
Abstract
Digastric muscle (DGM) is a powerful jaw-opening muscle that participates in chewing, swallowing, breathing, and speech. For better understanding of its contractile properties, five pairs of adult human DGMs were obtained from autopsies and processed with immunocytochemistry and/or immunoblotting. Monoclonal antibodies against α-cardiac, slow tonic, neonatal, and embryonic myosin heavy chain (MHC) isoforms were employed to determine whether the DGM fibers contain these MHC isoforms, which have previously been demonstrated in restricted specialized craniocervical skeletal muscles but have not been reported in normal adult human trunk and limb muscles. The results showed expression of all these MHC isoforms in adult human DGMs. About half of the fibers reacted positively to the antibody specific for the α-cardiac MHC isoform in DGMs, and the number of these fibers decreased with age. Slow tonic MHC isoform containing fibers accounted for 19% of the total fiber population. Both the α-cardiac and slow tonic MHC isoforms were found to coexist mainly with the slow twitch MHC isoform in a fiber. A few DGM fibers expressed the embryonic or neonatal MHC isoform. The findings suggest that human DGM fibers may be specialized to facilitate performance of complex motor behaviors in the upper airway and digestive tract.
Keywords
T
It is widely accepted that contractile properties of a given skeletal muscle are determined mainly by myosin heavy chain (MHC) isoforms expressed by muscle fibers. In addition to the major MHC isoforms (i.e., types I, IIa, IIx, and IIb), some rare MHC isoforms have been found in certain specialized muscles. For example, slow tonic MHC isoform (MHC-ton)-containing muscle fibers (ton-mfs) are rare in mammals but are commonly present in the skeletal muscles of amphibians (Kuffler and Williams 1953; Pliszka and Strzelecka-Golaszewska 1981) and birds (Ovalle 1978; Simpson 1979). However, MHC-ton has been found in the extrafusal muscle fibers of human extraocular (Pierobon-Bormioli et al. 1979), laryngeal (Han et al. 1999a; Sanders et al. 2000a), and tongue (Sanders et al. 2000b) muscles. Another unique isoform is the α-cardiac MHC isoform (MHC-α), which was thought to exist almost exclusively in the heart. However, MHC-α-containing muscle fibers (α-mfs) have also been found in the extraocular muscles of humans (Pedrosa-Domellöf et al. 1992) and animals (Rushbrook et al. 1994; Rubinstein and Hob 2000), the masticatory muscles of humans (Bredman et al. 1991; Pedrosa-Domellöf et al. 1992; Sciote et al. 1994; Stal et al. 1994) and some animals (Bredman et al. 1991; d'Albis et al. 1993), and the human tongue (Sanders et al. 2000b) and laryngeal muscles (Han et al. 1999b). During development, MHC-α is also transiently expressed in the mammalian body skeletal muscles (Lefaucheur et al. 1997). The MHC-ton (Eriksson et al. 1994; Liu et al. 2002) and MHC-α (Pedrosa et al. 1990; Kucera et al. 1992; McWhorter et al. 1995; Liu et al. 2003) were also found in the intrafusal fibers of muscle spindles in humans and other mammals.
The fiber type composition of the DGM has been studied by using cytochemistry, immunocytochemistry, electrophoresis, and immunoblotting techniques in humans (Eriksson et al. 1982; Bredman et al. 1991; Korfage et al. 2000; Monemi et al. 2000), primates (Rowlerson et al. 1983; Miller and Farias 1988; Andreo et al. 1994), and other mammals (Rowlerson et al. 1983; Rokx et al. 1984; Bubb and Sims 1986; Lev-Tov and Tal 1987; Bredman et al. 1991; Cobos et al. 2001). The majority of studies demonstrated that both bellies of the human DGM showed a fiber composition similar to that of normal limb and trunk muscles. The proportion of each of the main fiber types (i.e., types I, IIA, and IIX/B) in the DGM appears to vary with the techniques used and the cases selected (Eriksson et al. 1982; Monemi et al. 2000; Korfage and Van Eijden 2003). The rare IIM fiber type that seems to be associated with an aggressive bite was not found in the human DGM (Rowlerson et al. 1983). The previous investigators reported that MHC-α was lacking (Bredman et al. 1991; Monemi et al. 2000) or expressed in only a few fibers (Korfage et al. 2000) of the human DGM. To date, MHC-ton has not been reported in the human DGM.
