
Abstract
The final elimination step of self-reactive T cells occurs in the medulla of the thymus where a complex framework provided by stromal cells supports an optimal milieu for their selection. Here we present evidence that tight junctions (TJs) widely join medullary stromal cells of the human thymus. Occludin (OCLN) and claudin-1 (CLDN-1) of TJ-associated molecules were dominantly expressed in medullary thymic epithelial cells (mTECs), and CLDN-4 and CLDN-7 were also localized in some mTECs near Hassall's corpuscles. Interestingly, p53-like transcription factors were found to upregulate OCLN and CLDN-1 in human TEC lines, as recently suggested in the regulation of mTEC function. Furthermore, dendritic cells (DCs) of the medulla, with a major role for selection of thymocytes, expressed CLDN-1 and OCLN as well, implying that the interposition of DCs within the mTEC scaffold is also helped by TJs. Analysis of freeze-fracture replicas of the thymus revealed TJ strand structures in the vicinity of gap junction plaques through which small molecules might move, as implied by dye-transfer analysis of a medullary cell line. Thus, it is thought that p53-like molecules regulate TJ-associated interactions of medullary stromal cells and that this mechanism might be associated with an intercellular communication network, probably for preserving the medullary niches.
T
Tight junctions (TJs) that form strands have basically been recognized as hallmark structures of epithelia with central biological importance as “fences” and “gates.” TJs are composed of the integral membrane proteins occludin (OCLN), first found as a component of TJ strands, and claudins (CLDNs), a family of over 20 genes in humans (Tsukita and Furuse 2000). There is accumulating evidence that, in addition to epithelial cells, circulating blood cells such as neutrophils bind through the TJ apparatus to stationary epithelium to efficiently induce inflammatory responses (Huber et al. 2000; Kucharzik et al. 2001; Zen et al. 2004). More recently, it was reported that DCs also expressed TJs within intestinal and nasal mucosal epithelia, suggesting that TJs might have a role not only in arranging interepithelial connections but also for engaging DCs to promote their immunological activities (Rescigno et al. 2001; Ichiyasu et al. 2004; Takano et al. 2005). DCs are highly efficient antigen-presenting cells that induce and regulate immune responses in various tissues and, in the thymus, eliminate self-reactive T cells in the medulla (Sprent and Webb 1995; Brocker 1999; Banchereau and Steinman 1998; Groux et al. 2004). Thus, it is also of great interest whether TJs take part in interstromal cell connections among TECs and DCs, particularly in medullary regions where TECs and DCs are densely localized.
In the current study we show that OCLN and CLDNs, including CLDN-1, CLDN-4, and CLDN-7, of TJ-associated molecules are preferentially expressed in the mTEC network of the human thymus. OCLN and CLDN-1 are broadly expressed there, whereas CLDN-4 and CLDN-7 are localized in the regions near Hassall's corpuscles. Like other epithelial tissues, DCs in the medulla also express CLDN-1 as well as OCLN, as assessed by studies on thymic tissues and primary culture cells. We have previously reported particular distribution profiles of p63 and p73 of p53-related transcription factors in the nuclei of human TEC subtypes (Ichimiya et al. 2002; Yang et al. 2002; Melino et al. 2003; Kikuchi et al. 2004), and in vitro studies using human TEC lines suggest that p63 and p73 have the capacity to upregulate the expression of TJ strand-associated molecules. Freeze-fracture replicas of thymic tissues demonstrate a junctional complex including TJ strand structures and gap junction (GJ) plaques on the cell surface of certain cell types. Interestingly, micro-injection analysis of mTECs revealed that small molecules could be intercellularly transferred, probably through connexin 43 forming GJs. Together these findings imply that mTECs might establish a communication network mutually transferring small molecules, whose architecture is supported by TJs under the control of p53 family members.
Materials and Methods
Thymic Tissues
All tissue was obtained with informed consent and approval by institutional review boards. Normal human thymuses were obtained from patients <3 years of age undergoing cardiovascular surgery for congenital cardiac disease at the Hokkaido Children's Hospital and Medical Center in Japan. Before use for cryostat tissue sections, small tissue specimens were mounted in optimal cutting temperature compound and frozen in liquid nitrogen to be stored at −80C. Six-μm tissue sections were cut and fixed with ice-cold acetone for 5 min. After fixation, slides were air dried and stored at −80C before use.
