
Abstract
The functional upper esophageal sphincter (UES) is composed of the cricopharyngeus muscle (CP), the most inferior part of the inferior pharyngeal constrictor (iIPC), and the upper esophagus (UE). This sphincter is collapsed and exhibits sustained muscle activity in the resting state; it only relaxes and opens during swallowing, vomiting, and belching. The tonic contractile properties of the UES suggest that the skeletal muscle fibers in this sphincter differ from those in the limb and trunk muscles. In this study, myosin heavy chain (MHC) composition in the adult human UES muscles obtained from autopsies was investigated using immunocytochemical and immunoblotting techniques. Results showed that the adult human UES muscle fibers expressed unusual MHC isoforms such as slow-tonic (MHC-ton), α-cardiac (MHC-α), neonatal (MHC-neo), and embryonic (MHC-emb), which coexisted with the major MHCs (i.e., MHCI, IIa, and IIx). MHC-ton and MHC-α were coexpressed predominantly with slow-type I MHC isoform, whereas MHC-neo and MHC-emb coexisted mainly with fast-type IIa MHC. A slow inner layer (SIL) and a fast outer layer (FOL) in the iIPC and CP were identified immunocytochemically. MHC-ton- and MHC-α-containing fibers were concentrated mainly in the SIL, whereas MHC-neo- and MHC-emb-containing fibers were distributed primarily to the FOL. Identification of the specialized muscle fibers and their distribution patterns in the adult human UES is valuable for a better understanding of the physiological and pathophysiological behaviors of the sphincter.
Keywords
U
The contractile properties of a muscle are determined primarily by the fiber type and MHC composition. In addition to the three major fiber types (i.e., type I, IIA, and IIX) and corresponding MHC isoforms (i.e., MHCI, MHCIIa, and MHCIIx) that are widely present in the adult human skeletal muscles (Schiaffino and Reggiani 1994), some unusual MHC isoforms [i.e., slow-tonic (MHC-ton), α-cardiac (MHC-α), neonatal (MHC-neo), and embryonic (MHC-emb)] not found in normal adult limb and trunk muscles have also been identified in some specialized cranial muscles (i.e., extraocular, masseter, middle ear, laryngeal, and suprahyoid muscles) (for review, see Mu et al. 2004; Ren and Mu 2005). We hypothesized that the consistent tonic contraction of the UES at rest may contribute to its specialized fibers expressing unique and unusual MHC isoforms.
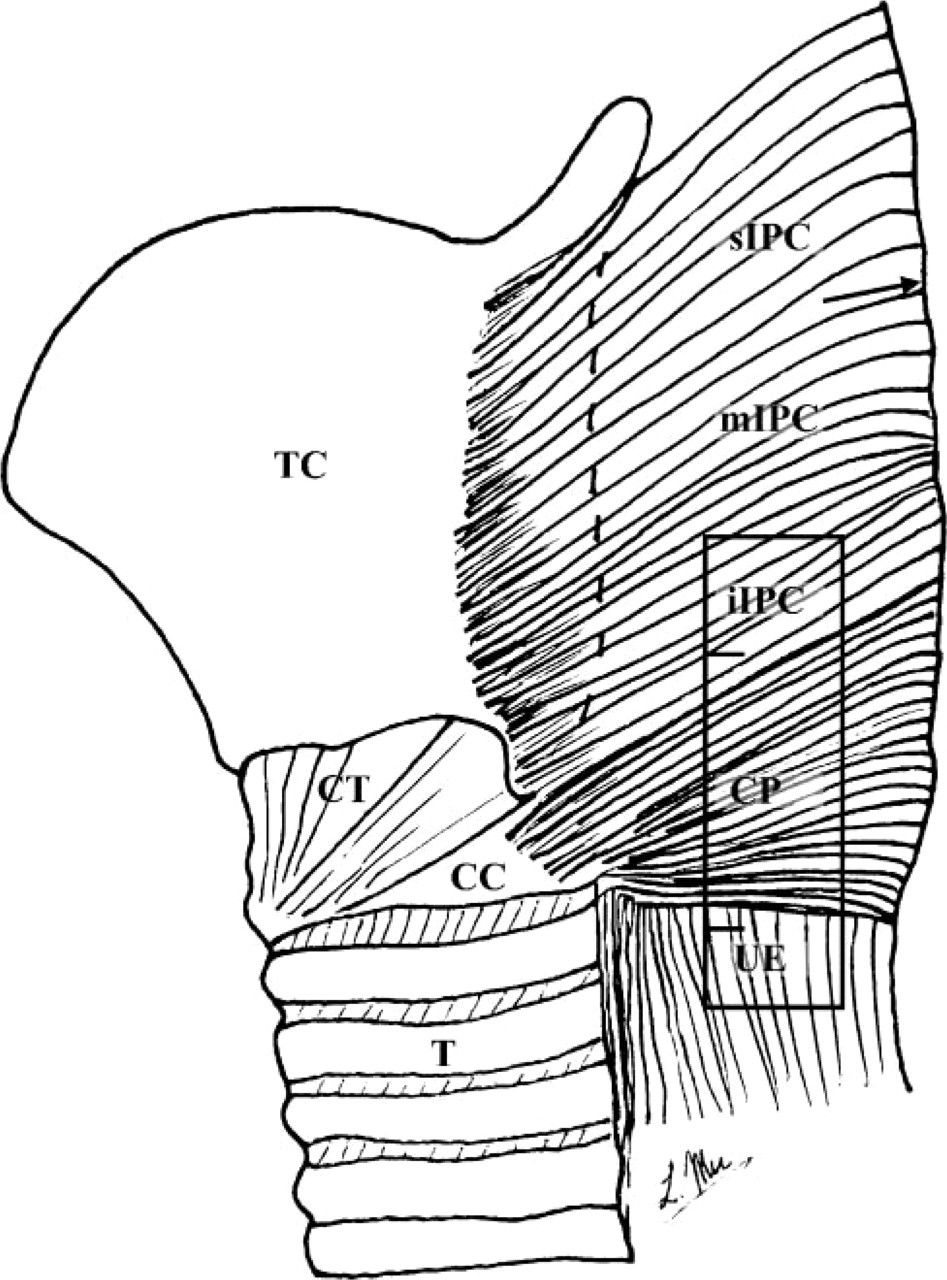
Schematic of the human upper esophageal sphincter (UES) and its surrounding structures (lateral view), showing the tissue sampling position (enclosed region) in the most inferior portion of the inferior pharyngeal constrictor (iIPC), cricopharyngeus (CP), and upper esophagus (UE) for immunocytochemistry. CC, cricoid cartilage; CT, cricothyroid muscle; mIPC, middle portion of the inferior pharyngeal constrictor muscle; sIPC, superior portion of the inferior pharyngeal constrictor muscle; T, trachea; TC, thyroid cartilage. Arrow indicates the median raphe of the IPC muscle.
