
Abstract
The family of Ca2+-dependent activator proteins for secretion (CAPS) is involved in dense-core vesicle exocytosis. CAPS1/CADPS1 and CAPS2/CADPS2 have been identified in mammals. CAPS1 regulates catecholamine release from neuroendocrine cells, whereas CAPS2 is involved in the release of brain-derived neurotrophic factor and neurotrophin-3 from cerebellar granule cells. CAPS1 and CAPS2 are predominantly expressed in brain. Here we show the immunohistochemical localization of the CAPS family proteins in various mouse tissues. In the pituitary gland, CAPS1 and CAPS2 were localized to the pars nervosa and the pars intermedia, respectively. In non-neural tissues, CAPS1 was observed in the islets of Langerhans, minor cell types of the spleen and stomach, and medullary cells of the adrenal gland, whereas CAPS2 was present in bronchial epithelial cells, thyroid parafollicular cells, chief cells of the stomach, ductal epithelium of the salivary gland, kidney proximal tubules, and minor cell types of the thymus, spleen, and colon. These results suggest that secretion from distinct cell types in various tissues involves either or both members of the CAPS family.
Ca2+-DEPENDENT activator protein for secretion (CAPS) participates in the priming step of dense-core vesicle (DCV) exocytosis (Grishanin et al. 2004). Two CAPS family members, CAPS1/CADPS1 (Berwin et al. 1998; Tandon et al. 1998; Renden et al. 2001) and CAPS2/CADPS2 (Cisternas et al. 2003; Speidel et al. 2003; Sadakata et al. 2004), have been identified in mammals. A recent study showed that mice lacking the CAPS1 gene die shortly after birth, suggesting that CAPS1 is involved in catecholamine loading of DCVs in embryonic chromaffin cells (Speidel et al. 2005). In Drosophila (Renden et al. 2001) and Caenorhabditis elegans (Avery et al. 1993), only a single CAPS gene is expressed, and Drosophila lacking the CAPS gene is embryonic lethal due to impairment in DCV release (Renden et al. 2001).
DCV release is an important function in various tissues. However, the mechanism of secretion is not well known because the detailed molecular regulation of DCV release has not yet been thoroughly examined (Kallenberg 2000; Cremona et al. 2004). Both CAPS1 and CAPS2 mRNAs are detected in various mouse tissues, including brain, at different expression levels (Cisternas et al. 2003; Speidel et al. 2003; Sadakata et al. 2004). We previously showed that CAPS1 and CAPS2 proteins are differentially distributed throughout the mouse brain (Sadakata et al. 2006). In the postnatal brain, CAPS1 is widespread at different levels. On the other hand, CAPS2 is localized to distinct cell types and fibers of various brain regions including the olfactory bulb, cerebrum, hippocampal formation, thalamus, mesencephalic tegmentum, cerebellum, medulla, and spinal cord, except for some regions that overlapped with CAPS1 (Sadakata et al. 2006). However, the cellular distribution of CAPS1 and CAPS2 proteins in non-neural tissues remains largely unclear. In the present study we analyzed the immunohistochemical distribution patterns of both CAPS1 and CAPS2 proteins in mouse non-neural tissues to understand the biological significance of the CAPS protein family and indicated the comparative distribution patterns of the CAPS family in these tissues.
Materials and Methods
Animal Experiments
Animals were purchased from Nihon SLC (Hamamatsu, Japan). All procedures were conducted according to the RIKEN guidelines for animal research and were approved by the RIKEN Animal Experiments Committee.
Antibodies
Guinea pig polyclonal anti-CAPS2 antibody and rabbit polyclonal anti-CAPS1 antibody (Sadakata et al. 2004) were used for Western blotting and immunohistochemical analysis (diluted 1:10,000 and 1:5000, respectively). Specificity of the CAPS antibodies was examined as previously described (Sadakata et al. 2006). Briefly, Western blotting indicated that the affinity-purified anti-CAPS2 and anti-CAPS1 antibodies specifically recognized single major bands (∼150 kDa) in mouse brain homogenates, which is consistent with previous studies (Speidel et al. 2003). Specificity of the antibodies was verified by immunostaining of either endogenous CAPS1 protein or exogenously expressed CAPS2 protein in PC12 cells: the anti-CAPS1 antibody reacted with endogenous CAPS1 but not exogenous CAPS2, whereas the anti-CAPS2 antibody reacted with exogenous CAPS2 protein but not with endogenous CAPS1. Rabbit anti-oxytocin antibody (cat. #AB911; Chemicon International, Temecula, CA), which was prepared against synthetic oxytocin (Sigma; St Louis, MO) conjugated to thyroglobulin as an immunogen, was used at a dilution of 1:5,000 and produced a specific staining pattern in the pars nervosa of the rat pituitary as described previously (Samson and Schell 1995). For immunohistochemical analyses, Alexa 555-conjugated anti-guinea pig and Alexa 647-conjugated anti-rabbit antibodies (Molecular Probes; Eugene, OR) were used as secondary antibodies at a dilution of 1000.
Preparation of Subcellular Fractions
P21 C57BL/6J male mice (8.5-9.5 g) were used after being sacrificed by anesthesia with diethyl ether. Postnuclear supernatants from various tissues were prepared as previously described (Yan et al. 2000). Briefly, mouse tissues were dissected and homogenized in homogenization buffer (10 mM HEPES, pH 7.4, 38 mM NaCl, 5 mM EDTA, 1 mM 2-ME, protease inhibitor cocktail) using a glass/Teflon homogenizer. Homogenates were centrifuged at 900 × g for 10 min, and equal amounts of protein samples per lane (10 μg/lane) were subjected to Western blotting.