Cell Lines and Transfection
F2.5 cell lines derived from human TEC (Fernandez et al. 1994) were maintained in MEM supplemented with 10 mmol/liter HEPES, 100 U/ml penicillin G, 50 μg/ml streptomycin, and 10% heat-inactivated fetal bovine serum in a humidified atmosphere at 37C in 5% CO2. Cell line HTS91, originated from human thymic medullary epithelium (Crisa et al. 1997), was maintained in calcium-free WAJAC404 medium, pH 7.5 (Kyokuto Pharmaceutical Industrial; Tokyo, Japan), supplemented with 28 mmol/liter HEPES, 14 mmol/liter sodium bicarbonate, 10 ng/ml epidermal growth factor, 10 nM dexa-methasone, 50 μg/ml streptomycin, and 2% heat-inactivated horse serum in a humidified atmosphere at 37C in 5% CO2. To establish transformed cell lines, p63α and p73α recombined in pIRES-hyg2 and pIRES-puro, respectively, were introduced using Lipofectamine 2000 reagent (Invitrogen; Carlsbad, CA). Transformed cells were subsequently selected in medium supplemented with hygromycin or puromycin. pcDNA3.1 vectors carrying p63α or p73α cDNAs were used to produce expression recombinants of pIRES vectors.
Antibodies
A mouse anti-OCLN monoclonal antibody (MAb, clone 1A8) was developed and characterized previously (Tobioka et al. 2002). A rabbit anti-CLDN-1 polyclonal antibody (PAb, JAY8), mouse anti-CLDN-4 MAb (clone 3E2C1), and rabbit anti-CLDN-7 PAb (ZMD241) were purchased from Zymed Laboratories (South San Francisco, CA). A rabbit anti-p63 PAb (H-137) was obtained form Santa Cruz Biotechnology (Santa Cruz, CA). ST-2G rabbit anti-p73 antibodies were described previously (Ichimiya et al. 2002). Anti-cytokeratin antibodies used were a mouse MAb (clone KL1; Immunotech, Marseille, France) and a rabbit PAb (Biogenesis; Kingston, NH). A mouse anti-CD11c MAb (clone KB90) was obtained from Dako Cytomation (Tokyo, Japan). Rabbit anti-AIRE antibodies were provided by Ismo Ulmanen (National Public Health Institute; Helsinki, Finland).
Primary Culture of Thymic Stromal Cells
Procedures used for primary thymic epithelial cell culture were previously reported (Crisa et al. 1997; Kikuchi et al. 2004). In brief, the thymus was fragmented into small pieces 2-3 mm3 in volume and washed with PBS. These tissue specimens were suspended in a dispersing solution with 0.4 mg/ml DNase I (Sigma; St Louis, MO) and 0.14 mg/ml Liberase Blendzyme 3 (Roche; Basel, Switzerland) in PBS and then incubated at 37C for 30 min with vigorous agitation. After filtration of dissociated fragments suspended in a dispersing solution with a cell strainer, cells were washed with PBS and cultured in calcium-free WAJAC404 medium supplemented with 28 mmol/liter HEPES, 14 mmol/liter sodium bicarbonate, 10 ng/ml epidermal growth factor, 10 nM dexamethasone, 50 μg/ml streptomycin, and 2% heat-inactivated human serum on collagen type I-coated dishes in a humidified atmosphere at 37C in 5% CO2. After overnight incubation, adherent cells were generously washed with PBS and then fixed in 3% paraformaldehyde in PBS for 10 min. Subsequently, cells were permeabilized in 0.5% Triton X-100-PBS for 5 min. Following this, cells were incubated in 20 mg/ml BSA-PBS for 1 hr at room temperature and washed with PBS for subsequent immunohistochemical (IHC) studies. To obtain primary DCs, adherent cells from thymic tissues dispersed with the same procedures as above were cultured in RPMI1640 supplemented with 50 μg/ml streptomycin and 2% heat-inactivated human serum on collagen type I-coated dishes in a humidified atmosphere at 37C in 5% CO2. After overnight incubation, cells were washed with PBS and then fixed with the same procedures as with primary TECs for IHC.