This study was designed to detect whether the adult human UES muscles have been specialized by containing muscle fibers expressing unusual MHC isoforms for a better understanding of the contractile properties of the sphincter.
Materials and Methods
Tissue Sampling
Four adult human pharynges were obtained from autopsies (two males and two females; mean age 55 years) without known systemic neuromuscular disorders. UES muscles including the iIPC, CP, and UE (n=8 each) were sampled as illustrated by Figure 1 for immunocytochemistry. In addition, mylohyoid (MH) and biceps brachii were sampled from the same subject to serve as positive and negative controls, respectively. These muscle samples were placed in parallel, snap frozen together in isopentane cooled by dry ice, cut transversely (10-μm thick) at −25C on a cryostat (Reichert-Jung 1800; Mannheim, Germany), and stored at −70C until staining procedures were performed. Small muscle samples were also taken from each muscle studied for electrophoretic immunoblotting.
Immunocytochemistry
Monoclonal Antibodies (MAbs). Seven type-specific anti-MHC MAbs (Table 1) were used to incubate the serial cross-sections from each muscle for identifying the three major (type I, IIa, and IIx) and four unusual (MHC-ton, MHC-α, MHC-emb, and MHC-neo) MHC isoforms. Specificity of these MAbs has been established by our recent studies and others (for review, see Mu et al. 2004).
Avidin—Biotin—Complex (ABC) Method. Muscle fibers containing the major MHC isoforms were identified on the sections incubated with the MAbs NOQ7-5-4D, MY-32, and SC-71 using the ABC method according to Mu et al. (2004). Briefly, serial cross-sections were fixed in 4% paraformaldehyde for 10 min, blocked in 2% BSA with 0.1% Triton X-100 for 20 min, incubated with anti-MHC MAbs for 1 hr at room temperature (MAb NOQ7-5-4D) or overnight at 4C (MAbs MY-32 and SC-71), processed with a Vectastain antimouse IgG (ATCC; Rockville, MD) for 1 hr at room temperature and reacted for 1 hr at room temperature in ABC reagent, reacted for ∼5-10 min at room temperature with a solution containing 3,3′-diaminobenzidine (DAB) as chromogen to localize peroxidase for primary antibodies according to DAB substrate kit (SK-4100; Vector Laboratories, Burlingame, CA), dehydrated in graded concentrations of ethanol, cleared in xylene, and mounted with Permount. Two additional control sections were treated using the aforementioned procedures but without incubation with a primary antibody.