It is hypothesized that the wide range of physiological behaviors of the DGM may be correlated with the content of MHC isoforms expressed by the fibers of this muscle. The present study was designed to examine whether adult DGM fibers contain some unique (i.e., α-cardiac and slow tonic) and developmental (i.e., embryonic and neonatal) MHC isoforms.
Specificities of the monoclonal antibodies used

Department of Cell Biology and Anatomy, Cornell Medical College, New York, NY.
Materials and Methods
Tissue Preparation
Ten adult human DGM specimens were obtained from five autopsy cases (three males and two females, mean age 56 years). The muscles were obtained within 24 hr post mortem. These individuals were without known systemic neuromuscular disorders. The anterior and posterior bellies of the DGMs were exposed and removed together. The muscle samples (∼6 mm in length) were taken from the middle portions of both bellies of all left DGMs and two right DGMs. All muscle blocks were placed in small aluminum foil boats, embedded in OCT compound (Tissue-Tek; Miles, Elkhart, IN), and frozen in 2-methylbutane cooled by direct immersion of CO2 dry ice. For comparison, a piece of human masseter (as positive control) and a piece of biceps brachii (as negative control) were placed beside each DGM muscle block and frozen in the same boat. Serial transverse 10-μm sections were cut at −25C by using a cryocut 1800 microtome (Reichert–Jung; Mannheim, Germany). For immunocytochemistry, the serial cross-sections were reacted with a panel of type- or isoform-specific monoclonal antibodies (MAbs). In addition, small muscle fascicles taken from the middle portions of three right DGMa and DGMp were prepared for electrophoretic immunoblotting. Small muscle fascicles taken from two human masseters and a biceps brachii were also used for this technique as controls.
Immunocytochemistry
Monoclonal Antibodies. The serial cross-sections used for immunocytochemistry were incubated with a panel of type-specific anti-MHC MAbs against major (I, II, and IIa), unique (MHC-ton and MHC-α), and developmental (embryonic and neonatal IIa) MHC isoforms (Table 1).
Immunofluorescence. The immunofluorescence was performed according to our previous description (Han et al. 1999a). The sections were incubated at 4C in the hybridoma tissue culture supernatant of MAbs BA-G5 and ALD-58 for 24 hr and in N1.551 and F1.652 for 12 hr.
Immunoperoxidase. The biotin–avidin–immunoperoxidase procedure was performed according to the instructions for the Vectastain ABC kit (Vector Labs; Burlingame, CA), with some modification. The frozen sections were fixed in 4% paraformaldehyde for 10 min, blocked in 1% BSA with 0.1% Triton X-100 for 30 min, and incubated in MAb NOQ7.5.4D for 1 hr at room temperature and in MAbs MY-32 and SC-71 overnight at 4C. Then the sections were incubated in diluted biotinylated secondary antibody for 1 hr and in ABC reagent for 1 hr. The peroxidase substrate kit–DAB was used to visualize the specific binding sites of the different MAbs.
To examine the co-expression of different MHC isoforms in individual muscle fibers, a combination of antibodies was used to stain the same fibers in serial sections of the DGMa. Fiber type specificity was determined by direct comparison of individual fibers in serial sections reacted with different MAbs and visualized by immunofluorescence (BA-G5 and ALD-58) and immunoperoxidase (NOQ7.5.4D, MY-32, and SC-71).
All the sections were examined with a Zeiss Axiophot-2 universal microscope equipped with epifluorescence and DIC (differential interference contrast). The images were photographed with a Spot-32 digital camera (Diagnostic Instruments; Keene, NH) attached to the microscope and connected with a computer. Fiber quantification was carried out using a SigmaScan program (Jandel Scientific, Jandel Corporation, Point Richmond, CA).
Electrophoretic Immunoblotting
Sample Preparation. The existence of α-cardiac, slow tonic, neonatal IIa, and embryonic MHC isoforms was confirmed by electrophoretic immunoblotting in both anterior and posterior bellies of three human DGMs. The samples were prepared and further processed according to our previous description (Han et al. 1999a), with some modification. Small muscle fascicles were minced with single-edge blades and ground with a small electric grinder in a centrifuge tube. The ground tissue was then suspended in 0.1 M Tris buffer containing 1 mM EDTA to form a crude homogenate. The crude homogenate was spun at 800 rpm for 10 sec to remove large tissue debris. The supernatant was removed and mixed with the sample buffer (Hames 1990) in a 2:1 ratio. Before electrophoresis, the samples were heated in the boiling water bath for 4 min and then stored at −20C. The protein concentration was assayed by using an Ultrospec 2000 Spectrophotometer (Pharmacia Biotech; Piscataway, NJ). Ten to 30 μg protein was loaded to each well of the gel.
