
Abstract
We have previously demonstrated that α-smooth muscle (α-SM) actin is predominantly distributed in the central region and β-non-muscle (β-NM) actin in the periphery of cultured rabbit aortic smooth muscle cells (SMCs). To determine whether this reflects a special form of segregation of contractile and cytoskeletal components in SMCs, this study systematically investigated the distribution relationship of structural proteins using high-resolution confocal laser scanning fluorescent microscopy. Not only isoactins but also smooth muscle myosin heavy chain, α-actinin, vinculin, and vimentin were heterogeneously distributed in the cultured SMCs. The predominant distribution of β-NM actin in the cell periphery was associated with densely distributed vinculin plaques and disrupted or striated myosin and α-actinin aggregates, which may reflect a process of stress fiber assembly during cell spreading and focal adhesion formation. The high-level labeling of α-SM actin in the central portion of stress fibers was related to continuous myosin and punctate α-actinin distribution, which may represent the maturation of the fibrillar structures. The findings also suggest that the stress fibers, in which actin and myosin filaments organize into sarcomere-like units with α-actinin-rich dense bodies analogous to Z-lines, are the contractile structures of cultured SMCs that link to the network of vimentin-containing intermediate filaments through the dense bodies and dense plaques.
I
SMCs express four actin isoforms: two smooth muscle (α-SM and γ-SM) and two non-muscle (β-NM and γ-NM) actins. During vascular development, the differentiating SMCs display a switch of isoactin expression from a predominance of non-muscle to smooth muscle isoforms. This is accompanied by a functional change from proliferation and synthesis of extracellular matrix (ECM) to contraction (Thyberg 1996; Campbell and Campbell 1997). Some non-SMCs, such as myofibroblasts during wound repair, also transiently express α-SM actin and acquire the ability to contract (Schmitt–Gräff et al. 1994). This supports the hypothesis that different isoactins are functionally distinct and therefore should be structurally sorted into different subcellular regions or actin-containing structures in the cell.
Studies by Small and coworkers using freshly isolated gizzard smooth muscle strips have demonstrated that β-NM actin is co-localized with intermediate filaments in longitudinal channels that they termed the “cytoskeletal domain” because myosin was absent in these regions. They suggested that at least two different thin filaments exist in SMCs: non-muscle (β- and γ-NM) actin filaments which are contained in the “cytoskeletal domain,” and smooth muscle (γ- and γ-SM) actin filaments, which co-localize with myosin in a so-called “contractile domain” (Small et al. 1986; Draeger et al. 1990; North et al. 1994; Small and North 1995). However, the segregation of the γ-SM actin (chicken gizzard SMCs do not express α-actin) from the cytoskeletal domain was not clearly distinguished in their model because the fibrillar structures were too closely packed in the cytoplasm, even in artificially expanded thin sections (North et al. 1994). The existence of two different populations of actin filaments was also challenged by Drew and co-workers (Drew et al. 1991; Drew and Murphy 1997), who showed that γ-SM and β-NM actins are randomly distributed within native SMC thin filaments isolated from chicken gizzard.
In cultured cells, intracellular filaments are well spread with the flattening of cells on a substrate. This allows the different components to be clearly distinguished by immunolabeling techniques, with individual filaments able to be traced along their whole course and their dynamic changes monitored in living cells. An additional property of cultured SMCs is that the cells spontaneously undergo a phenotypic modulation from a “contractile” to a “synthetic” phenotype (Chamley–Campbell et al. 1979; Campbell and Campbell 1993; Thyberg 1996). They lose their ability to contract, with a concomitant decrease in myofilaments and contractile proteins, over the first few days of primary culture (Owens et al. 1986; Glukhova et al. 1988; Campbell et al. 1989). In a recent study to investigate the changes in three-dimensional architecture of microfilaments associated with SMC phenotypic modulation, we found that actin isoforms are heterogeneously distributed in cultured vascular SMCs, with α-SM actin predominant in the central region of the cytoplasm and β-NM actin in the periphery. Furthermore, the partial disassembly of stress fibers during SMC phenotypic modulation, which may be responsible for the loss of contractile function of the cell, was mainly due to a selective degradation of α-SM actin (Song et al. 1998). These results seem to support Small's hypothesis that the smooth muscle and non-muscle actins are differently distributed in a single SMC and may involve different functions.
In culture, however, cells usually reorganize their cytoskeletal system to fit the unnatural growth conditions. We still do not know what represents the contractile structure in a cultured SMC and how this relates to the cytoskeletal system. Therefore, it is uncertain whether the heterogeneous distribution of the isoactins in different regions in a cultured SMC reflects a special form of subcellular segregation of the contractile and cytoskeletal domains. Therefore, the present study systematically investigated the distribution relationship of the structural proteins in cultured rabbit aortic SMCs using high-resolution confocal laser scanning fluorescent microscopy.
Materials and Methods
Culture of SMCs
SMCs were prepared as previously described (Campbell et al. 1989). Briefly, freshly dissected aortas from 9–12-week-old rabbits were placed in cold Hanks’ balanced salt solution plus 5% fetal calf serum (FCS; PA Biologicals, Sydney, Australia). After removal of the fat and connective tissue under a dissecting microscope, the vessels were cut longitudinally and the endothelial lining was removed by a 20-min digestion with collagenase (LSO-4177; Worthington Biochem, Freehold, NJ). Watchmaker's forceps were then used to strip the inner two thirds of the media in 1-mm-wide horizontal pieces from the outer third of the media plus adventitia. The muscle pieces were dispersed into single cells with collagenase and elastase (LSO-2292; Worthington Biochem). Freshly dispersed cells were plated out into wells of a 24-well plastic culture plate containing 10-mm round glass coverslips, and cultured in medium 199 (CSL; Melbourne, Australia) supplemented with 5% FCS, at 37C in an atmosphere of 5% CO2 in air. Cells were seeded at a density of 0.25 × 105 cells/cm2 and observed at Day 3–4 of primary culture. Previous studies have demonstrated that rabbit aortic SMCs at the above time point of primary culture display a similar phenotype to those in vivo (Campbell et al. 1989; Song et al. 1998).
Immunocytochemistry and Confocal Laser Scanning Fluorescence Microscopy
The following primary monoclonal antibodies (MAbs) were employed in the present study for single or double labeling of contractile and cytoskeletal components in SMCs: anti-α-actin (mouse IgG1, HHF35; DAKO, Corpinteria, CA, at a dilution of 1:150 or mouse IgG2a, 1A4; Sigma, St Louis, MO, 1:400), anti-β-actin (mouse IgG2a, AC-74; Sigma, 1:100 or mouse IgG1, AC-15; Sigma, 1:100), anti-smooth muscle myosin heavy chain (SM-MHC) (mouse IgG1, hSM-V; Sigma; 1:300), anti-vimentin (mouse IgG2a,Vim3B4; DAKO, 1:150 or mouse IgG1, Vim V9; Sigma, 1:200), anti-α-actinin (mouse IgM, BM-75.2; Sigma, 1:200) and anti-vinculin (mouse IgG1, hVIN1; Sigma, 1:200).