Immunohistochemistry
P21 C57BL/6J male mice (8.5-9.5 g) were used after being sacrificed by anesthesia with diethyl ether. Postnatal 15-week (P15w) nurturing female mice (21.0-23.0 g) were used only for immunohistochemistry of the pituitary. Mice were transcardially perfused, initially with PBS and then with 4% PFA in PBS. Tissues were dissected out, postfixed in 4% PFA at 4C for 5 hr, and cryoprotected by immersion in 15% sucrose in PBS overnight at 4C. After embedding in Tissue-Tek OCT compound (Sakura Finetechnical; Tokyo, Japan), tissues were frozen in dry ice powder and sectioned at a thickness of 14 μm using a cryostat (CM1850; Leica Microsystems, Frankfurt, Germany) at −18C. Sections were air dried for 1 hr and rinsed three times in PBS. After blocking with 5% normal donkey serum (Vector Laboratories; Burlingame, CA) in PBS, sections were reacted with specific primary antibodies at 4C overnight, rinsed in PBS, reacted with the appropriate secondary antibody at room temperature for 1 hr, and again rinsed in PBS. Immunoreacted sections were mounted with Vectashield (Vector Laboratories) mounting medium and observed using a confocal laser microscope (LSM 510 META; Carl Zeiss, Jena, Germany). Digital images were processed using Adobe Photoshop 6.0 software (Adobe Systems; Palo Alto, CA). To determine the cellular localization of CAPS proteins in each tissue, two independent immunohistochemical experiments were performed on each tissue sample from each of three different animals. All results shown in this study were reproducible. All sections used were processed together in the same batch of reagents.
Semi-quantitative RT-PCR
RNA was extracted from the fresh pancreas of P21 C57BL/6J male mice (8.5-9.5 g) with TRIzol Reagent (Invitrogen; Carlsbad, CA) according to the manufacturer's instructions. RT-PCR reactions were carried out in a 20-μl volume in the GeneAmp PCR System 9700 (Perkin Elmer; Wellesley, MA). The Qiagen OneStep RT-PCR kit (Qiagen; Hilden, Germany) was used according to the manufacturer's instructions. A 50-ng sample of total RNA template was used per reaction. Reverse transcription cycling conditions were 30 min at 50C followed by 15 min at 95C. PCR cycling conditions consisted of 27 or 30 cycles of 94C for 30 sec, 60C for 30 sec, and 72C for 30 sec, followed by a final hold at 72C for 5 min. Primers were as follows. Mouse CAPS1 exon 2-3 forward primer: 5′-tcagcaaacagcagctacag-3′; exon 2-3 reverse primer: 5′-agatggcatcaaacttggcc-3′ (exon 2-3 PCR product: 291 bp); exon 5-6 forward primer: 5′-attgaggagctgaagtcctc-3′; exon 5-6 reverse primer: 5′-tggacttccatgatcaccac-3′ (exon 5-6 PCR product: 206 bp); exon 10-11 forward primer: 5′-acacttttgccatgtgcagc-3′; exon 10-11 reverse primer: 5′-catccagctgaggcacattt-3′ (exon 10-11 PCR product: 294 bp); exon 11-12 forward primer: 5′-cgagccttcttcaatgcagt-3′; exon 11-12 reverse primer: 5′-tctgtgatccaaagtcagcc-3′ (exon 11-12 PCR product: 312 bp); exon 14-15 forward primer: 5′-tggccaggtgtttgtgttag-3′; exon 14-15 reverse primer: 5′-cagaactcggagtctctctt-3′ (exon 14-15 PCR product: 250 bp); exon 19-20 forward primer: 5′-tgtaggccgcttaatcactc-3′; exon 19-20 reverse primer: 5′-aagagtgggaaactgtccca-3′ (exon 19-20 PCR product: 228 bp); exon 21-22 forward primer: 5′-aaacacctgcaggacctgtt-3′; exon 21-22 reverse primer: 5′-tatagcagccatccatgacg-3′ (exon 21-22 PCR product: 241 bp); exon 23-24 forward primer: 5′-ggaaacttgatgcccttcag-3′; exon 23-24 reverse primer: 5′-tcttggcccatttccatgct-3′ (exon 23-24 PCR product: 277 bp); exon 29-30 forward primer: 5′-ggtacaacagctccatgaac-3′; and exon 29-30 reverse primer: 5′-tgctgataccttgtaagccc-3′ (exon 29-30 PCR product: 243 bp). PCR was carried out at least three times for each number of cycles. PCR products were electrophoresed on 2% agarose gels with a 100-bp DNA Ladder (cat. #15628-050; Invitrogen).
Results
Expression Patterns of CAPS Family Proteins
We first examined the expression patterns of CAPS1 and CAPS2 in 11 tissues of P21 male mice by Western blotting. CAPS2 immunoreactivity was detected as a 150-kDa band at high levels in brain and pituitary, at low to moderate levels in lung and pancreas, and at very low levels in stomach and kidney (Figure 1A). An additional 140-kDa band immunoreactive to anti-CAPS2 antibody was detected in stomach and was likely to be an alternatively spliced variant (Speidel et al. 2003). On the other hand, CAPS1 immunoreactivity was localized as a 150-kDa band at high levels in brain and pituitary and at a low level in pancreas (Figure 1B). An additional 70-kDa band immunoreactive to anti-CAPS1 antibody was detected in pancreas. To evaluate the integrity of protein samples without proteolytic degradation, we incubated pancreatic samples at 37C for 15 min and analyzed their Western blotting patterns. As a result of the incubation, the 150-kDa CAPS1-immunoreactive band disappeared with two bands of 40 and 70 kDa being detected instead (Figure 1B, rightmost lane). Moreover, to examine the expression of CAPS1 mRNA or its splicing variation in pancreas, we performed quantitative RT-PCR using nine sets of exon-specific PCR primers (for exon 2-3, exon 5-6, exon 10-11, exon 11-12, exon 14-15, exon 19-20, exon 21-22, exon 23-24, and exon 29-30) and total pancreas RNAs as a template. The reactions with eight primer sets produced PCR products of the expected molecular sizes after 30 cycles but not after 27 cycles; however, the exon 21-22-specific primer set we designed did not produce the expected band even after 30 cycles (data not shown).