IHC
Procedures used for double-staining analysis of tissue sections or primary culture cells were as previously described (Ichimiya et al. 2002). Briefly, specimens were subjected to staining with a primary mouse MAb and rabbit PAb at 4C overnight. Subsequently, these were visualized with secondary goat IgG conjugated to Alexa Fluor 488 (green) or 594 (red) (Molecular Probes; Eugene, OR). Specimens were examined with a laser scanning confocal microscope (R2100AG2; Bio-Rad, Hercules, CA) or immunofluorescence microscope (IX71 inverted research microscope; Olympus, Tokyo, Japan).
Reverse Transcription (RT)-PCR and TaqMan PCR
Total RNA was isolated from the thymus and cells using TRIzol reagent (Invitrogen). First-strand cDNA synthesis and subsequent PCR to detect transcripts of CLDNs were performed as previously described (Go et al. 2004). To find transcripts of connexins, PCR reaction was conducted with specific primers as follows: connexin 26 (sense 5î-ATGGATTGGGGCACGC-3î and antisense 5î-TTAAACTGGCTTTTTTGACTTCCC-3î; amplicon length, 665 bp), connexin 32 (sense 5î-ATGAACTGGACAGGTTTGTACACCTTGCTC-3î and antisense 5î-TCAGCAGGCCGAGCAGCGG-3î; amplicon length, 822 bp), and connexin 43 (sense 5î-GGGTTAAGGGAAAGAGCGACC-3î and antisense 5î-CCCCATTCGATTTTGTTCTGC-3î; amplicon length, 227 bp). After agarose gel electrophoresis, amplified products were visualized under a UV illuminator. TaqMan PCR was performed as described in the manufacturer's protocol for Assays-on-Demand Gene Expression products (Applied Biosystems; Foster City, CA). Briefly, 50 μl of PCR reaction mixture was made from 100 ng of cDNA, Assay-on-Demand Gene Expression Assay mix, GAPDH control mix, and TaqMan Universal PCR master mix and analyzed with an ABI7000 (Applied Biosystems). To compare levels of expression of target molecules, the delta delta CT method was used to analyze triplicate specimens as described in the manufacturer's protocol (Applied Biosystems).
Freeze-Fracture Replica Electron Microscopy
The procedure for freeze-fracture replica electron microscopy was described previously (Go et al. 2004; Takano et al. 2005). Briefly, small glutaraldehyde-fixed tissue blocks of human thymus were frozen in liquid nitrogen by vigorous shaking for 5 min. Then a specimen was set and struck with a microtome knife in the evaporation chamber of the freeze-etching apparatus (JFD-7000; JEOL, Tokyo, Japan). The fracture surface of the specimen was covered with evaporated platinum. Subsequently, the tissue was covered with a uniform layer of carbon over the top of the metal at an angle, producing a template of cellular constituents. Freeze-fracture replicas were observed using a transmission electron microscope (1200EX; JEOL) at an acceleration voltage of 100 kV.
Microinjection
Microinjection into a single HTS91 cell was achieved on an IX71 inverted research microscope (Olympus) with the aid of microinjection devices (FemtoJet and InjectMan NI2; Eppendorf AG, Hamburg, Germany). The pressure of the microinjector was maintained at a constant 80-100 hPa on the tip of a finely pulled glass capillary (Sterile Femtotips II; Eppendorf AG), and the injection was done by increasing the pressure to 100-120 hPa for ~0.6 sec. After microinjection of Alexa 488 molecules, time-lapse images of signals were chased under the IX71 imunofluorescence microscope using MetaMorph imaging software (Molecular Devices; Sunnyvale, CA).