Specificity and characterization of the monoclonal antibodies (MAbs) used ∗
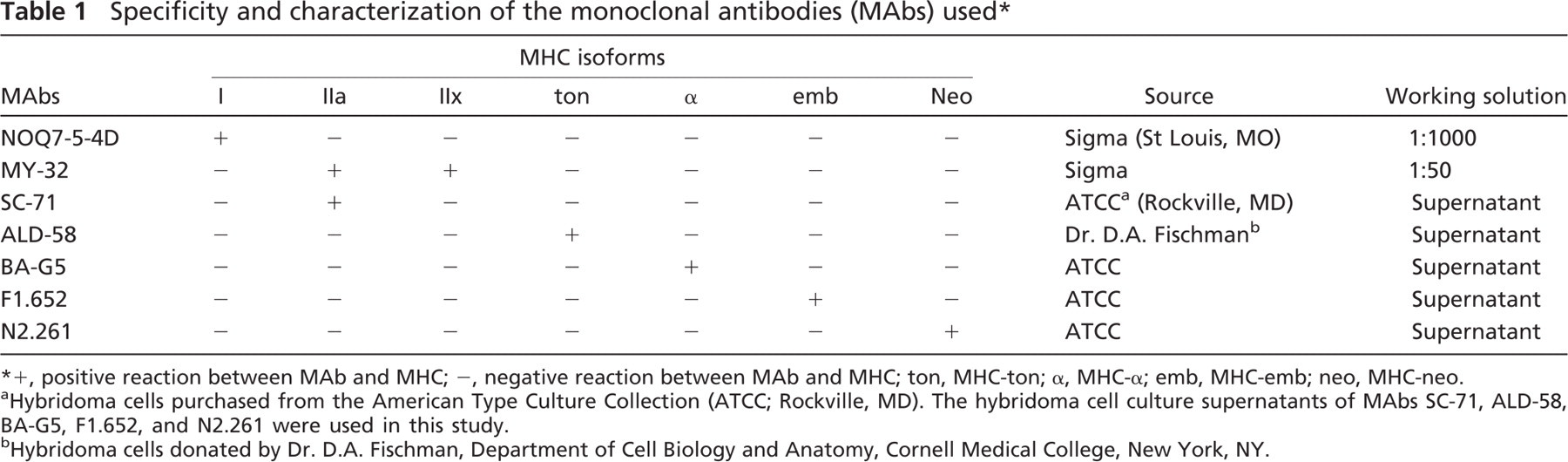
+, positive reaction between MAb and MHC; -, negative reaction between MAb and MHC; ton, MHC-ton; α, MHC-α emb, MHC-emb; neo, MHC-neo.
Hybridoma cells purchased from the American Type Culture Collection (ATCC; Rockville, MD). The hybridoma cell culture supernatants of MAbs SC-71, ALD-58, BA-G5, F1.652, and N2.261 were used in this study.
Hybridoma cells donated by Dr. D.A. Fischman, Department of Cell Biology and Anatomy, Cornell Medical College, New York, NY.
Immunofluorescence. Indirect immunofluorescent staining according to the protocols described by Mu et al. (2004) was used to identify the unusual MHC-containing fibers in the UES muscles. Serial sections adjacent to those stained with ABC method were fixed in 4% fresh paraformaldehyde for 10 min, blocked in 2% BSA with 0.1% Triton X-100 for 20 min, incubated with four anti-MHC MAbs at 4C for either 12 hr (MAbs N2.261 and F1.652) or 24 hr (MAbs ALD-58 and BA-G5), blocked again with 1% BSA at room temperature for 5 min, incubated in a fluorescein isothiocyanate-labeled goat antimouse IgG (FITC-GAM, 1:50 dilution; Sigma, St Louis, MO) at room temperature for 60 min, washed in PBS (3 × 5 min), mounted with Vectashield mounting medium (Vector Laboratories), and kept in the dark at 4C. Control sections were stained as described above except that incubation with the primary antibody was omitted.
Identification of the Unusual MHC-containing Fibers
Stained serial cross-sections were viewed under a Zeiss photo-microscope (Axiophot-2; Carl Zeiss, Goettingen, Germany) equipped with epifluorescence optics and differential interference contrast. Sections were photographed using a digital camera (Spot-32; Diagnostic Instruments, Keene, NH) attached to the photomicroscope and connected to a personal computer with image analysis software. Because the adult human iIPC and CP muscles are composed of a slow inner layer (SIL) and a fast outer layer (FOL) (Mu and Sanders 2001,2002), photomicrographs were taken from each of the fiber layers in both muscles.
Positive and negative fibers that reacted to a specific MAb were manually identified and marked using a computerized image analysis system (SigmaScan; Jandel Scientific, San Rafael, CA). The unusual MHC-containing fibers were counted. The proportion and distribution of each of the unusual MHC-containing fibers were determined.