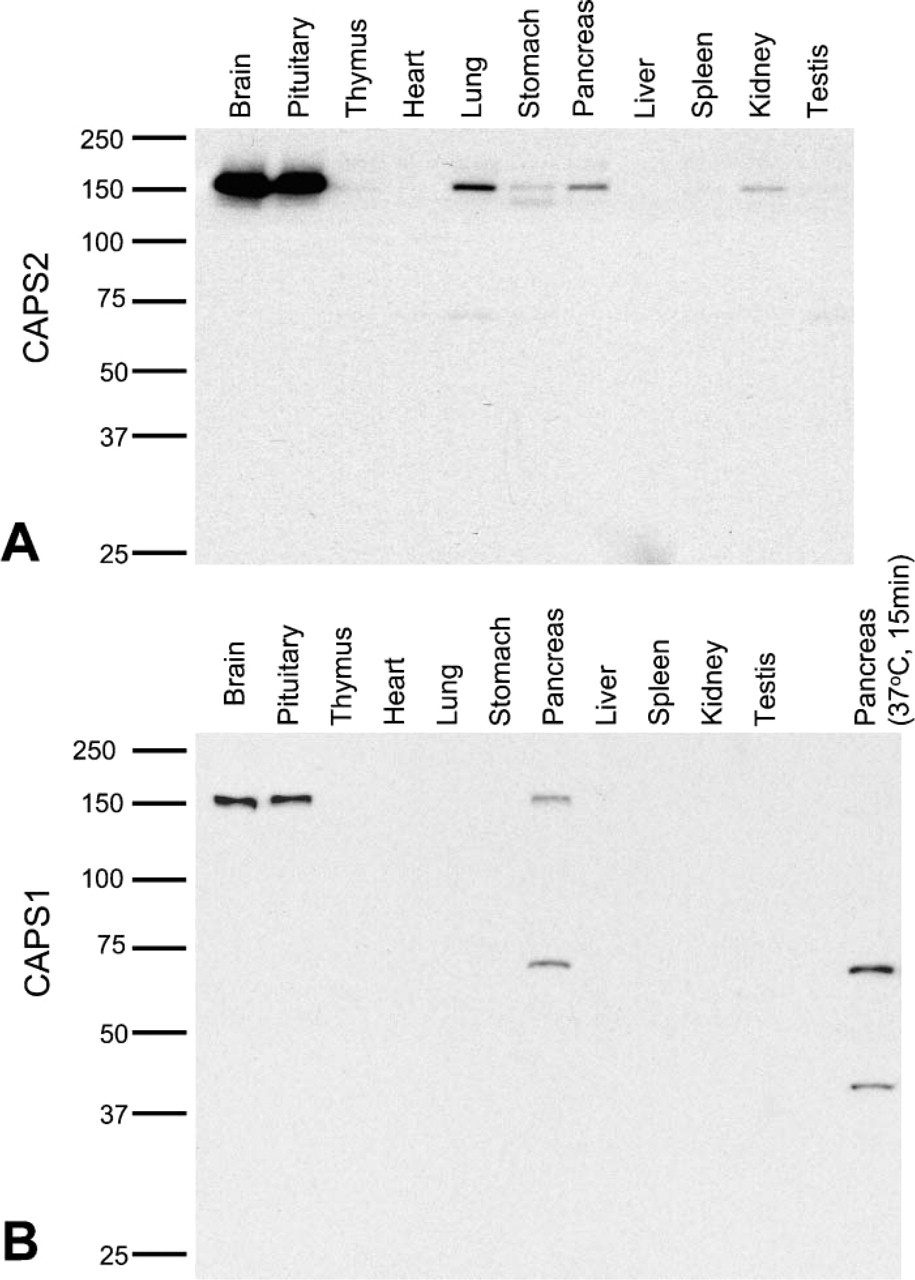
CAPS protein expression in different tissues. Homogenates from the indicated mouse tissues [postnatal week 21 (P21w)] were analyzed by Western blotting using specific antibodies to CAPS2
Distribution of CAPS Family Proteins in the Endocrine System
To examine the cellular distribution of CAPS family proteins in the endocrine system, we immunohistochemically analyzed mouse pituitary gland, thyroid, and adrenal gland. In the pituitary, the master gland controlled secretion of hormones and consisted of three lobes (pars anterior, pars intermedia, and pars nervosa), CAPS1 immunoreactivity was predominantly localized in the pars nervosa but was present in low to moderate levels in the pars intermedia (Figure 2B). In contrast, strong immunoreactivity for CAPS2 was observed in the pars intermedia with only very low immunoreactivity seen in the pars nervosa (Figure 2A). In addition, CAPS2 immunoreactivity was partially overlapping with that of oxytocin and was also located in the surroundings in the pars nervosa of nurturing female mice (Figures 2D-2F). Either CAPS1 or CAPS2 immunoreactivity was barely detected in the pars anterior (Figures 2A-2C).
In the thyroid, intense CAPS2 immunoreactivity was observed in parafollicular cells, a subset of cells lining sphere-shaped structures called follicles (Figures 2G and 2J). On the other hand, there was little immunoreactivity for CAPS1 in either thyroid follicular or parafollicular cells (Figures 2H and 2K). Neither CAPS1 nor CAPS2 immunoreactivity was observed in the parathyroid (data not shown).