Coverslips bearing SMCs were rinsed briefly in PBS, fixed with 4% paraformaldehyde in PBS (pH 7.4) for 10 min, and then permeabilized with 0.1% Triton X-100 in PBS for 5 min at room temperature (RT). For anti-β-actin antibody labeling, cells were fixed and permeabilized with cold methanol for 5 min because the antibody does not react well with aldehyde-fixed tissue. After washing in PBS, the SMCs were treated with 1% BSA in PBS for 30 minutes to block nonspecific binding and then incubated at 4C overnight with a single antibody or a mixture of primary antibodies specific for related contractile or cytoskeletal proteins, followed by a single antibody or a mixture of FITC-labelled anti-mouse IgG1 antibodies (LO-MG1–2; Serotec, Oxford, United Kingdom 1:200) and biotin-conjugated anti-mouse IgG2a (Silenus; Melbourne, Australia 1:200) or anti-mouse IgM (Southern Biotech, Lexington, KY 1:300), with streptavidin–Texas Red (Amersham; Arlington Heights, IL, 1:100) as the third layer. In negative control cultures the primary antibody was replaced by nonimmune mouse IgG1 or IgG2a.
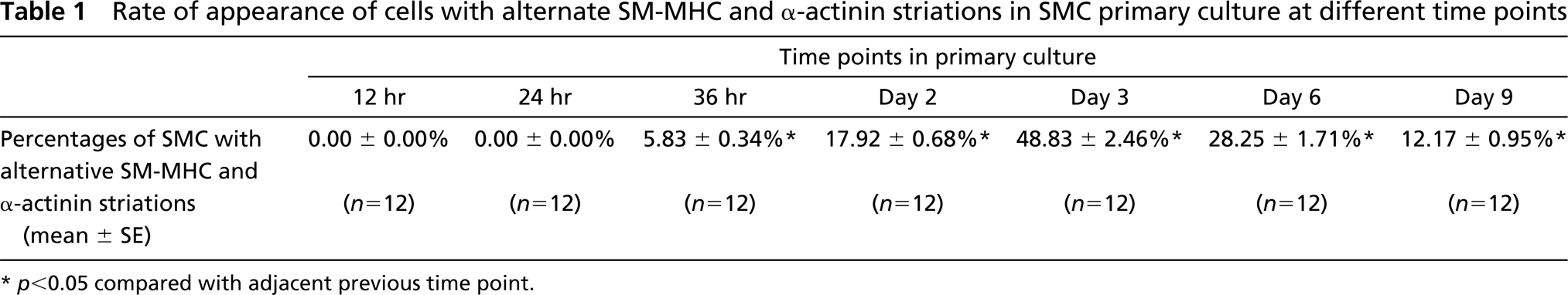
In further experiments, the rate of appearance of SMCs with SM-MHC and α-actinin striations was investigated at different time points (12 hr, 24 hr, 36 hr, Day 2, Day 3, Day 6, and Day 9) of culture. The cells were double labeled with anti-SM-MHC and anti-α-actinin antibodies. To determine the percentage of cells with striated staining patterns, four coverslips were taken randomly from each time point and 50 cells counted on each coverslip under an objective of ×60. This procedure was repeated with three different cultures. The mean percentages of cells with myosin and α-actinin striations were calculated from 12 coverslips from each time point and the data were statistically analyzed with SigmaStat 1.02 statistical software (Jandel; San Rafael, CA), using a one-way ANOVA. To determine whether the formation of SM-MHC and α-actinin striations is an artifact due to inappropriate chemical treatment of the cell, the effects of different fixation and permeabilization protocols on the antibody staining patterns were also compared, including (a) fixing cells with 2% glutaraldehyde in PBS followed by permeabilization of 0.1% Triton X-100 (Hartwig 1992), (b) permeabilizing cells with 0.1% Triton X-100 (for only 30 sec) before fixation with 4% paraformaldehyde or 0.5% glutaraldehyde (Langanger et al. 1986), or (c) without fixation (only 20-min incubation for each round of antibody staining), and (d) fixing cells with either cold acetone or methanol without Triton X-100 permeabilization.
After staining, the coverslips were gently mounted upside down on glass slides with gelvatol mounting medium and observed with a Bio Rad (Hercules, CA) MRC 1024 confocal system, incorporating a krypton/argon laser and an Olympus BX60 microscope. Serial optical sections were collected by scanning the whole thickness of the cell in the Z-direction with a scanning step of 0.4 μm. For double-labeled samples, a sequential scanning method for different channels was selected to avoid the possibility of “bleed-through” between channels. Images were reconstructed using Bio-Rad Laser Sharp.
Results
Relationship of Actin Isoforms to Vinculin and α-actinin
Although both α-SM and β-NM actins was localized on stress fibers in cultured SMCs (Figure 1A), α-SM actin was predominantly distributed on the stress fibers in the central region of the cytoplasm and β-NM actin in the ends of the fibers at the cell periphery. In the spreading lamellae, β-NM actin labeling also appeared in the cytoplasm in a diffuse form that clearly delineated the cell margin (Figure 1A, arrows), whereas α-SM actin was restricted to stress fibers. Higher magnification showed that the switch from a predominance of α-SM actin to β-NM actin in stress fibers (from middle to terminal parts of the fiber) was gradual and continuous (Figure 1B).
Double labeling with anti-vinculin in combination with either anti-α-SM or β-NM actin antibody showed that vinculin was present on the cell membrane in the form of long oval plaques. The plaques were more densely distributed in the periphery of the cell (Figure 2A), where the majority of stress fibers ended. Both α-SM and β-NM actin appeared in vinculin plaques at the two ends of the stress fibers (Figures 2B and 2C), but where the vinculin plaques were concentrated, the stress fibers stained more strongly for β-NM actin (Figure 2C) than α-SM actin (Figure 2B).
α-Actinin was present in a fine punctate form on the stress fibers that were also continuously stained by both anti-α-SM (Figures 3A–3C) and β-NM (Figures 3D–3F) actin antibodies. In the peripheral region of some cells, the punctate α-actinin labeling aligned to form bars or cross-striations (Figures 3A and 3C, arrow). However, in cells fixed with methanol (for labeling in combination with anti-β-actin), this cross-striated labeling was seldom found (Figures 3D and 3F). Enlarged images revealed that the discrete α-actinin spots were randomly distributed on stress fibers (Figures 3C and 3F), but the bar-like and cross-striated labeling showed a periodicity with regular intervals (Figure 3C, arrow). Where the cross-striations of α-actinin were obvious, the α-SM actin staining of stress fibers became weaker and a little diffuse (Figure 3B, arrow).