Electrophoresis. One-dimensional electrophoresis was performed on Nu PAGE 4–12% Bis-Tris gradient gels at 100 V in a 4C refrigerator for 2 hr. Some of the gels were stained with Coomassie Blue and the others were used for immunoblotting.
Immunoblotting. The proteins displayed by electrophoresis were transferred to an Immobilon-P PVDF membrane (Millipore; Bedford, MA). After transfer, the membrane with target protein was immersed in a 5% BSA blocking solution, incubated overnight in 1:500 diluted supernatant of the first MAb, and then incubated for 1 hr in 1:1000 diluted HRP-conjugated goat anti-mouse IgG (Amersham; Arlington Heights, IL) at RT. The immunoblot was developed with ECL (enhanced chemiluminescence) detection reagents (Amersham) according to Durrant and Fowler (1994). The chemiluminescent signal was captured by Polaroid film using an ECL mini camera (Amersham).
Results
Immunofluorescent Microscopy
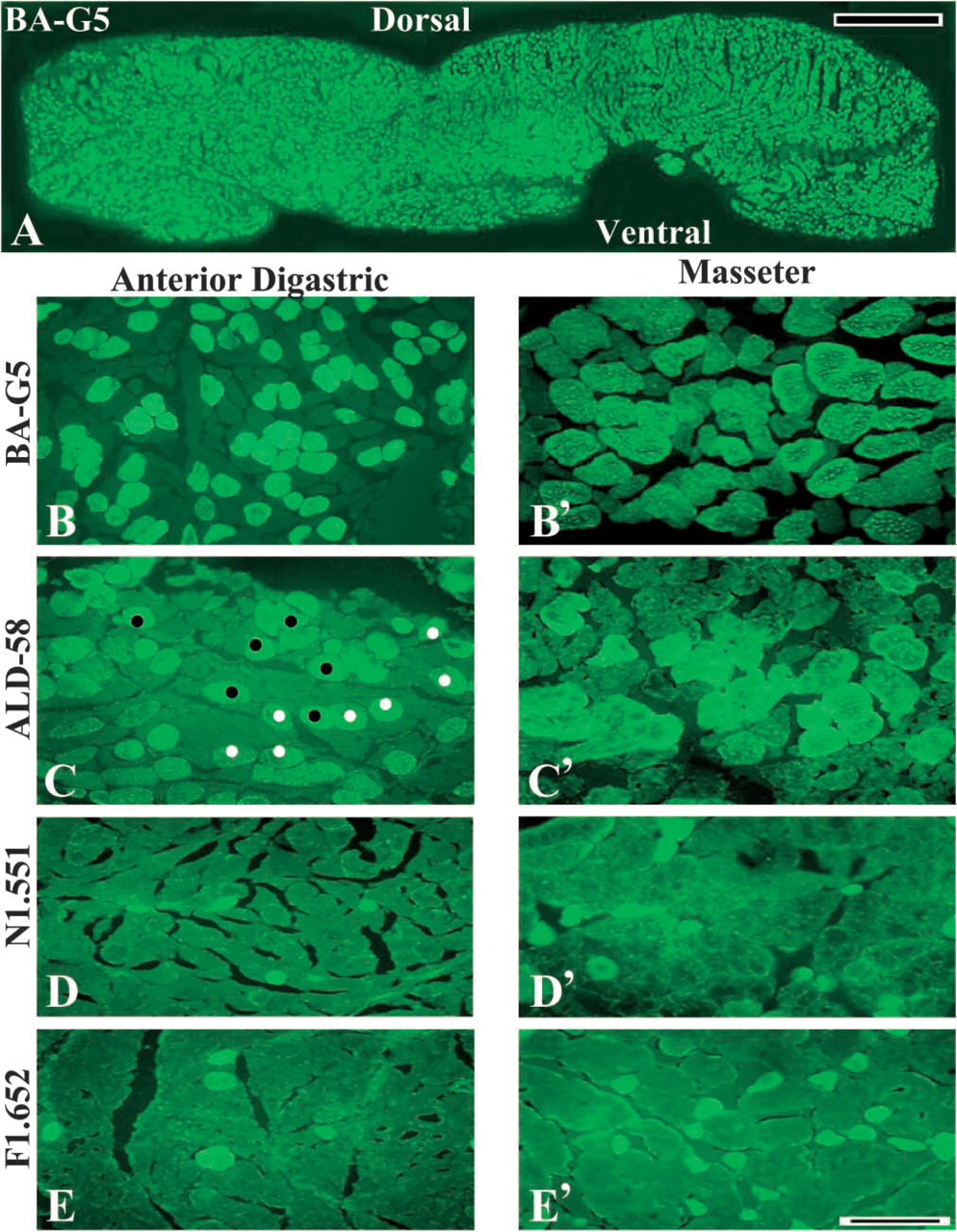
Reaction with MAb BA-G5. Both the anterior and posterior bellies of the DGM contained many muscle fibers that reacted positively to MAb BA-G5 specific for MHC-α (Figures 1A and 1B). Most of these fibers appeared singly, though some of them were clustered in small fascicles containing several fibers (Figure 1B). The α-mfs were intermingled with muscle fibers that did not react to MAb BA-G5 (Figures 1B and 2A). The DGMa showed that the proportions of α-mfs varied with muscle regions and age in all samples examined. The average percentage of α-mfs was higher in the dorsal part (54.52%) than in the ventral part (48.10%) of the muscle (p<0.05) (Figure 3). In the DGMa, a relatively lower percentage of α-mfs was found in aged muscles (35.04%) compared with adult muscles (51.77%; Figure 2E). The difference was highly significant (p<0.01). In the DGMp, almost half of the muscle fibers reacted to MAb BA-G5 and no significant difference in the fiber-type distribution was found between muscle regions and ages (p>0.05).