In the adrenal gland, strong CAPS1 immunoreactivity was observed in the medulla, which is responsible for producing epinephrine and norepinephrine, but very little was seen in the cortex (Figure 2N) as previously shown by biochemical analyses (Walent et al. 1992). On the other hand, CAPS2 immunoreactivity was low in these medullary cells (Figure 2M). We noted that in the adrenal medulla there was a patchy distribution of at least two cell clusters, each expressing one CAPS family member more preferentially than the other, i.e., CAPS1-dominant clusters and CAPS2-dominant clusters (Figures 2P-2R).
Distribution of CAPS Family Proteins in the Lymphatic System
In lymph-forming organs such as thymus and spleen, the immunostaining levels for both CAPS1 and CAPS2 were very low (Figure 1). In the medullary zone of the thymus, moderate levels of CAPS2 were localized in a small subset of cells (Figure 3A). In the red pulp of the spleen, moderate levels of CAPS1 and CAPS2 were specifically colocalized in very minor cell types (Figures 3D-3F).
Distribution of CAPS Family Proteins in the Digestive System
In the salivary gland, immunoreactivity of CAPS2 was observed in the ductal epithelia of the submandibular gland (Figures 4A and 4D). Similarly, only CAPS2 immunoreactivity was observed in the ductal epithelia of the other salivary glands, the sublingual gland, and the parotid gland (data not shown).
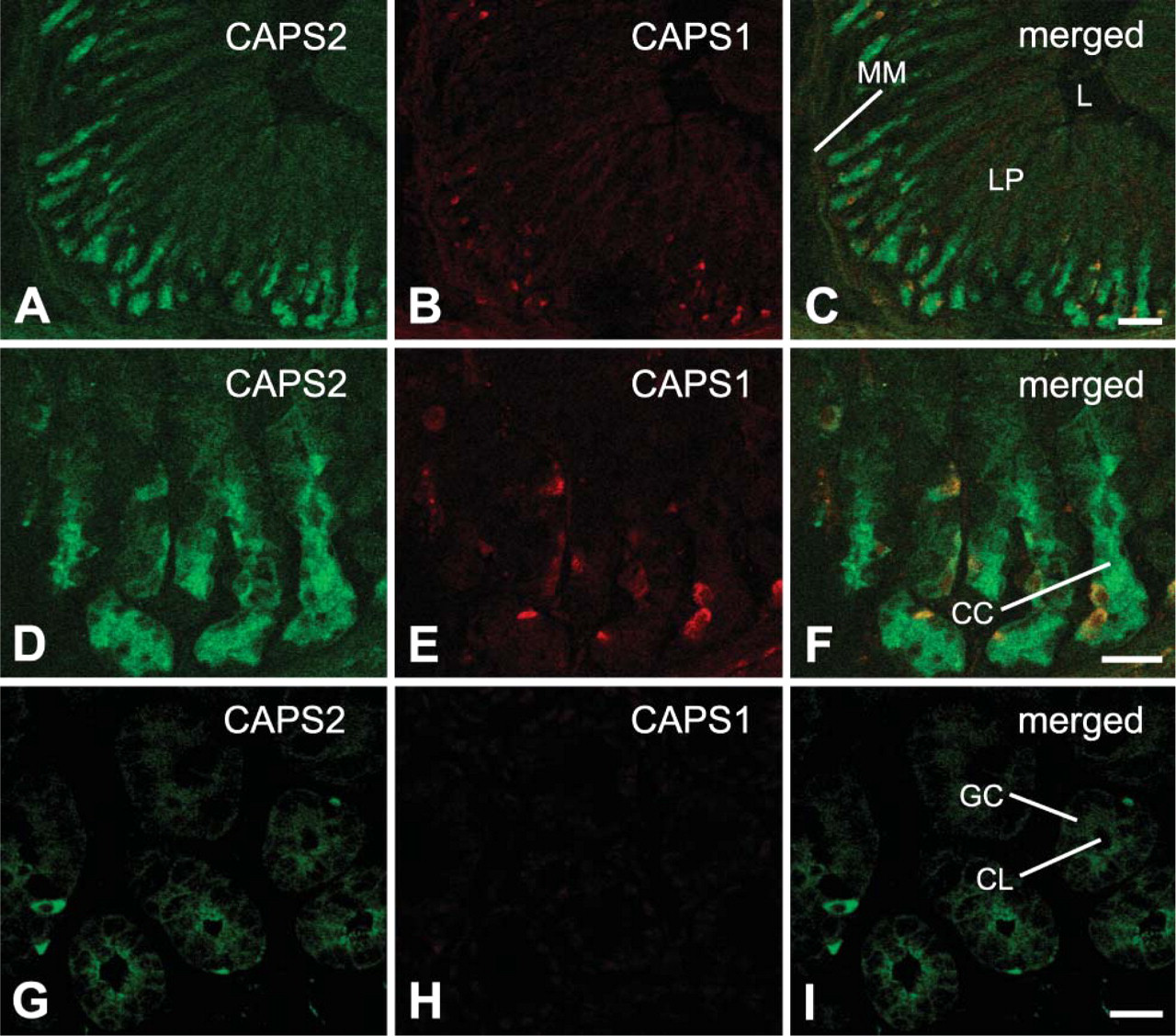
In the pancreas, high levels of CAPS1 immunoreactivity were detected in the islets of Langerhans, the endocrine portion of the pancreas (Figures 4H and 4K). In contrast, CAPS2 immunoreactivity was low in the islets (Figures 4G and 4J).
In the stomach, Western blotting showed expression of CAPS2 protein at a low level but that of CAPS1 at an undetectable level (Figure 1). CAPS2 immunoreactivity was concentrated in gastric chief cells deeper in the gastric glands (Figures 5A and 5D). In addition, both CAPS1 and CAPS2 were shown to be colocalized in minor cell types (Figures 5D-5F).