Relationship of SM-MHC to α-actinin and Actin Isoforms
SM-MHC was present in different forms in individual SMCs. In the central part of the cytoplasm, it distributed continuously on stress fibers. In the peripheral region or leading lamellae, SM-MHC appeared as discrete spots (Figure 4A). Higher magnification showed that the SM-MHC spots were either localized periodically with regular intervals on the terminal branches of stress fibers (Figure 4B), or randomly distributed close to the leading edge (Figure 4C). Some SM-MHC spots aggregated into small bars that crossed the stress fibers (Figure 4C). In some cells, the aggregated SM-MHC bars were well aligned to give the cell a striated appearance (Figure 4D). The average center-to-center distance between the myosin striations was calculated as 0.724 ± 0.023 μm (mean ± SE, n = 50 stained pairs) with a range of 0.658–0.940 μm.

Double labeling of cultured rabbit aortic SMCs for α-SM actin (green) and β-NM actin (red). Arrows (
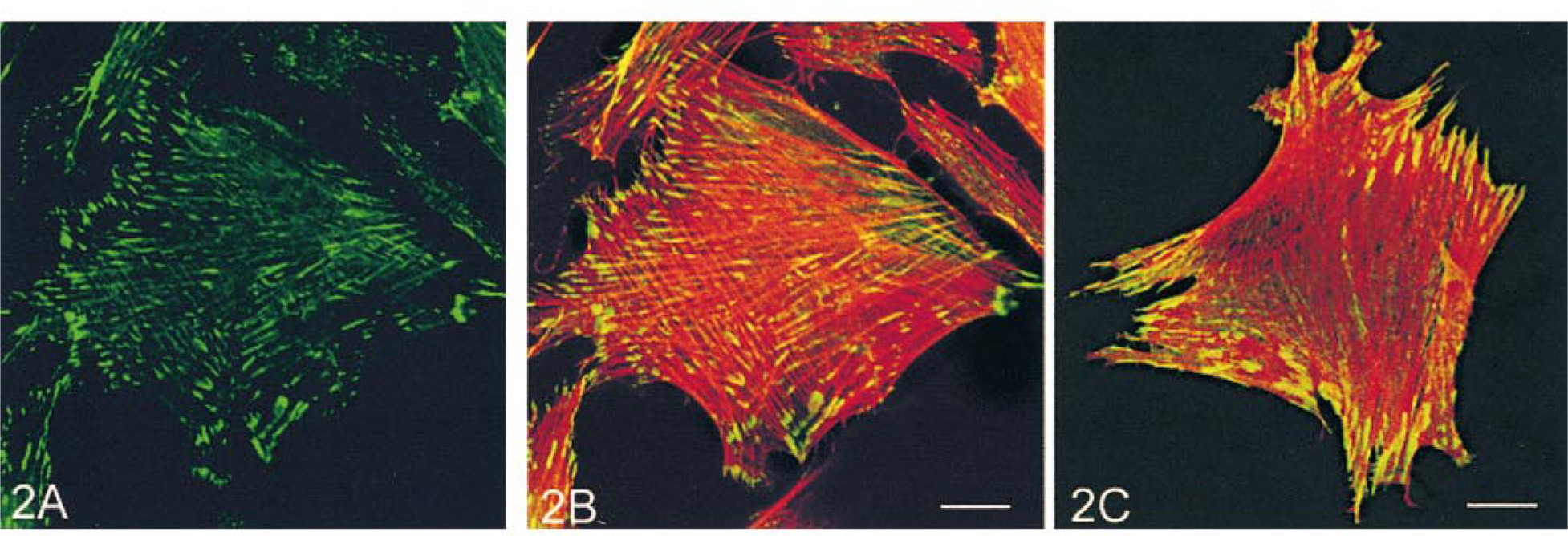
Labeling of cultured rabbit aortic SMCs for vinculin (green) (
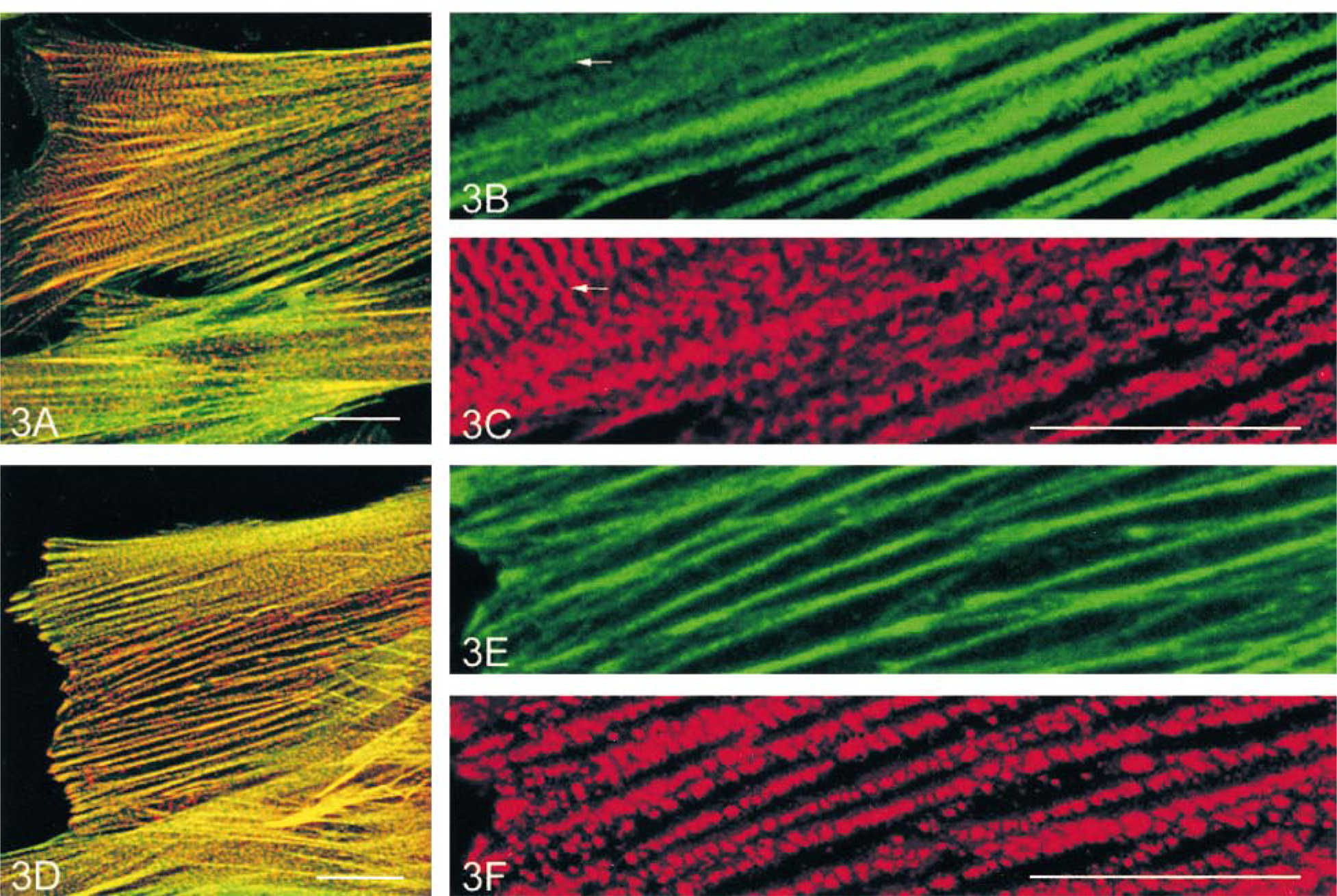
Labeling of cultured rabbit aortic SMCs for α-actinin (red) in combination with α-SM actin (green) (
Double labeling for SM-MHC and α-actinin in cultures showed that in the area of the cell at which SM-MHC displayed periodic staining, α-actinin also formed similar bars or striations (Figure 4E) that alternated between those formed by myosin (Figure 4F), very similar to sarcomeric structures. The average center-to-center distance between the α-actinin striations (0.769 ± 0.014 μm, range 0.658–0.840 μm) was very similar to that of myosin. The periodic distribution of the two proteins disappeared in the central part of the cytoplasm in most of the cells, where the staining of stress fibers for SM-MHC became continuous (Figures 4G and 4H) with punctate α-actinin labeling (Figures 