Fiber-to-fiber comparisons of the serial cross-sections from the DGMa showed that MHC-α usually coexisted with the slow twitch MHC isoform. In this belly, ∼98% of the α-mfs also reacted positively to MAb NOQ7.5.4D (Figures 2A and 2B). The remaining 2% reacted with both NOQ7.5.4D and SC-71. In other words, the MHC-α coexisted mostly with slow I MHC to form I/α hybrid fibers, but rarely with both slow type I and fast type IIa MHC to form I/α/IIa hybrid fibers, and none only with fast type II MHC isoform (Figures 2A–2D).
Reaction with MAb ALD-58. A number of muscle fibers in both bellies of the DGMs reacted strongly to MAb ALD-58. These fibers were assumed to be tonmfs (Shafiq et al. 1984). As with α-mfs, the ton-mfs occurred singly or in fascicles of three to five fibers (Figure 1C). In the DGMa there were more ton-mfs in the peripheral region than in the center of the muscle. Specifically, the dorsal part of the DGMa contained more ton-mfs (22.90%) compared with the 15.5% of the ventral part (p<0.05) (Figure 3). Some ton-mfs were found in the DGMp but no regional difference was observed in this belly.
In the DGMs, the average diameter of the ton-mfs (48.63 ± 5.84 μm) was larger than that of the muscle fibers that did not react to MAb ALD-58 (28.72 ± 8.23 μm) (p<0.01) (Figure 1C). No significant age-related differences in the concentration of ton-mfs were observed. Fiber-to-fiber comparison in the serial cross-sections incubated with various MAbs revealed that MHC-ton usually coexisted with the slow type I MHC isoform (result not shown).