Electrophoretic Immunoblotting
Tissue Preparation. Electrophoretic immunoblotting was used to confirm the existence or absence of a given unusual MHC isoform according to the methods described by Mu et al. (2004). Small blocks of muscle tissue were minced and ground. The ground tissue was then suspended in 0.1 M Tris buffer containing 1 mM EDTA (pH 7.5) and mixed with a Vortex mixer to form crude homogenate. The crude homogenate was spun at 800 rpm for 10 sec to remove large tissue debris. The supernatant was collected and mixed with the sample buffer in a 2:1 ratio (Hames 1990). Samples were heated in the boiling water bath for 4 min, gradually cooled to room temperature, and stored at −20C before use. Protein concentration was assayed by using an Ultrospec 2000 spectrophotometer (Pharmacia Biotech; Piscataway, NJ). Ten to 30 μg protein was loaded onto each well of the gel.
Electrophoresis. One-dimensional SDS-PAGE was performed using the protocols described by Mu et al. (2004). Electrophoresis was carried out at 100 V in a 4C refrigerator for 2 hr until the bromophenol blue tracking dye migrated to the gel bottom. Some of the gels were stained with Coomassie Blue and others were used for immunoblotting.
Immunoblotting. After electrophoresis, the displayed proteins on the gel were transferred to an immobilon-P PVDF membrane (Millipore; Bedford, MA) by electroblotting in a Transphor Tank transfer unit (Pharmacia Biotech) at 70 V for 1 hr at 4C. After transfer, the membrane with target protein was immersed for 30 min in a 5% BSA blocking solution and incubated overnight in 1:500 diluted supernatant of MAbs. The membrane was then washed and incubated for 1 hr at room temperature in 1:1000 diluted horseradish peroxidase-conjugated goat antimouse IgG (Amersham; Arlington Heights, IL). Immunoblot was developed with enhanced chemiluminescence (ECL) detection reagents (Amersham) according to Durrant and Fowler (1994). Chemiluminescent signal was captured by Polaroid film using an ECL minicamera (Amersham).
Data Analysis
Relative proportions of various unusual MHC-containing fibers in each of the muscles and muscle regions were determined by averaging overall samples. All data were reported as means ± SE. Mean values for various unusual MHC-containing fibers were compared among the studied muscles (i.e., iIPC, CP, and UE) and between distinct regions (i.e., SIL and FOL in the iIPC and CP muscles). One-way ANOVA was performed for intra- and intermuscular comparisons of different variables. Significant differences between the means were then determined using Student's t-test, and statistical significance was accepted at p<0.05.
Results
SIL and FOL in the Adult Human iIPC and CP Muscles
Serial cross-sections immunocytochemically stained with the MAbs specific to the major MHC isoforms (i.e., MAbs NOQ7-5-4D to type I, MY-32 to all fast-type II, and SC-71 to type IIa) showed that the adult human iIPC and CP muscles were composed of a SIL and a FOL (Figures 2A and 2B) as demonstrated by myofibrillar adenosine triphosphatase (mATPase) staining (Mu and Sanders 2001,2002). The SIL was situated just beneath the mucosa, whereas the FOL consisted of the outer portion of the muscles. Slow-type I MHC-containing fibers were concentrated predominantly in the SIL, whereas fast-type II MHC-containing fibers were distributed mainly to the FOL. The ratio of the thickness of the SIL to FOL was ∼1:1 for the iIPC and ∼2:1 for the CP.
Expression of the Unusual MHC Isoforms in the UES Muscle Fibers
Adult human UES muscle fibers expressed unusual MHC isoforms (i.e., MHC-ton, MHC-α, MHC-neo, and MHC-emb). The unusual MHC-containing fibers in the iIPC and CP muscles were distributed in a fiber layer-dependent manner. Specifically, MHC-ton- (Figure 2C) and MHC-α- (data not shown) containing fibers were concentrated mainly in the SIL, whereas muscle fibers expressing MHC-neo (Figure 2D) and MHC-emb (data not shown) isoforms were distributed primarily to the FOL in the iIPC and CP muscles. In the control muscles, similar unusual MHC isoforms were also found in the MH muscle but not in the biceps brachii.