4G and 4I). To investigate the dynamic course of myosin and α-actinin striation formation, cells at different spreading stages were observed. Twelve hours after seeding, almost all cells were still round, with a diffuse staining of SM-MHC and α-actinin throughout the cytoplasm. After 24 hr of culture, a small number of cells began to spread. In these cells the diffuse myosin staining was concentrated in the outer cytoplasm, with sparsely distributed myosin spots appearing in extending leading lamellae (Figure 4J, arrows). α-Actinin staining became punctate throughout the cytoplasm but was more densely distributed in newly formed stress fibers that aligned radially at the cell periphery (Figure 4J). After 36 hr, increased myosin aggregates appeared in the leading lamellae as cells became flattened. At this stage, the alternate myosin and α-actinin striations began to appear at the cell periphery (Figure 4K) in about 5% of cells. By Day 2, as cells became more elongated, the myosin aggregates gradually aligned into small bars and then well-defined striations alternated with α-actinin from the leading edge toward the inner cytoplasm (Figure 4L). The percentages of the cells with SM-MHC and α-actinin striations at different time points of culture and in the cultures treated with various fixation and permeabilization protocols are shown in Table 1 and Table 2, respectively.
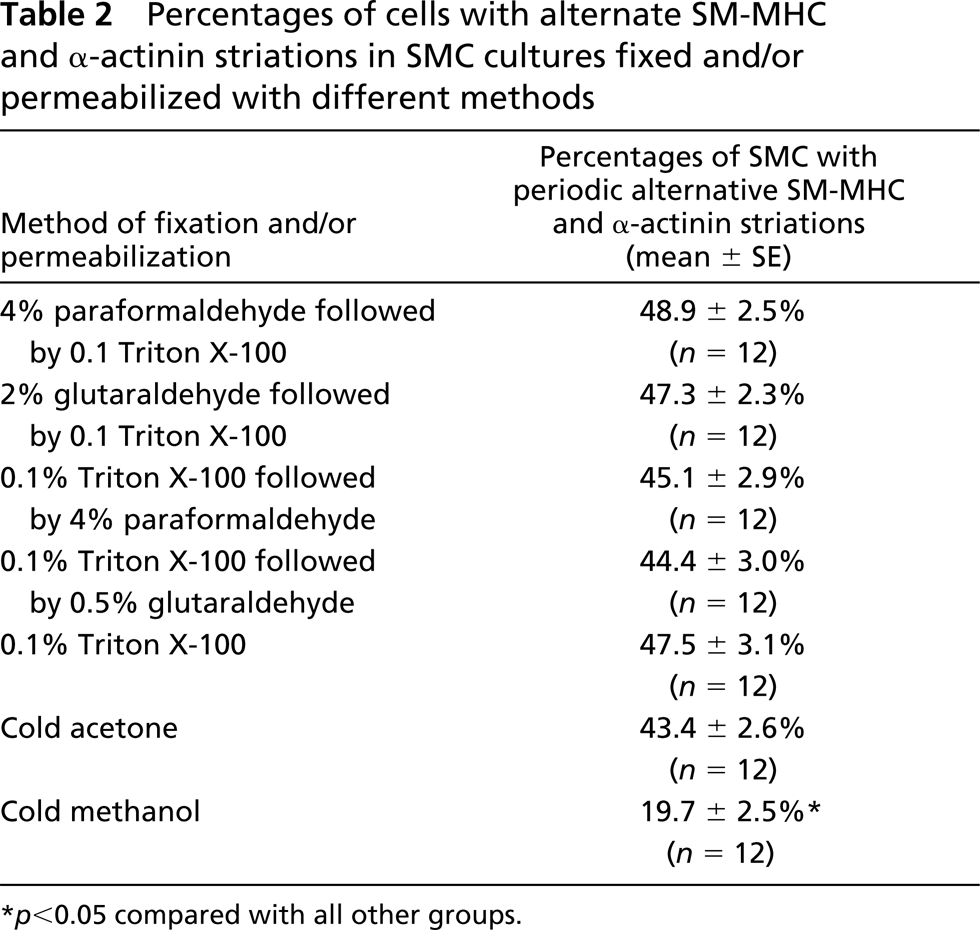
Double labeling of SMCs with anti-SM-MHC in combination with either anti-α-SM or anti-β-NM actin antibody revealed that myosin co-localized with both α-SM and β-NM actins on the stress fibers in the central part of the cytoplasm. However, in the periphery, especially the leading lamellae of the cell where there was an obvious decrease in α-actin staining (Figure 5A), myosin labeling became interrupted along the stress fibers or displayed cross-striations (Figures 5B and 5C).