Immunofluorescence of muscle fibers in adult human anterior digastric muscle. (
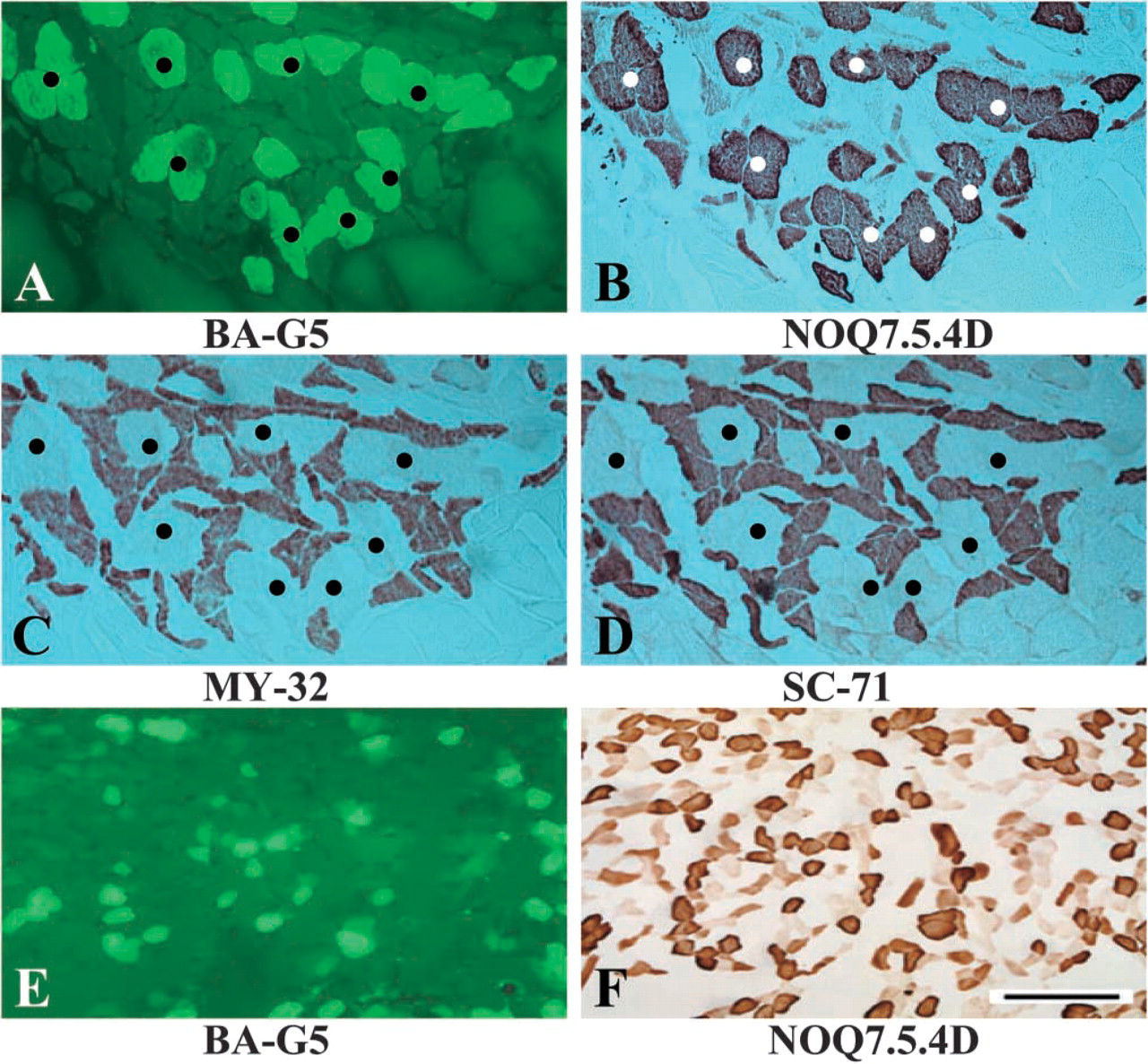
Immunocytochemistry of serial cross-sections of an adult (
Reactions with MAbs N1.551 and F1.652. In the human adult DGMa, 3.2% of the muscle fibers reacted positively to MAb N1.551 (Figures 1D and 3) and 4% reacted positively to MAb F1.652 (Figures 1E and 3). These positively reacting fibers were assumed to be neonatal IIa- and embryonic MHC-containing fibers, respectively. These two developmental MHC isoform-containing fibers were difficult to identify in the DGMp.