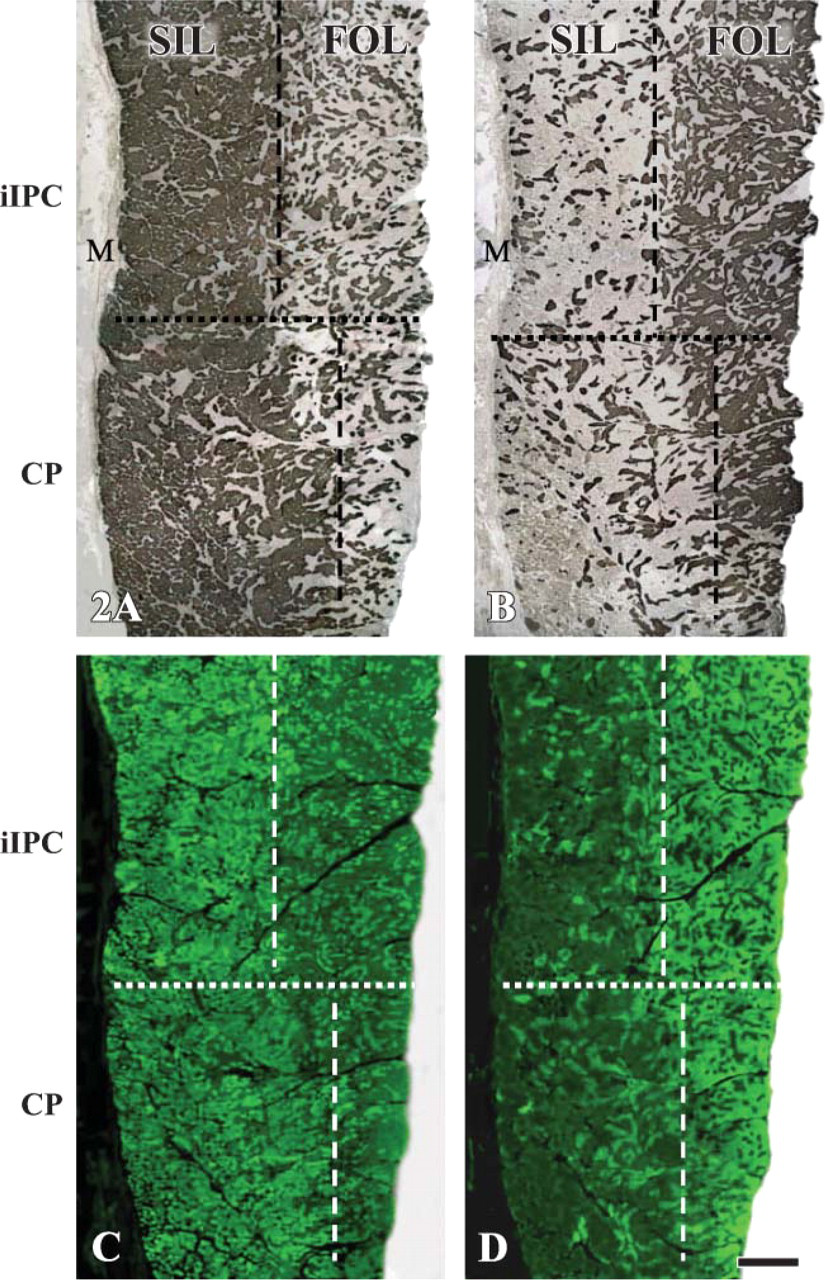
Low-power view of the immunocytochemically stained serial cross-sections from the adult human iIPC (top) and CP (bottom) muscles, showing the slow inner layer (SIL) and fast outer layer (FOL) in both muscles. The sections were stained with the monoclonal antibodies (MAbs) NOQ7-5-4D specific for type I (
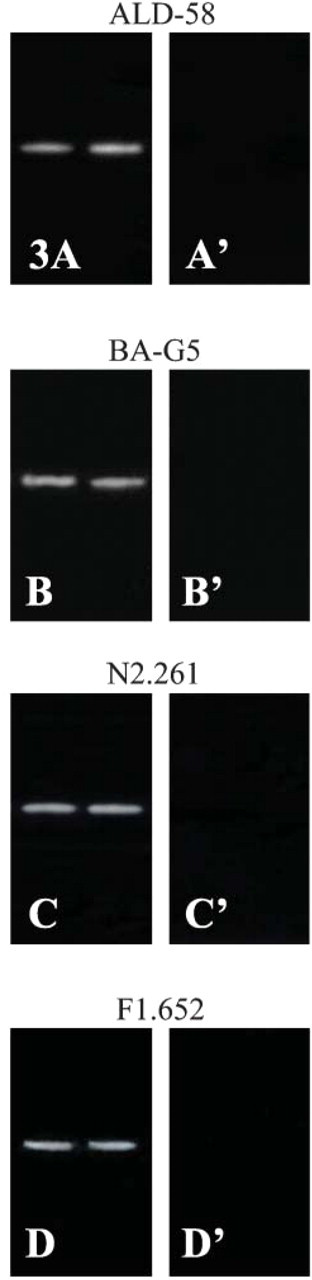
Immunoblotting analysis of unusual MHC isoforms in adult human CP and biceps brachii (control) muscles. Electrophoretic immunoblotting was performed for the muscle samples studied. After transfer, membranes with proteins were incubated with MAbs ALD-58 specific to MHC-ton (
Electrophoretic immunoblotting confirmed the existence of the four unusual MHC isoforms in the adult human CP (Figures 3A-3D), iIPC, and UE (data not shown). Existence of the four unusual MHC isoforms was also confirmed in the MH control muscle (data not shown), but not in the biceps brachii (Figures 3A′-3D′).