Relationship of Vimentin to Actin Isoforms, α-actinin, and Vinculin
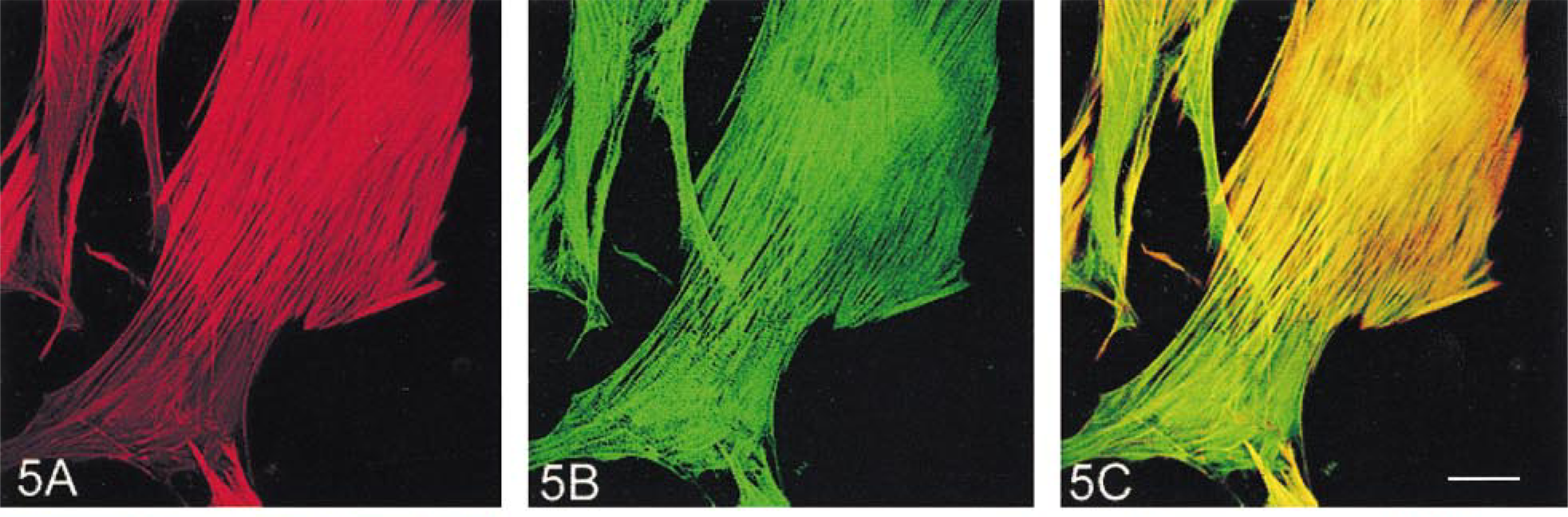
Vimentin-positive intermediate filaments were distributed as a meshed network throughout the cytoplasm, but with greatest concentration in the perinuclear region (Figure 6A). Staining of vimentin, in combination with either anti-α-SM or anti-β-NM actins, showed that the actin-containing stress fibers extended through the network until they reached the periphery of the cell, where the vimentin filaments became sparsely distributed (Figure 6B). High magnification showed that, towards the periphery of the cytoplasm, the vimentin filaments tended to become longitudinally and rather tortuously oriented and wrapped around the actin-containing stress fibers (Figure 6C). There were no apparent differences between spatial relationships of vimentin and the two actin isoforms (Figures 6C and 6D).
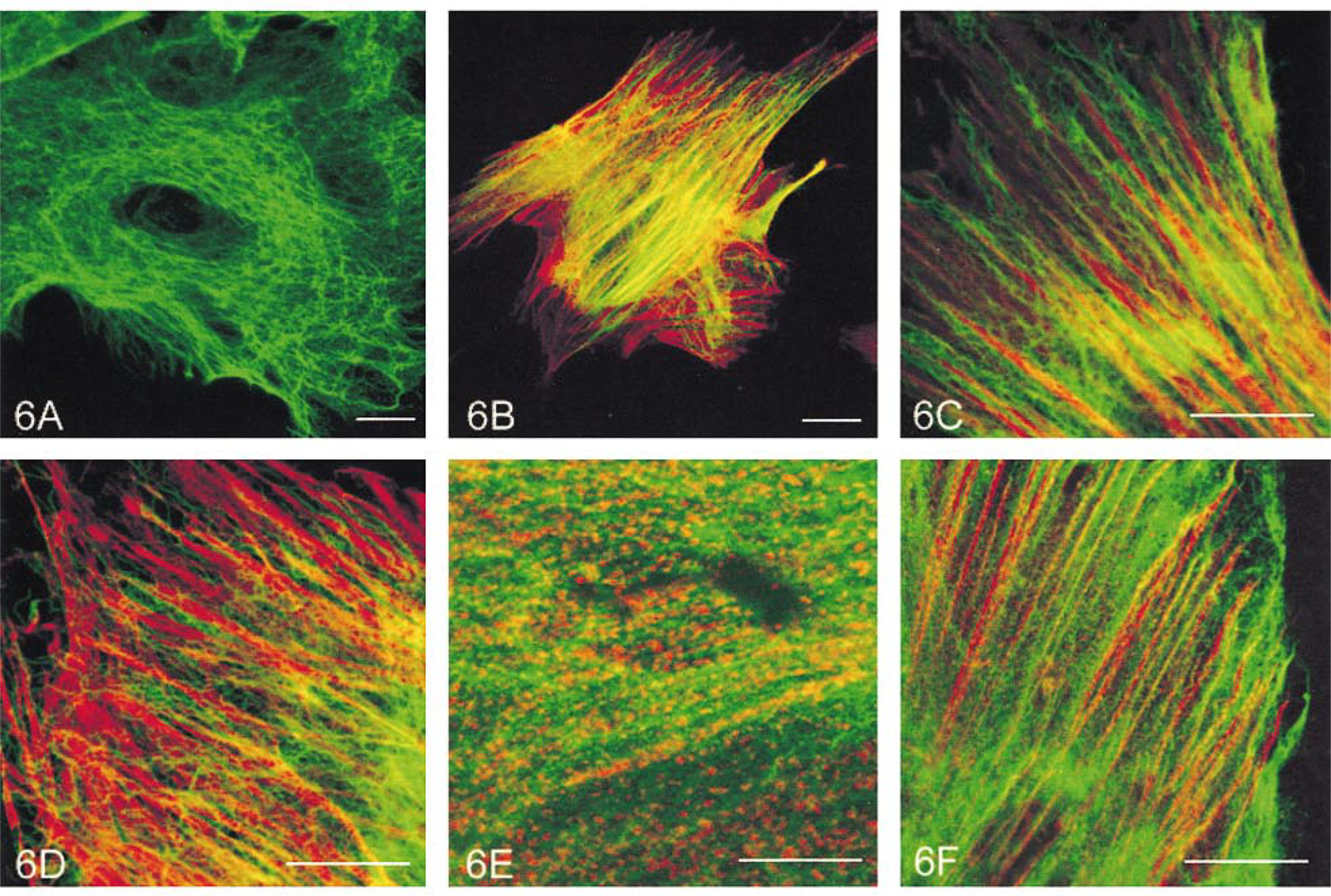
Double staining for vimentin in combination with either anti-α-actinin or anti-vinculin antibodies showed that vimentin labeling did not appear in α-actinin-containing dense bodies in either the central (Figure 6E) or peripheral (Figure 6F) regions of the cell, nor in vinculin-containing membrane-associated dense plaques (Figure 7A).