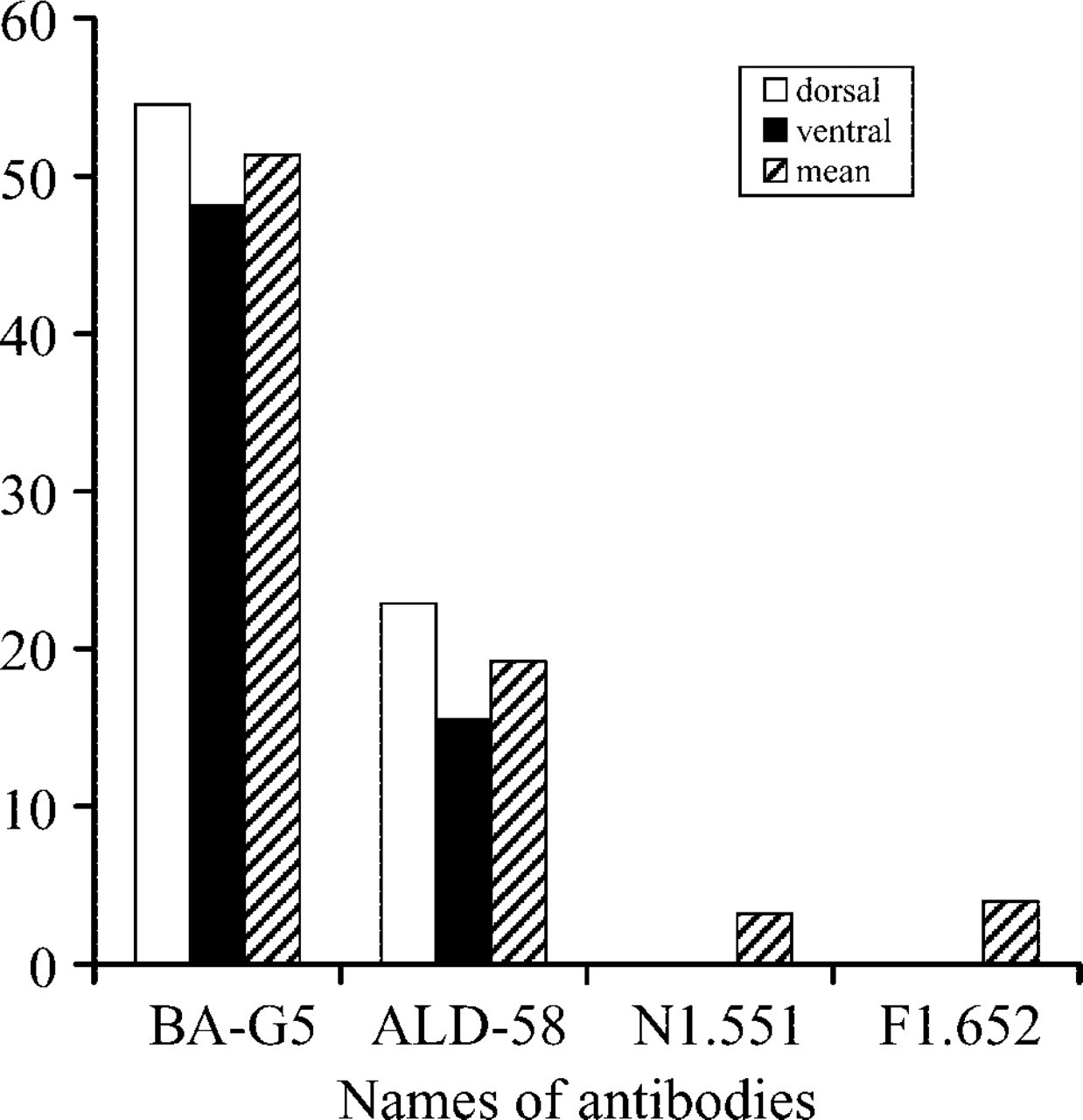
Immunofluorescence showing the percentage of muscle fibers containing unique and developmental MHC isoforms in adult human anterior DGM. More MHC-α– and MHC-ton-containing fibers are concentrated in the dorsal portion compared with the ventral portion.
In addition, two DGMs removed from the right side were processed using the aforementioned methods. The results did not show significant differences in fiber type and distribution between the DGMs from the left and right sides.
To confirm the specificity of the MAbs used in this study, the sections from the human masseter and biceps brachii served as positive and negative controls, respectively. The former is a well-characterized cranial muscle that contains specific and developmental MHC isoforms in its fibers, whereas the latter does not contain these MHC isoforms (Bredman et al. 1991; Sciote et al. 1994; Mu et al. unpublished data) The immunofluorescent microscopy showed that the masseter muscle contained many α-mfs (Figure 1B′) and ton-mfs (Figure 1C). Some masseter muscle fibers also reacted with MAb N1.551 (Figure 1D′) and F1.652 (Figure 1E′). Conversely, the muscle fibers of the adult human biceps brachii tested were negative for the unique and developmental MHC isoforms (results not shown).