Coexistence of the Unusual MHC Isoforms with the Major MHCs
The unusual MHC isoforms were found to coexist with the major MHCs. Coexpression of the unusual MHC isoforms with the major MHCs was identified by comparing all the stained serial cross-sections. The unusual MHC isoforms coexisted in a fiber type-specific manner. Specifically, MHC-ton and MHC-α were coexpressed predominantly with slow-type I MHC, whereas MHC-neo and MHC-emb coexisted primarily with fast-type II MHCs (Figure 4).
In the UES muscles, no pure fibers expressing a single MHC isoform were identified. Instead, a single UES fiber was found to contain two to six MHC isoforms. Fibers expressing two to three MHC isoforms were predominant (70% of the total fiber population). Almost half of the UES fibers expressed three MHC isoforms.
Proportions of Various Unusual MHC-containing Fibers in Each of the UES Muscles
On average, proportions of the iIPC, CP, and UE fibers expressing the individual unusual MHC isoforms were ∼75% for either MHC-ton fibers or MHC-α fibers, 40% for MHC-neo fibers, and 8% for MHC-emb fibers. Evidently, MHC-ton- and MHC-α-containing fibers were much more than the MHC-neo- and MHC-emb-containing fibers in the UES muscles (p<0.01).
However, differences in the percentage distribution of a given unusual MHC-containing fiber were noted among muscles (i.e., iIPC, CP, and UE) and between muscle regions (i.e., SIL and FOL in the iIPC and CP muscles). For instance, the UE contained more MHC-ton and MHC-α fibers (95%) than the iIPC (60%) and CP (68%) (p<0.01). The SIL of the iIPC and CP muscles contained more MHC-ton and MHC-α (75%) fibers than the FOL of the muscles (55%) (p<0.01). In contrast, the FOL in the iIPC and CP muscles contained more MHC-neo fibers (46%) as compared with the SIL (25%) of the muscles (p<0.01). However, there was no difference in the distribution of MHC-emb fibers among muscles and between muscle regions studied (p>0.05).
Discussion
To our knowledge, this is the first study to detect unusual MHC isoforms in adult human UES muscles. The notable findings are as follows: (1) Our previously described histochemical SIL and FOL in the adult iIPC and CP muscles (Mu and Sanders 2001,2002) were also delineated by using immunocytochemistry. (2) Adult human UES muscle fibers expressed unusual MHC isoforms (i.e., MHC-ton, MHC-α, MHC-neo, and MHC-emb) not identified in the biceps brachii muscle fibers. (3) Unusual MHC isoforms were expressed in a fiber type-specific manner. Specifically, MHC-ton and MHC-α coexisted predominantly with slow-type I MHC isoform, whereas MHC-neo and MHC-emb were coexpressed mainly with fast-type II MHC isoforms. (4) Unusual MHC-containing fibers were distributed in a fiber layer-dependent manner. Specifically, MHC-ton- and MHC-α-containing fibers were concentrated predominantly in the SIL, whereas MHC-neo- and MHC-emb-containing fibers were distributed primarily to the FOL in the iIPC and CP muscles. (5) Pure muscle fibers commonly seen in the limb muscles were extremely rare or absent in the UES. Instead, almost all adult human UES muscle fibers were hybrid fibers that expressed multiple (two to six) MHC isoforms. These observations indicate that the adult human UES muscles have been specialized as indicated by containing fibers expressing several unusual MHC isoforms. The unusual MHC-containing fibers in the UES may play an important role in the regulation of the muscle tone to maintain the tonic contraction of the sphincter in the resting state.
Unusual MHC-containing Fibers in the Specialized Cranial Muscles Including the UES
Unusual MHC-containing fibers have been identified in certain normal mammalian cranial muscles, but not in limb and trunk muscles. For example, MHC-ton-containing fibers common in lower vertebrates and birds (Hess 1970) have been found in extraocular muscles (Pierobon-Bormioli et al. 1979; Sartore et al. 1987), middle ear (Mascarello et al. 1982), laryngeal muscles (Han et al. 1999), masseter (Mu et al. 2004; Ren and Mu 2005), suprahyoid muscles (Mu et al. 2004; Wang et al. 2004; Ren and Mu 2005), UES (present study), and in muscle spindles (Eriksson et al. 1994).