In the intestines, CAPS2 immunoreactivity was localized in minor cell types of the ascending colon (Figure 5G), whereas immunoreactivity for either CAPS2 or CAPS1 was barely detected in jejunum and ileum (data not shown).
Distribution of CAPS Family Proteins in the Respiratory and Urinary System
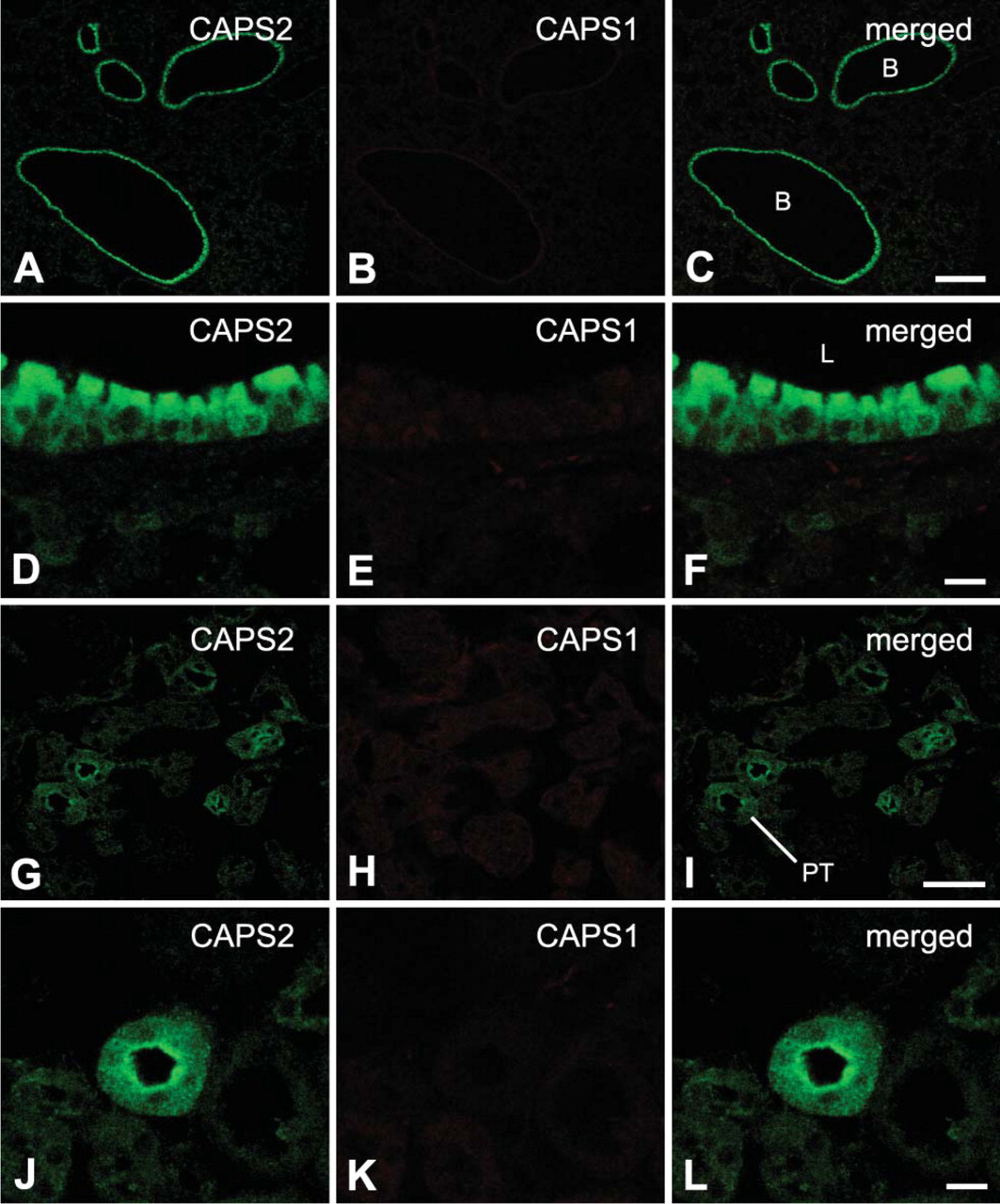
In the lung, CAPS2 immunoreactivity was observed in the bronchial epithelium (Figures 6A and 6D), whereas CAPS1 was detected at a low level within the bronchial smooth muscle layer (Figures 6E).
In the kidney, low to moderate immunostaining levels for CAPS2 were detected in renal proximal convoluted tubules (Figures 6G and 6J), which secrete diverse organic anions (Anzai et al. 2005). However, only weak immunoreactivity for CAPS1 was detected in this tissue (Figures 6H and 6K).
Discussion
We demonstrated by immunohistochemistry that the two CAPS family member (CAPS1 and CAPS2) proteins are differentially localized to distinct cell types of various tissue regions including the pituitary, salivary gland, thyroid gland, lung, thymus, stomach, pancreas, spleen, kidney, adrenal gland, and intestine. CAPS family proteins are membrane associated; however, a substantial portion of these proteins can be found in the soluble fraction (Berwin et al. 1998; Speidel et al. 2003). Both CAPS1 and CAPS2 immunoreactivities in non-neural tissues observed showed largely cytosolic localization patterns.