Western Blotting Analysis
Immunoblotting of the proteins from adult DGMa showed distinct bands that reacted to MAbs BA-G5 (Figure 4A), ALD-58 (Figure 4B), N1.551 (Figure 4C), and F1.652 (Figure 4D). Even though the same amount of protein was loaded in the wells, the densities of the bands were not the same. In general, the bands that reacted with MAb N1.551 or F1.652 were lighter than those that reacted with MAbs BA-G5 and ALD-58. The electrophoretic immunoblotting for three DGMp samples also showed distinct bands that reacted with the aforementioned four MAbs. Although almost no positive muscle fibers could be observed in the sections of DGMp reacted to MAb N1.551 or F1.652, the existence of neonatal IIa and embryonic MHC isoforms was distinctly identified by the immunoblotting technique (results not shown). The immunoblots using proteins from human masseter showed a positively labeled single band on each lane of the membranes, which reacted respectively to the four MAbs mentioned above (Figures 4A′–4D′). The human biceps brachii sample did not exhibit any positive band on membrane reacted with the same MAbs (Figures 4A“–4D”).
Discussion
The human DGM actively participates in chewing, swallowing, breathing, and speech as demonstrated by electromyography (EMG) studies (Van de Graaff et al. 1984; Strohl et al. 1987; Spiro et al. 1994; Thexton and McGarrick 1994). During swallowing, both bellies display EMG activities with high amplitude and short duration (Widmalm et al. 1988). The most marked action of the DGMa has been observed during jaw opening. However, it is also active during protrusion, lateral jaw movements to either side, and placement of the tongue tip on the hard and soft palate and on the floor of the mouth (Berzin 1995; Castro et al. 1998, 1999). This belly also responds to sudden unloading of the mandibular elevator muscle in younger and older adults (Karkazis et al. 1993).
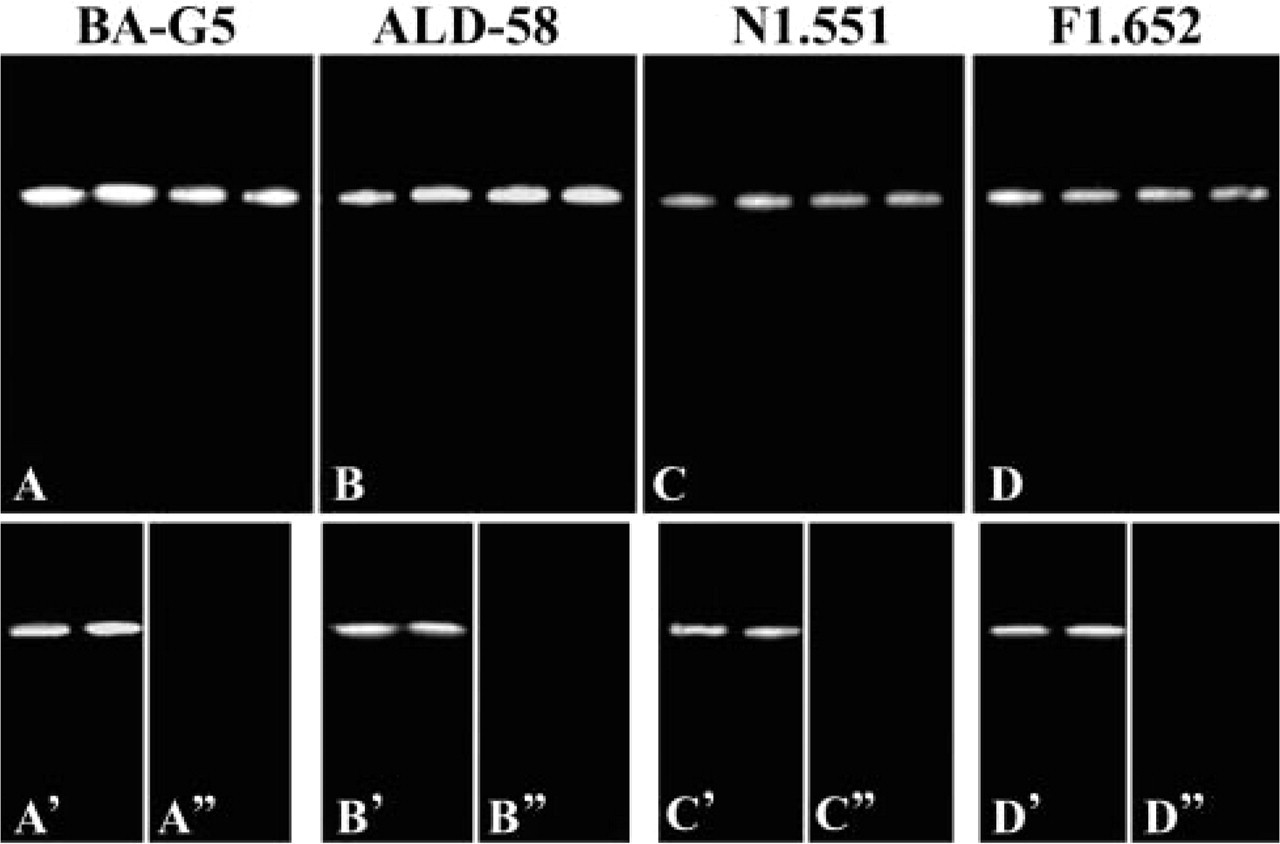
Immunoblotting analysis of unique and developmental MHC isoforms in human anterior DGM and two control muscles. Electrophoretic immunoblotting was performed for the samples of human anterior DGM (
The functional characteristics of a muscle are mainly related to the expression of different MHC isoforms in its muscle fibers. In the skeletal muscles of human trunk and limbs, three major MHC isoforms (i.e., MHCI/β, IIa, and IIx) have been reported, and a single fiber may contain either only one type of MHC isoform or multiple isoforms. The expression and distribution patterns of the MHC isoforms in a muscle highly correlate with the functional demands of the muscle. Electrophysiological studies have demonstrated that the maximal velocity of shortening of muscle fibers is largely determined by and highly dependent on their MHC isoform composition (Reiser et al. 1985; Larsson and Moss 1993). In addition to the major MHC isoforms (i.e., MHCI, IIa, IIx, and IIb), the unique (i.e., MHC-α and MHC-ton) and developmental (i.e., neonatal and embryonic) MHC isoforms could be found in restricted cranial muscles, and the tissue-specific superfast MHC isoform was found in extraocular and masticatory muscles (Rowlerson et al. 1982; Sartore et al. 1987). In normal