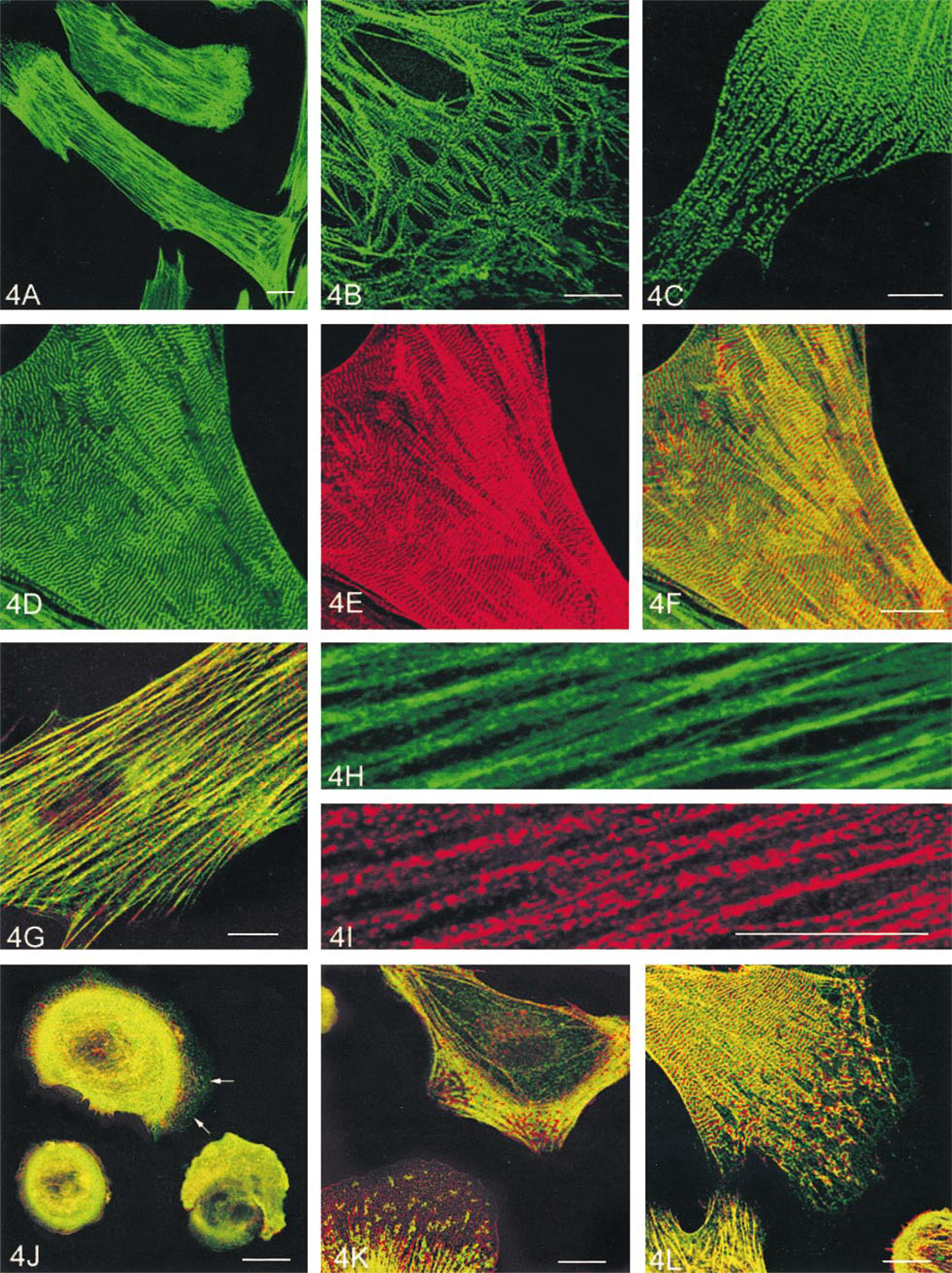
Labeling of cultured rabbit aortic SMCs for SM-MHC (
Rate of appearance of cells with alternate SM-MHC and α-actinin striations in SMC primary culture at different time points

∗ p < 0.05 compared with adjacent previous time point.
Relationship Between α-actinin and Vinculin
In the SMCs double labeled with anti-α-actinin and anti-vinculin antibodies, the α-actinin, whether present in punctate or striated form, appeared not to exist in vinculin-containing plaques (Figures 7B and 7C).
The main distribution properties of the contractile and cytoskeletal proteins in cultured rabbit aortic SMCs are summarized in Table 3.
Discussion
Consistent with our previous report (Song et al. 1998), this study showed that whereas both α-SM and β-NM actin were located on stress fibers in cultured vascular SMCs, the two isoactins were heterogeneously distributed within the cell. α-SM actin was predominant in the middle portions of the stress fibers that occupied the central region of the cytoplasm and β-NM actin in the terminal parts of the fibers usually ending at the cell periphery. Anti-β-NM actin antibody strongly labeled not only the terminal parts of stress fibers but also the peripheral cytoplasm, suggesting that the peripheral distribution of the isoactin also involves a β-NM actin monomer pool or randomly oriented actin filament networks that may not be recognized by confocal microscopy. Double labeling of vinculin in combination with either α-SM or β-NM actin showed that vinculin-positive plaques colocalized with both α-SM and β-NM actins at the ends of stress fibers, indicating that if two kinds of thin (α-SM and β-NM) filaments exist in an SMC, both of them enter the membrane-associated dense plaques. However, the stronger labelling of β-NM actin near the ends of stress fibers and in the area where the vinculin plaques were densely distributed indicates a closer relationship of β-NM actin with focal adhesion plaques.
Percentages of cells with alternate SM-MHC and α-actinin striations in SMC cultures fixed and/or permeabilized with different methods
∗ p < 0.05 compared with all other groups.