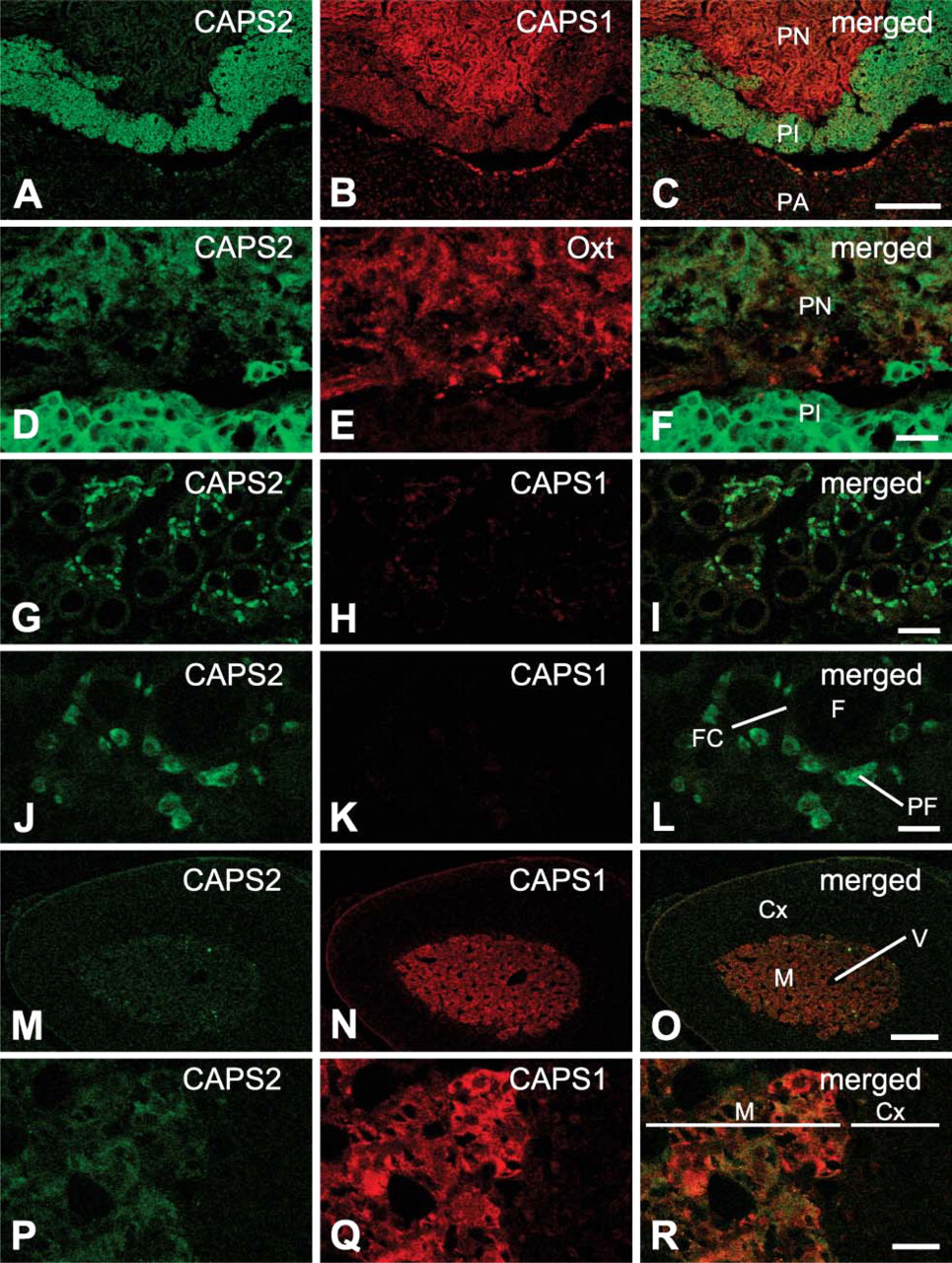
Immunohistochemical distribution of the CAPS family proteins in the mouse endocrine system.
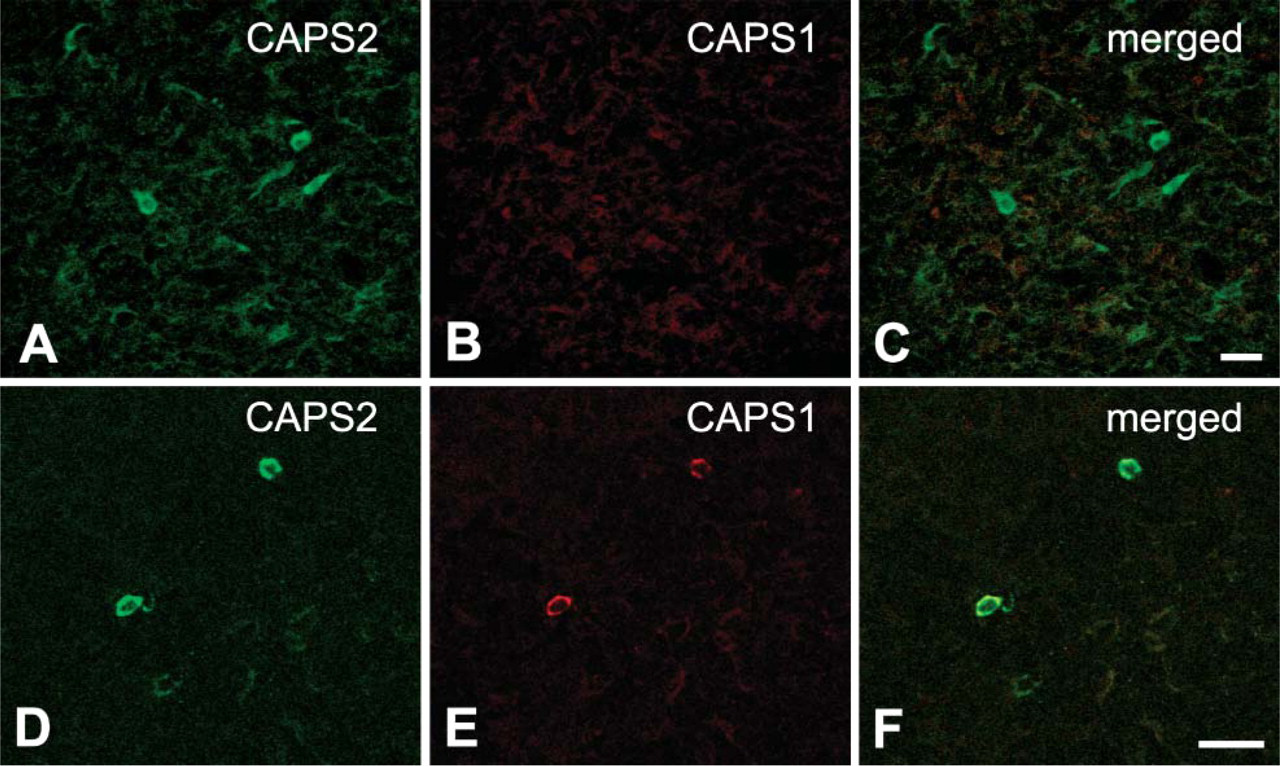
Immunohistochemical distribution of the CAPS family proteins in the mouse lymphatic system.
