranin A immunoreactivity appeared in the C-cells at late stages of fetal development. Furthermore, the appearance of calcitonin immunoreactivity in the C-cells was very late, i.e., at birth. In accordance with our data, Xu et al. (2002) have reported that calcitonin immunoreactivity is first detected in newborn mice.