Results
Expression and Localization of OCLN in the Thymus
Epithelial integrity is generally preserved by intercellular adhesion with a junctional apparatus including TJs and desmosomes, to which intermediate filaments and actin are linked in cells, respectively (Tsukita et al. 2001). Although desmosome-associated junctions of thymic epithelial meshwork have been reported (Rimm et al. 1984; Nuber et al. 1996; Langbein et al. 2003), expression profiles and functional significance of TJs in thymic epithelium are not fully understood. To address the question of whether TJs have a role in forming the TEC meshwork, we initially conducted IHC to determine the tissue distribution of OCLN, the most reliable IHC marker of TJ in the thymus. Results showed that OCLN was expressed in stromal cells, but not in immature T cells, of medullary and subcapsular regions (Figure 1A). Next, we examined localization of OCLN and the p63 and p73 homologous to p53, because p63 and p73 are localized in the nuclei of epithelial cells within the human thymus (Di Como et al. 2002; Ichimiya et al. 2002; Kikuchi et al. 2004). By virtue of the expression profiles of p53 homologs, we considered that further analysis of these TEC subtypes would afford new insight into the mechanisms associated with construction of the stromal network. Results obtained using confocal laser microscopy showed that p63 was preferentially colocalized in the nuclei of stromal cells expressing OCLN in the medullary and subcapsular regions, indicating that epithelial cells in these regions expressed OCLN (Figure 1B). Stromal cells with OCLN in the medulla and subcapsule also appeared to express p73 in their nuclei (Figure 1C), suggesting that TJs mainly appeared in interepithelial cell connections to form mTEC and sTEC networks.
Expression and Localization of CLDN Subtypes in the Thymus
To establish TJ-mediated connections, it is well recognized that OCLN helps 22-24 kDa CLDN molecules to compose strand-like structures of TJs carried on the cell surfaces of many epithelial cell types (Tsukita and Furuse 2000; Tsukita et al. 2001). Therefore, we next investigated the expression of transcripts of CLDN subtypes by RT-PCR analysis of the thymus. In whole thymic tissues, transcripts of CLDN-1, CLDN-4, CLDN-6, CLDN-7, CLDN-8, CLDN-9, and CLDN-14 were detected (Figure 2A). Further RT-PCR analysis of TECs from primary culture showed that of these CLDN subtypes found in thymic tissues, there were transcripts of CLDN-1, CLDN-4, and CLDN-7 in primary TECs. Thus, CLDN-1, CLDN-4, and CLDN-7, as well as OCLN, of TJ-associated molecules, might participate in the formation of TJ strands to make epithelial networks in the medulla and subcapsule. To further study histological localization of CLDN-1, CLDN-4, and CLDN-7, we carried out IHC for thymic tissues to compare their distribution to that of OCLN. These OCLN subtypes were conspicuously localized in the medullary regions; CLDN-1 was broadly expressed over the medulla, whereas CLDN-4 and CLDN-7 were expressed on cells adjacent to Hassall's corpuscles (Figure 2B). In contrast to the medullary regions, expression of CLDN subtypes in cortical and subcapsular regions was undetectable. Confocal laser microscopy of the medulla revealed almost the same distribution of OCLN and CLDN-1, whereas some cells in the medulla with OCLN seemed to express CLDN-7 (Figure 2C) Thus, these findings indicated that OCLN and CLDN-1 would be a major combination in the formation of TJ strands in mTECs, whereas CLDN-4 and CLDN-7 were more limited to cells around Hassall's corpuscles in which a combination of OCLN and CLDN-7 at least was inferred. CLDN-2 was not observed in any thymic region by IHC analysis or detected in the thymus by RT-PCR analysis (data not shown).