MHC-α, one of the major isoforms in the heart, has been identified in extraocular muscles (Wieczorek et al. 1985; Pedrosa-Domellof et al. 1992; Rushbrook et al. 1994; Rubinstein and Hoh 2000), masseter (Bredman et al. 1991; Pedrosa-Domellof et al. 1992; Stal et al. 1994; Kwa et al. 1995; Korfage et al. 2000; Mu et al. 2004; Ren and Mu 2005), suprahyoid muscles (Mu et al. 2004; Wang et al. 2004; Ren and Mu 2005), UES (present study), and in muscle spindles (Michel et al. 1996).
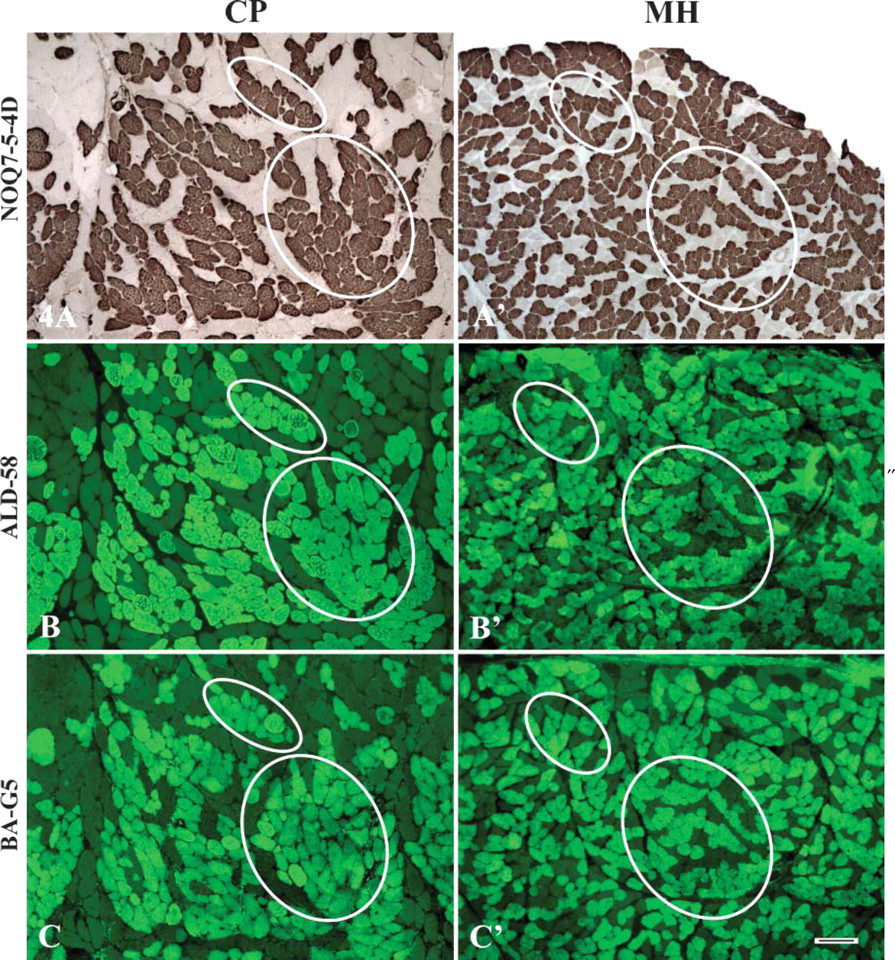
Photomicrographs of serial cross-sections from the adult human CP (left column) and mylohyoid (MH) (right column; control) muscles, showing the coexpression patterns of the unusual MHC isoforms with the major MHCs. The sections were stained with MAbs NOQ7-5-4D specific to slow-type I MHC isoform (
MHC-neo and MHC-emb (developmental MHCisoforms) normally expressed in developing muscle fibers and completely substituted by the adult isoforms in limb muscles still persist in some adult cranial muscles such as extraocular muscles (Wieczorek et al. 1985; Sartore et al. 1987; Rubinstein and Hoh 2000), laryngeal muscles (Jung et al. 1999), masseter (d'Albis et al. 1986; Butler-Browne et al. 1988; Eriksson et al. 1994; Sciote et al. 1994; Stal et al. 1994; Korfage et al. 2000; Mu et al. 2004; Ren and Mu 2005), suprahyoid muscles (Mu et al. 2004; Wang et al. 2004; Ren and Mu 2005), UES (present study), and in muscle spindles (Rowlerson et al. 1985).