Intracellular sorting and functional diversity of isoactins have been demonstrated by recent molecular genetic studies in both muscle and non-muscle cells (Gunning et al. 1997; Schedlich et al. 1997). For example, transfected muscle and non-muscle actins are differently sorted by cultured SMCs and non-muscle cells, with muscle actins incorporated into stress fibers and non-muscle actin into cytoplasm (Mounier et al. 1997). Transfection of the β-NM actin gene into myoblasts promotes cell spreading (Schevzov et al. 1992) and treatment of the cells with antisense oligonucleotides that inhibit peripheral localization of β-NM actin mRNA results in a dramatic change in lamellipodial structures (Kislauskis et al. 1994). It was also found that injection of anti-α-SM actin antibody into fibroblasts inhibits collagen gel contraction by the cells but does not affect total actin assembly and cell spreading (Arora and McCulloch 1994). In chicken gizzard SMCs, β-NM actin was localized in longitudinal channels complementary to myosin distribution regions, but whether γ-SM actin is segregated from these channels was uncertain (North et al. 1994). The results of the present study show that, in cultured SMCs, α-SM and β-NM actins are not completely separated but are predominantly distributed in different regions. The predominant peripheral concentration of β-NM actin and its close relationship to vinculin plaques is consistent with a role of the isoactin in cell adhesion and spreading, a universal cytoskeletal function of anchorage-dependent growth cells. Enlarged images reconstructed from the whole thickness of the SMCs showed that the switch from α-SM actin-predominant to β-NM actin-concentrated regions on stress fibers was gradual and continuous. At the confocal microscopic level, we could not find any defined fibers either positive only for α-SM or only for β-NM actin. However, we do not exclude the possibility that the diffuse cytoplasmic staining of β-NM actin at the cell periphery may represent randomly distributed individual β-actin filaments. Attempts to examine individual thin filaments isolated from the cells using immunoelectron microscopy have been unsuccessful because of high backgrounds, probably due to the unavoidable presence of unpolymerized actin monomers in the filament suspension. Therefore, we are not certain whether this different distribution of actin isoforms is due to a non-random assembly of different thin filaments into stress fibers or to a heterogeneous co-polymerization of the different isoactins into individual thin filaments.
Double labeling of cultured rabbit aortic SMCs for α-SM actin (red) (
Labeling of cultured rabbit aortic SMCs for vimentin (
Double labeling of cultured rabbit aortic SMCs for vinculin (green) in combination with vimentin (red) (
Although the heterogeneous distributions of α-SM and β-NM actins in cultured SMCs suggest different functions for the isoactins, the question remains as to whether this represents the subcellular segregation of contractile and cytoskeletal components in SMCs under culture conditions. If this is so, myosin presumably interacts preferentially with α-SM actin for contractile force generation and therefore should also be predominantly distributed in the central part of the cytoplasm. The vimentin-containing intermediate filaments should have a closer relationship with β-NM actin. However, the results of the present study reveal a complex distribution relationship for these related components.
In cultured SMCs, SM-MHC, like actins, was located mainly in stress fibers, but with distinct staining patterns observed in different subcellular regions. In the central part of the cytoplasm, it displayed strong and continuous labeling that co-localized with actin in stress fibers. In the periphery of the cell, especially in the leading lamellae, the myosin labeling either became periodically disrupted on the terminal branches of the stress fibers or aggregated in spots or bars that did not exactly coincide with the fibers. In some cells the myosin bars were well aligned, giving the cell a striated appearance. Significantly, in areas at which myosin displayed periodic labeling, α-actinin formed similar spots, bars, or striations that alternated with those of myosin, very similar to the sarcomeric structures in striated muscles. In a few cells, the periodically alternating myosin and α-actinin striations even appeared in whole cytoplasm. Several investigations have previously reported the striated staining pattern of myosin in isolated (Bagby and Pepe 1978) or cultured SMC (Gröschel–Stewart et al. 1975a,b), but it was dismissed as an artifact caused by inappropriate fixation or permeabilization of the cell (Gabella 1984).
To investigate this possibility, we tested several commonly used fixation and permeabilization protocols, including detergent permeabilization with or without aldehyde fixation and fixation with organic solvents (cold acetone or methanol) without detergent permeabilization. The results showed that the cells with periodically alternating myosin and α-actinin striations appeared in a similar frequency with all protocols except for methanol fixation, in which the frequency of the phenomenon was much less. It has been accepted that aldehyde fixation of cells in physiological conditions gives an optimal preservation of myosin-containing structures (Somlyo and Franzini–Armstrong 1985). A recent study also showed that the procedure of routine fixation and permeabilization of SMCs does not adversely affect the myosin filament structure (including the crossbridges) at the ultrastructural level (Hodgkinson et al. 1995). This, together with our findings, leads us to conclude that the periodically alternating myosin and α-actinin staining may represent myosin reorganization in cultured SMCs rather than an artifact caused by chemical fixation and permeabilization. The mechanism for this phenomenon is unknown. The alternate myosin and α-actinin spots, bars, and striations do not appear to represent mature and stable contractile structures, because they frequently appeared in the leading lamellae where α-SM actin labeling was obviously decreased and they did not exactly match the stress fibers. We also found that the active formation of the striations took place during cell spreading and that, after cells reached confluency, the phenomenon was less frequently observed and the striations were limited to very small regions at one or both ends of the cell. It is known that stress fibers are in a dynamic assembly–disassembly state and that the assembly process takes place mainly in the submembrane areas at the cell periphery (Bockus and Stile 1984; Bray and White 1988; Giuliano and Taylor 1990). A study by Verkhovsky et al. (1995) found that, when fluorescence-labeled myosin II was micro-injected into living fibroblasts, the myosin spots first appeared near the leading edge and then aligned to form myosin ribbons that were gradually integrated into loosely packed stress fibers. The myosin ribbons described by these authors are very similar to the myosin bars and striations found in the present study, and it is also known that these structures are formed by side to side-connected myosin filaments (Ashton et al. 1975; Verkhovsky et al. 1995). Thus, the appearance of myosin spots, bars, and striations in the periphery of cultured SMCs may reflect an ongoing process in which myosin filaments assemble into actin-containing stress fibers during the spreading of the cell. On the other hand, the periodic myosin and α-actinin striations indicate that in SMCs the contractile filaments are also able to form sarcomere-like units with α-actinin-containing dense bodies analoguous to Z-lines (Ashton et al. 1975). The disappearance of this periodic staining pattern in the central parts of the stress fibers (where myosin staining becomes continuous and the punctate α-actinin labeling is randomly distributed) may be due to the staggering of the regular filament order in the large and numerous filament-containing bundles. A similar periodic distribution of either α-actinin or myosin in stress fibers was previously described in some cultured non-muscle cell types (Gordon 1978; Langanger et al. 1986; Svitkina et al. 1989) and was suggested to be an organization of sarcomere-like units. In these cells the size of the sarcomere-like units (central-to-central distance of stained segments) varies from 0.20–0.28 μm (Svitkina et al. 1989) to 1.1–1.5 μm (Gordon 1978), and neither α-actinin nor myosin was found to be registered into striations. In the present study, the center-to-center distance of myosin or α-actinin striations in cultured SMCs is about 0.658–0.940 μm, much less than the size of the sarcomere in striated muscles (2–3 μm). This suggests that myosin molecules may assemble into thick filaments with different lengths in different cell types.