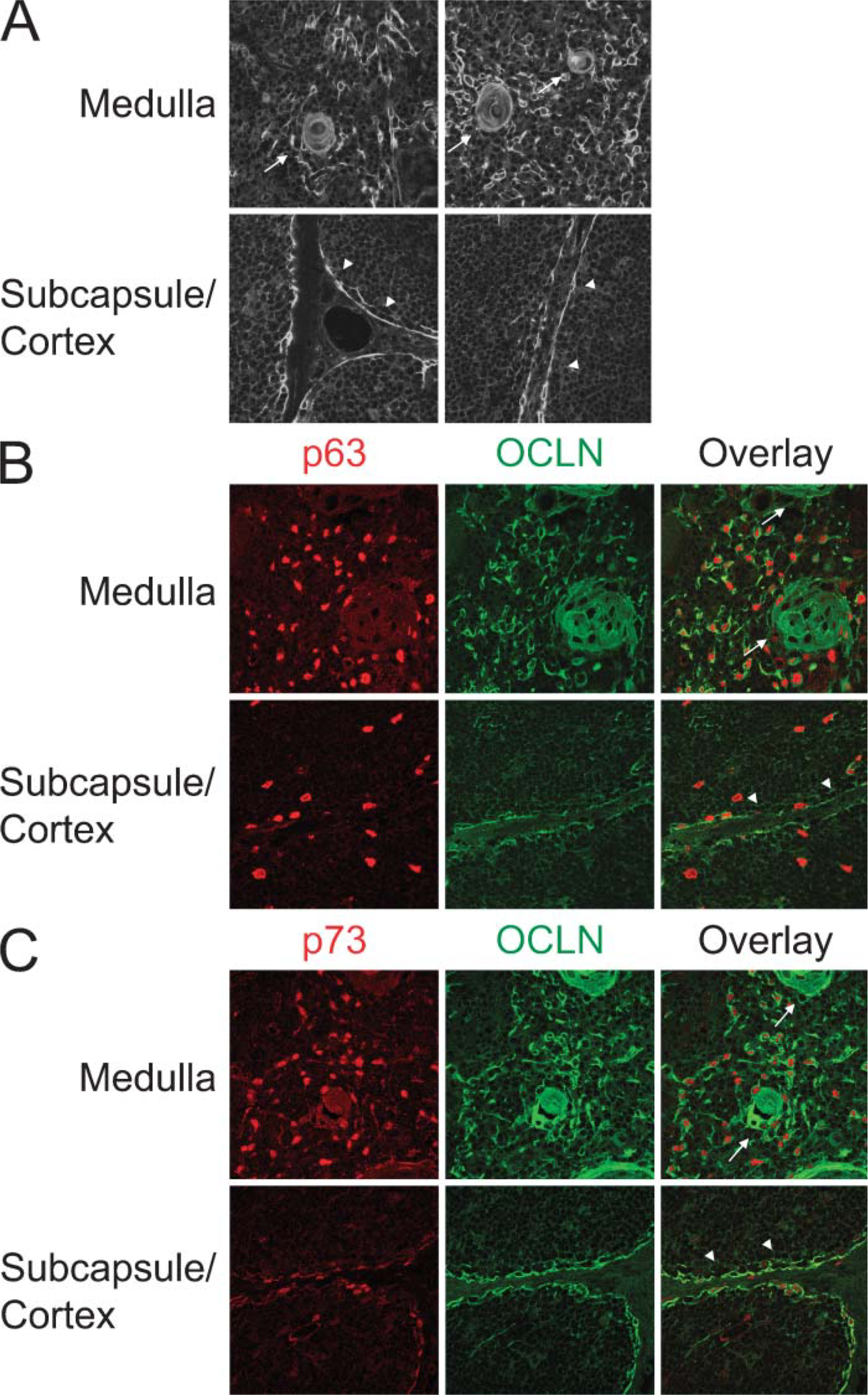
Tissue distribution of occludin (OCLN) and p53-like transcription factors in human thymic epithelial cells (TECs).