The cloned mouse CAPS2 cDNA encodes a 150-kDa protein, which can be detected in mouse brain, pituitary, lung, stomach, pancreas, and kidney by Western blotting as shown in Figure 1. On the other hand, the stomach expresses an additional 140-kDa immunoreactive band, which seems to correspond to one of several splicing variants reported previously (Speidel et al. 2003). The biological significance of these splicing variants has not been reported (Speidel et al. 2003; Sadakata et al. 2004). The present data indicate that the 150-kDa CAPS2 has a role in secretion in the stomach, although it remains to be studied whether the 140-kDa variant has a function different from that of the 150-kDa CAPS2 and/or a different expression pattern in stomach cells.
The pituitary gland consists of three lobes (pars anterior, pars intermedia, and pars nervosa). Axon terminals from the supraoptic and paraventricular nuclei of the hypothalamus are located in the pars nervosa. Oxytocin is produced by these hypothalamic neurons in the paraventricular nucleus and supraoptic nucleus and is transported to the pituitary for release (Kikusui et al. 1997). On the other hand, the pars intermedia secretes melanocyte-stimulating hormone as its principal product. This part of the pituitary also produces adrenocorticotropic hormone, corticotropin-like intermediate lobe peptide, and beta-endorphin (Saland 2001). Our data show that CAPS1 protein is present at high levels in the pars nervosa and at low to moderate levels in the pars intermedia. In contrast, CAPS2 protein is observed more preferentially in the pars intermedia than in the pars nervosa. CAPS2 immunoreactivity is partially overlapping with that of oxytocin and is also located in the surroundings in the pars nervosa of nurturing female mice (Figures 2D-2F). These findings indicate that the two CAPS family member proteins are involved in lobe-specific secretions in the pituitary.
The thyroid gland uptakes iodine from foods to produce two thyroid hormones, thyroxine and triiodothyronine (Morreale de Escobar et al. 2004). In the thyroid, intense CAPS2 immunoreactivity was observed in parafollicular cells (Figures 2G and 2J), whereas there was little immunoreactivity for CAPS1 in either thyroid follicular or parafollicular cells (Figures 2H and 2K). Neither CAPS1 nor CAPS2 immunoreactivity was observed in parathyroid (data not shown). These results suggest a strong involvement in the secretion of calcitonin from these cells (Inzerillo et al. 2002).
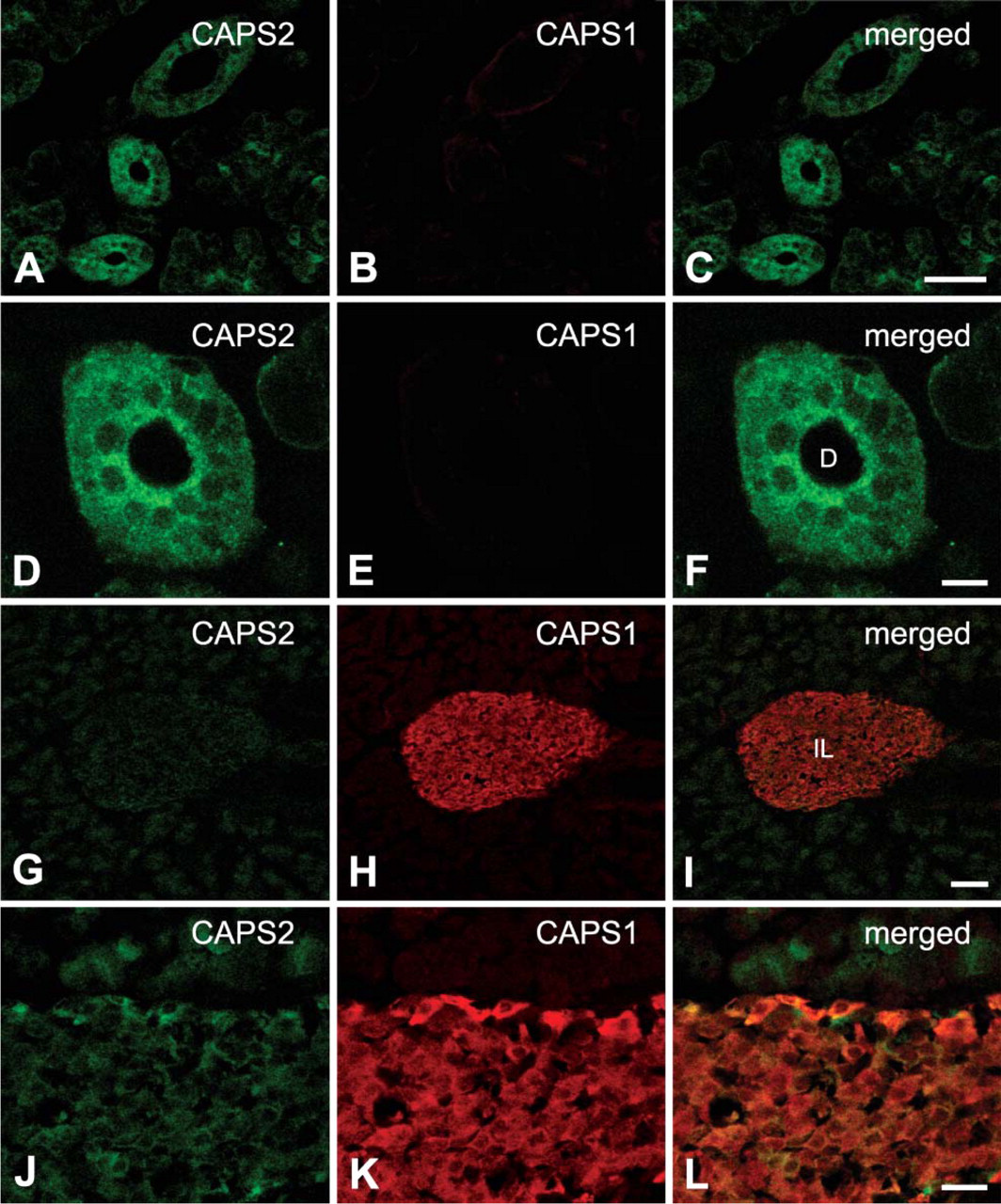
Immunohistochemical distribution of the CAPS family proteins in the mouse digestive system.