adult mammals, these MHC isoforms are not expressed in the extrafusal fibers of the skeletal muscles in the trunk and limbs (derived from the somites of the embryos) but are found in the muscle fibers of some specialized craniocervical skeletal muscles (derived from the rostral somitomeres). The differences may be attributable to their different embryonic origin, but the different concentrations and distribution patterns of these various isoforms probably reflect the physiological demands of the muscle for performance of various tasks. For example, Bredman and co-workers (1992) studied the α-MHC expression in rabbit masseter during postnatal development and found that the number of α-mfs increased gradually from a few to large numbers from the neonatal period to 28 days. These changes are closely correlated with changes in the diet of the young animals. The results suggested that the transformation of MHC expression occurs progressively throughout all the postnatal lifetime. As one of the craniocervical skeletal muscles, the expression of unique and developmental MHC isoforms in the DGM is based on its embryonic origin, which provides different potentials for expression of these isoforms. However, the numbers and localizations of the fibers containing these isoforms correlate with the work demands of this muscle. That is, the MHC isoform profile of muscle fibers is not definitively established but undergoes changes according to workload and activity.
Although the functional significance of the α-mfs and ton-mfs in skeletal muscles remains unclear, these fibers have contractile properties that differ from those seen in either type I or type II muscle fibers. For example, physiological studies at the single-fiber level have demonstrated that α-mfs are faster than type I but slower than type IIA fibers. They have an intermediate level of maximal velocity of shortening (Sciote and Kentish 1996). From fast to slow, the MHC isoforms are expressed in the following order: MHCIIb-MHCIIx/d-MHCIIa-MHCα-MHCI/β. The presence of MHC-α in a skeletal muscle is most likely to fill the functional gap between type I and type II fibers. A considerable number of α-mfs identified in the human DGM may represent an evolutionary adaptation for enhancing more precise control of this muscle. The MHC-ton fibers are usually characterized by multiple innervation and en-grappe motor endplates (Hess 1970; Han et al. 1999a). Physiologically, the contraction of the ton-mfs is slow and prolonged, and can be adjusted in small gradations with only modest expenditure of energy (Kuffler and Williams 1953). Their contraction is postural or prolonged isometric. The ton-fibers in the DGM appear to be essential for various postures of the mandible seen during chewing, swallowing, breathing, and speaking. The expression of MHC-α and MHC-ton provides the molecular bases that enable these versatile activities to be performed precisely in the human DGM.
Footnotes
Acknowledgements
Supported by an NIH Grant 1R01 DC-04728 from the National Institute on Deafness and Other Communication Disorders (to Dr. L. Mu).
We thank the Department of Pathology of the Mount Sinai School of Medicine for providing specimens for this study.