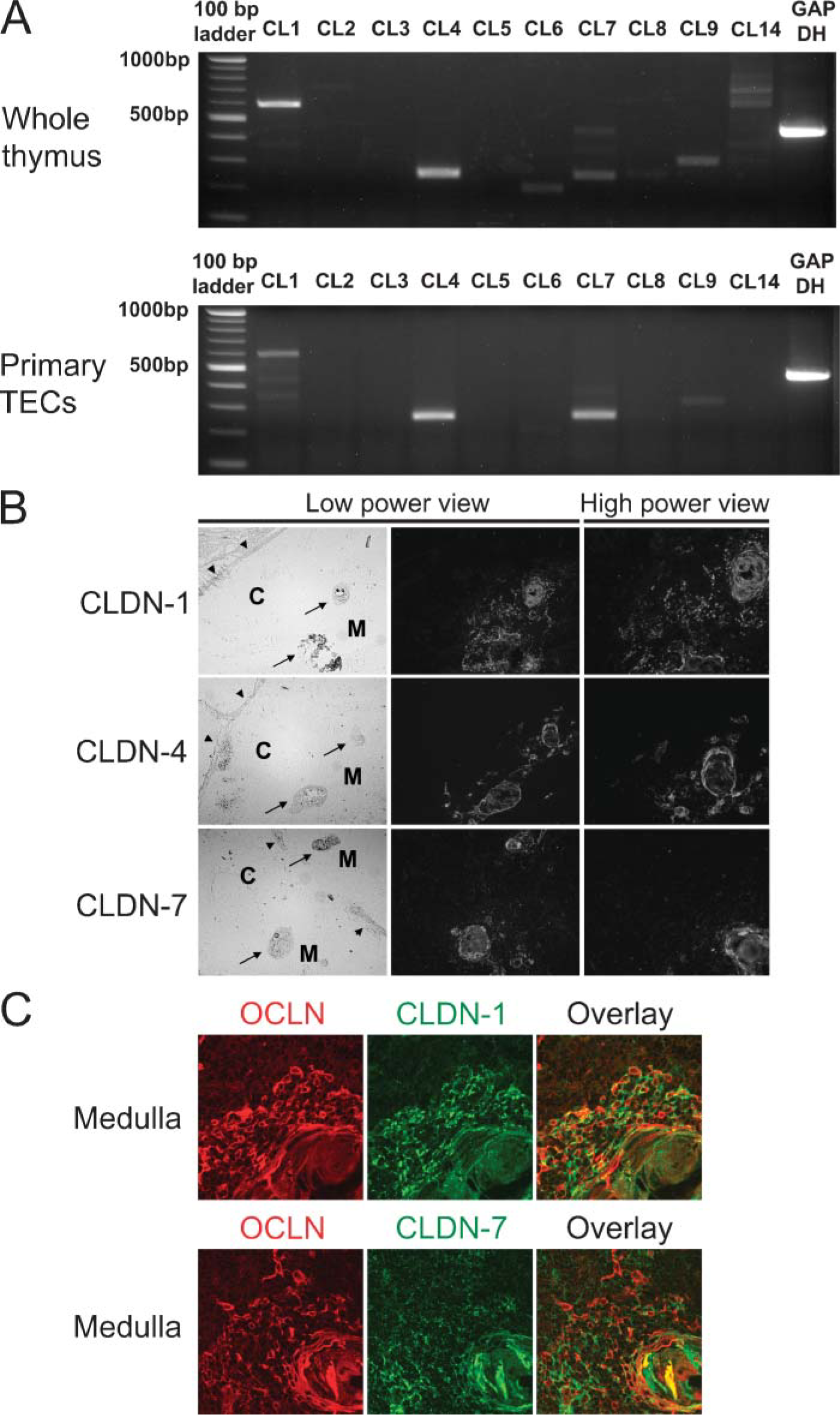
Predominant expression of claudins (CLDNs) in medullary TECs (mTECs).
Next, IHC studies of TECs from primary culture were performed to determine whether TECs really expressed CLDN-1, CLDN-4, or CLDN-7. In primary TECs attached to collagen-coated culture dishes, we could find flat polygonal cells expressing both cytokeratin and such CLDN subtypes (Figures 3A-3F). TECs positive for CLDN-1 were frequently observed to be adjacent to other TECs with the same phenotype, probably derived from part of the medullary epithelium (Figures 3A and 3B). In most of these vicinal cells, intensive signals of CLDN-1 were detected in the intercellular interface, indicating that CLDN-1 mainly participated in the formation of TJs in the medulla. Among TECs of primary culture, a single cell with cytokeratin and CLDN-4 was also occasionally found, but the cell number was smaller than for CLDN-1-positive TECs (Figures 3C and 3D). Cytokeratin-positive cells with CLDN-7 were observed in a manner similar to that of CLDN-4-positive cells (Figures 3E and 3F). These observations of primary TEC culture possibly reflect the expression profiles of CLDN-1 as well as CLDN-4 and CLDN-7 in the thymus and were limited to some of the mTECs as indicated in Figure 2.