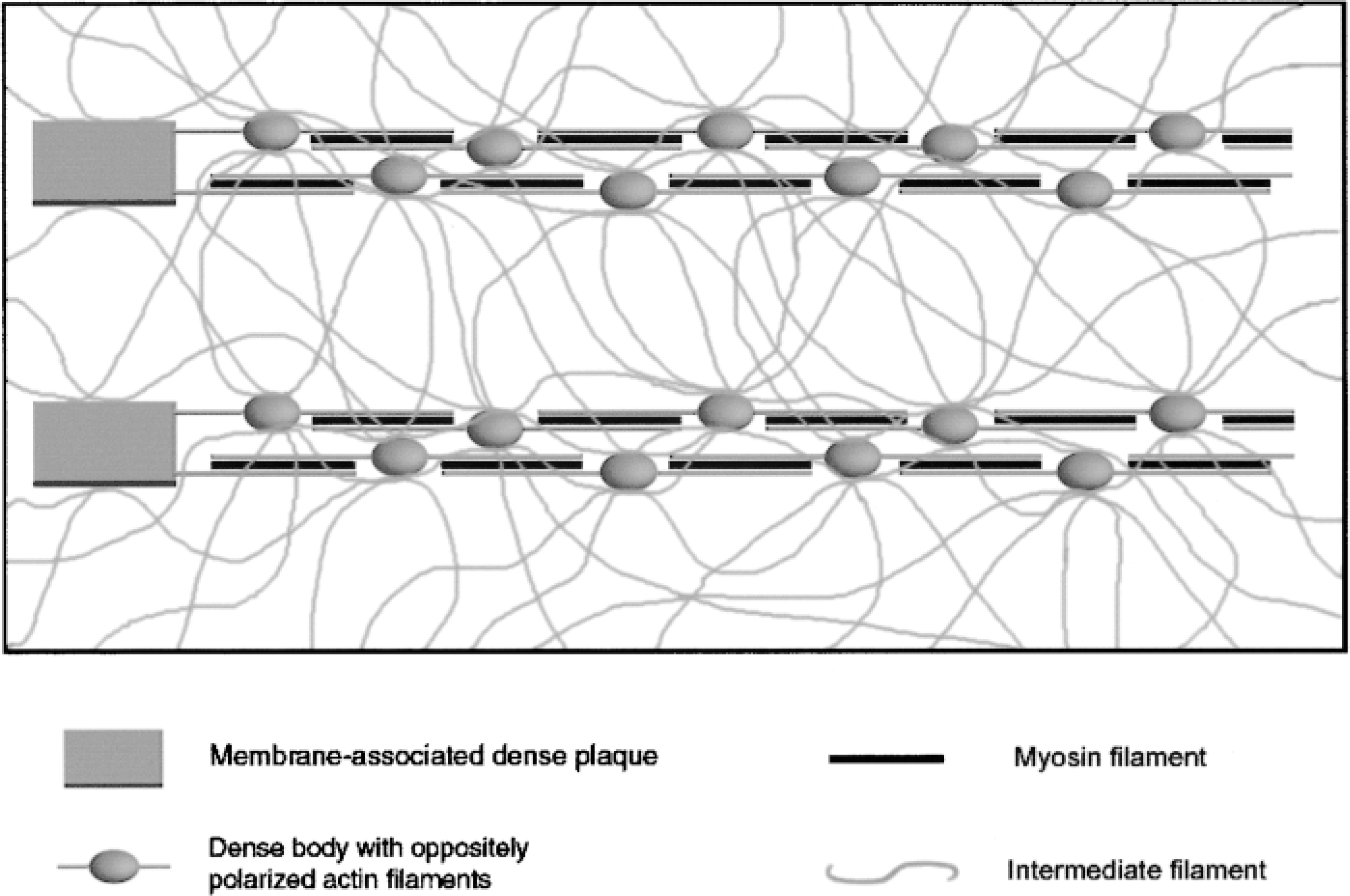
Schematic illustration of the contractile and cytoskeletal filament organization in cultured SMCs.
Localization or distribution patterns of contractile and cytoskeletal proteins in different subcellular regions in cultured rabbit aortic SMCs
As demonstrated by anti-vimentin antibody staining, the intermediate filaments in cultured SMCs form a meshed network throughout the cytoplasm. The filaments were more densely distributed in the perinuclear region, where the stress fibers were strongly stained for α-SM actin. Towards the periphery of the cell, the randomly oriented intermediate filaments gradually became parallel to the stress fibers but, contrary to our expectations, there was no evidence to indicate a closer relationship with β-NM actin.
The distribution properties of the contractile and cytoskeletal proteins revealed in the present study (as summarized in Table 3) suggest that the predominant distributions of α-SM and β-NM actins in different regions in cultured SMCs do not indicate a subcellular sorting of the contractile and cytoskeletal components into separate domains. More likely, the peripheral concentration of β-NM actin, together with densely distributed vinculin plaques and periodically disrupted or striated myosin–α-actinin aggregates, reflects a process of stress fiber assembly during cell spreading and focal adhesion formation. The high-level labeling of α-SM actin with continuous myosin and punctate α-actinin distributions along the central portions of stress fibers represents the maturation of the fibrillar structures.
What is the contractile structure in a cultured SMC, and how is it related to the cytoskeleton? Our previous study demonstrated an overall three-dimensional arrangement of the filamentous system in cultured vascular SMCs. The actin- and myosin-containing myofilaments constitute large filament bundles (stress fibers) that are interconnected and supported by a meshed network of intermediate filaments (Song et al. 1998). The present observations further reveal that α-actinin-rich cytoplasmic dense bodies also localize in a punctate form in the stress fibers, with vinculin-containing membrane-associated dense plaques at the two ends of the fibers. This indicates that the stress fiber is the contractile structure of SMCs when grown in culture. Figure 8 presents a schematic illustration of the contractile and cytoskeletal filament organization in cultured SMCs. Dense bodies couple thin (including both α-SM and β-NM actin) filaments into linear arrays that are tightly packed in large stress fibers and finally attach to the membrane-associated dense plaques. The side-polar SM–myosin filaments (Hodgkinson et al. 1995; Xu et al. 1996) are also integrated in this array and may be located between two oppositely polarized thin filaments from different dense bodies but the periodic order of dense bodies and of myosin and actin filaments is staggered in the large stress fibers. The intermediate filament network interconnects the contractile structures by linking dense bodies in different stress fibers and dense plaques on the cytoplasmic surface of the cell membrane. The present study showed that vimentin did not co-localize with either α-actinin or vinculin. This is consistent with previous investigations demonstrating that intermediate filaments do not enter the dense bodies or dense plaques but attach to their surfaces (Bond and Somlyo 1982; Tsukita et al. 1983).
Footnotes
Acknowledgements
Supported by the Heart Foundation of Australia.