The aforementioned unusual MHC isoforms in cranial muscle fibers coexist with the major MHCs and are expressed in a muscle- and fiber type-specific manner. For example, MHC-ton is coexpressed with type I MHC isoform in the tensor tympani (Mascarello et al. 1982), suprahyoid (Mu et al. 2004; Wang et al. 2004; Ren and Mu 2005), and UES (present study) muscles, but with type IIa MHC isoform in masseter (Mu et al. 2004; Ren and Mu 2005). MHC-α is coexpressed with type I (Kwa et al. 1995) or type IIa MHC isoform (Kwa et al. 1995; Mu et al. 2004; Ren and Mu 2005) in masseter or with type I in masseter (Bredman et al. 1991), extraocular (Rushbrook et al. 1994), MH (Mu et al. 2004; Ren and Mu 2005), and UES (present study) muscles. In contrast, MHC-neo coexists with type IIa and IIx MHC isoforms in the MH (Mu et al. 2004; Ren and Mu 2005) and UES (present study) muscles or with type I (Butler-Browne et al. 1988; Mu et al. 2004; Ren and Mu 2005), type IIx (Mu et al. 2004; Ren and Mu 2005), and type IIa and IIb MHC isoforms (Butler-Browne et al. 1988) in masseter. MHC-emb is coexpressed with IIa MHC in masseter (Mu et al. 2004; Ren and Mu 2005) or with type IIa and IIx MHC isoforms in the MH (Mu et al. 2004; Ren and Mu 2005) and UES (present study) muscles.
The difference in the coexpression patterns of the unusual MHC isoforms with the major MHCs as revealed in different cranial muscles may be related to their functional requirements.
Functional Implications of the Specialized Muscle Fibers and Fiber Layers in the Adult Human UES
MHC composition in a single muscle fiber is the primary determinant of contraction speed. Maximal shortening velocity of the major fiber types in mammalian skeletal muscles is ranked IIB > IIX > IIA > I (Reiser et al. 1985a; Bottinelli et al. 1991; Galler et al. 1994). However, there are no pure fibers in the adult human UES. The UES is composed of specialized skeletal muscle fibers that express unusual MHC isoforms under normal conditions. The unusual MHC isoforms expressed by the UES and other cranial muscle fibers coexist with the major MHC isoforms. Unfortunately, the contractile properties of the unusual MHC-containing fibers remain unknown. Previous studies showed that the MHC-ton fibers do not respond with a propagated action potential to a single nerve stimulation and require several seconds to reach maximum tension (Morgan and Proske 1984). MHC-α fibers have shortening speeds intermediate between type I and type IIA fibers (Kwa et al. 1995; Sciote and Kentish 1996). MHC-neo and MHC-emb fibers have lower shortening velocity compared with adult fast MHC-containing fibers (Reiser et al. 1985b). These observations suggest that the adult human UES would yield a greater variance in shortening velocities and maximal force generation suitable for the unique performances of the sphincter.
Adult human iIPC and CP muscles are composed of a SIL and a FOL as revealed by mATPase staining (Mu and Sanders 2001,2002) and immunocytochemistry (present study). However, the functional implications of both fiber layers in the UES remain unknown. Previous electromyographic (EMG) studies demonstrated that the UES is active during swallowing (Shipp et al. 1970; Elidan et al. 1990), respiration (Kawasaki and Takenouchi 1964), and speech (Baer et al. 1976). Unfortunately, no studies were designed to perform EMG recordings from each of the fiber layers. This study showed that MHC-ton- and MHC-α-containing fibers were concentrated preferably in the SIL, whereas MHC-neo- and MHC-emb-containing fibers were distributed primarily to the FOL. The layer-dependent MHC expression patterns suggest that both fiber layers may have distinct contractile properties and play different roles in upper airway functions. Previous studies showed that MHC-ton-containing fibers exist only in the muscles that perform fine movements as required by specific physiological tasks (Pierobon-Bormioli et al. 1979; Mascarello et al. 1982; Han et al. 1999). The SIL with predominance of slow type I, MHC-ton- and MHC-α-containing fibers appears to be well adapted for tonic contraction, fine movements, and postural adjustments that occur during certain motor tasks such as resting closure of the sphincter, respiration, and speech. In contrast, the FOL with predominance of fast type II, MHC-neo- and MHC-emb-containing fibers may facilitate rapid contraction during swallowing.
Further research is needed to determine the MHC-based fiber types and their distribution in the adult human UES and to explore the differences in EMG activities between the SIL and the FOL in the iIPC and CP muscles. Such data would be valuable for further investigation of the alterations in the fiber type and MHC composition of the UES induced by either normal aging or some pathological conditions.
Footnotes
Acknowledgements
This work was supported by NIH Grant 5 R01 DC-04728 from the National Institute on Deafness and Other Communication Disorders (to LM).
The authors are greatly indebted to the Department of Pathology of The Mount Sinai School of Medicine for providing specimens for this study.