Immunohistochemical distribution of the CAPS family proteins in the mouse digestive system.
In lung, both CAPS1 and CAPS2 proteins were colocalized in the bronchial epithelium (Figure 6F). Notably, a high level of CAPS2 was localized in ciliated epithelial cells that have an important role in airway function and also modulate airway inflammatory events by the secretion of a variety of proinflammatory mediators including cytokines (Levine et al. 1993). In addition to the bronchial epithelium, punctate immunoreactivity of CAPS1 was detected at a low level within the bronchial smooth muscle layer where nerve fibers are located (Figure 6E). These data suggest a role for the CAPS family in bronchial function.
Immunoreactivity of CAPS2 was concentrated in gastric chief cells deep in the gastric glands. Chief cells are known to secrete pepsinogen (an inactive form of the pepsin-digesting enzyme pepsin); thus, CAPS2 appears to have a role in pepsinogen secretion. Moreover, both CAPS1 and CAPS2 immunoreactivities were colocalized in minor cell types of the stomach. In this regard, it is of note that Wassenberg and Martin (2002) reported that CAPS is expressed in gastrin-containing G cells of the stomach. These data suggest that the two CAPS family members are involved in the mechanism underlying the secretion of substances such as gastrin.
Immunohistochemical distribution of the CAPS family proteins in the mouse respiratory and urinary systems.
In the lymph-forming thymus and spleen tissues, we detected positive immunoreactivity of CAPS family proteins only in a small subset of cells. Previous studies reported the expressions of neurotrophin-3 (Katoh-Semba et al. 1996), brain-derived neutrotrophic factor (Katoh-Semba et al. 1997), vasoactive intestinal peptide (Gomariz et al. 1993), and somatostatin (Aguila et al. 1991) in minor cell types of these two tissues. Expression of methionine-enkephalin, neuropeptide Y, and substance P were also reported in a similar cell subset of the thymus (Atoji et al. 1997). Together our data suggest a possible role for the CAPS family in the release of these secretory substances from distinct minor cell populations in the thymus and spleen.
It was previously shown that CAPS1 is involved in the exocytosis of insulin in the pancreas (Waselle et al. 2005). Our immunohistochemical analyses show strong immunoreactivity for CAPS1 in the islets of Langerhans (Figure 4H). Moreover, a low level of CAPS2 immunoreactivity was also observed in the islets of Langerhans (Figure 4G). An additional 70-kDa band immunoreactive to anti-CAPS1 antibody was detected in the pancreas with Western blotting (Figure 1B). Incubation of pancreatic samples at 37C for 15 min to evaluate the integrity of protein samples resulted in the disappearance of the 150-kDa CAPS1-immunoreactive band and the appearance of only two CAPS1-immunoreactive bands of 40 and 70 kDa (Figure 1B). Moreover, quantitative RT-PCR using nine sets of exon-specific PCR primers could not show a differential expression level for various CAPS1 exons (data not shown). Together these data indicate that the pancreatic 70-kDa band is a product of partial degradation of CAPS1 protein rather than a splicing variant. It remains elusive whether CAPS2, which is expressed at a low level, is functional or not in this tissue.
In the present study we could not conclusively determine the immunolocalization of CAPS1 and CAPS2 in testes. Although our previous study indicated expression of CAPS2 mRNA in the testes of adult mice by RT-PCR (Sadakata et al. 2004), the present study showed no detectable levels of CAPS2 protein in P21 tissues by either Western blotting or immunohistochemistry. This discrepancy may be due to instability of untranslated mRNAs in male germ cells (Kleene 1996).
Until now, the CAPS family proteins had been characterized as endocrine-related proteins. However, present data show that CAPS2 protein is localized in some exocrine-related cells such as gastric chief cells, renal cells of proximal convoluted tubules, and ductal epithelial cells of salivary gland. Thus, the CAPS family proteins are likely to have a common role in both secretory exocrine and endocrine events.
Footnotes
Acknowledgements
This study was supported by grants-in-aid for Scientific Research from the Japanese Ministry of Education, Culture, Sports, Science, and Technology (grant number #17700322), the Japan Society for the Promotion of Science, and the Institute of Physical and Chemical Research (RIKEN).