p53-Like Transcription Factors Regulating the Expression of TJ-associated Molecules
To further investigate the possible roles of p63 and p73 in regulation of expression of TJ-associated molecules in TECs, we established human F2.5 TECs transformed by p63 or p73, whose expression in cells was assessed by quantitative PCR analysis (Figure 4A). IHC analysis of transformed F2.5 cells suggested that p63 could have the capacity to upregulate expression of CLDN-1 and CLDN-7, whereas expression of CLDN-4 and OCLN was not likely to be regulated by p63 (Figure 4B). In contrast, p73 was associated with upregulation of OCLN, but not these CLDN subtypes. This indicated that expression of TJ-associated molecules of TECs might be regulated, at least in part, by p63 and p73. Together with the tissue distribution of p63 and p73 in mTECs, these two p53-like molecules might control expression of TJ-associated molecules in mTECs.
Medullary DCs Express TJ-associated Molecules
Of further interest are recent reports describing the functional significance of TJ-associated molecules of DCs within epithelia of several mucosal tissues (Rescigno et al. 2001; Ichiyasu et al. 2004; Takano et al. 2005). In the medulla of the thymus, DCs that express CD11c at high levels display potent ability to induce deletion of autoreactive T cells prone to move immature DP-T cells into the periphery (Crowley et al. 1990; Sotzik et al. 1994). Confocal laser microscopic analysis of the medullary regions revealed that there were cells expressing both OCLN and CD11c, indicating that medullary DCs might express OCLN (Figure 5A). Similarly, CD11c-positive DCs were also found to express CLDN-1. In fact, we could observe cells with several short cytoplasmic processes that expressed both CD11c and CLDN-1 in primary culture cells derived from thymic tissues (Figure 5B). Therefore, it was considered that TJ-mediated connections were widely utilized not only for establishing epithelial meshworks but also for joining DCs to epithelial cells within the medullary regions.
Electron Microscopic Structures of TJs of the Thymus
Our observations raised the question of how the in vivo structure of TJ strands of the thymus could be recognized. Electron microscopic analysis of the freeze-fracture replicas of the thymus revealed string-like fragments of TJ strands on the surfaces of certain cell types (Figures 6A-6D). TJ strands appear as rows of particles with center-to-center spacing and usually form a continuous anastomosing network of intermembranous particle strands and complementary grooves in epithelia of tissues like skin and the intestinal tract (Staehelin 1974; Anderson 2001). When compared with TJ strands of such ordinary epithelia, the number and density of TJ strands in the thymus varied, and short stretches of TJ strands were more frequently observed. It is of interest that, as shown in Figures 6A and 6B, GJ plaques were frequently identified in the same surface planes of TJ strands as observed in many epithelial cells (Good-enough and Paul 2003). GJs act as channels to transfer small molecules between the cytoplasms of adjacent cells, including signaling molecules such as cAMP and short peptide fragments that are presentable by MHC to T cells (Neijssen et al. 2005).
Cell-Cell Transfer of Small Molecules Between Medullary TECs
As shown in Figure 6, evidence of GJ plaques near TJ strands suggested that mTECs with TJs could have a capacity for molecular transfer between TECs. RT-PCR analysis indicated the expression of connexin 43 and connexin 23 of GJ-associated molecules in thymic tissues as well as TECs of primary culture (Figure 7A). We therefore used the microinjection technique with timelapse analysis for HTS91 mTECs that expressed autoimmune regulator (AIRE) in the nuclei and connexin 43 on the cell surface (Figure 7B; Tao et al. 2006). Results indicated that Alexa Fluor 488 (molecular mass 788) was easily transferred to adjacent cells within a few minutes (Figure 7C). In another microinjection experiment, molecules such as lucifer yellow (molecular mass 341) were also transferred (data not shown). These findings imply that mTECs might have the ability to communicate with adjacent cells within the medullary epithelial network.
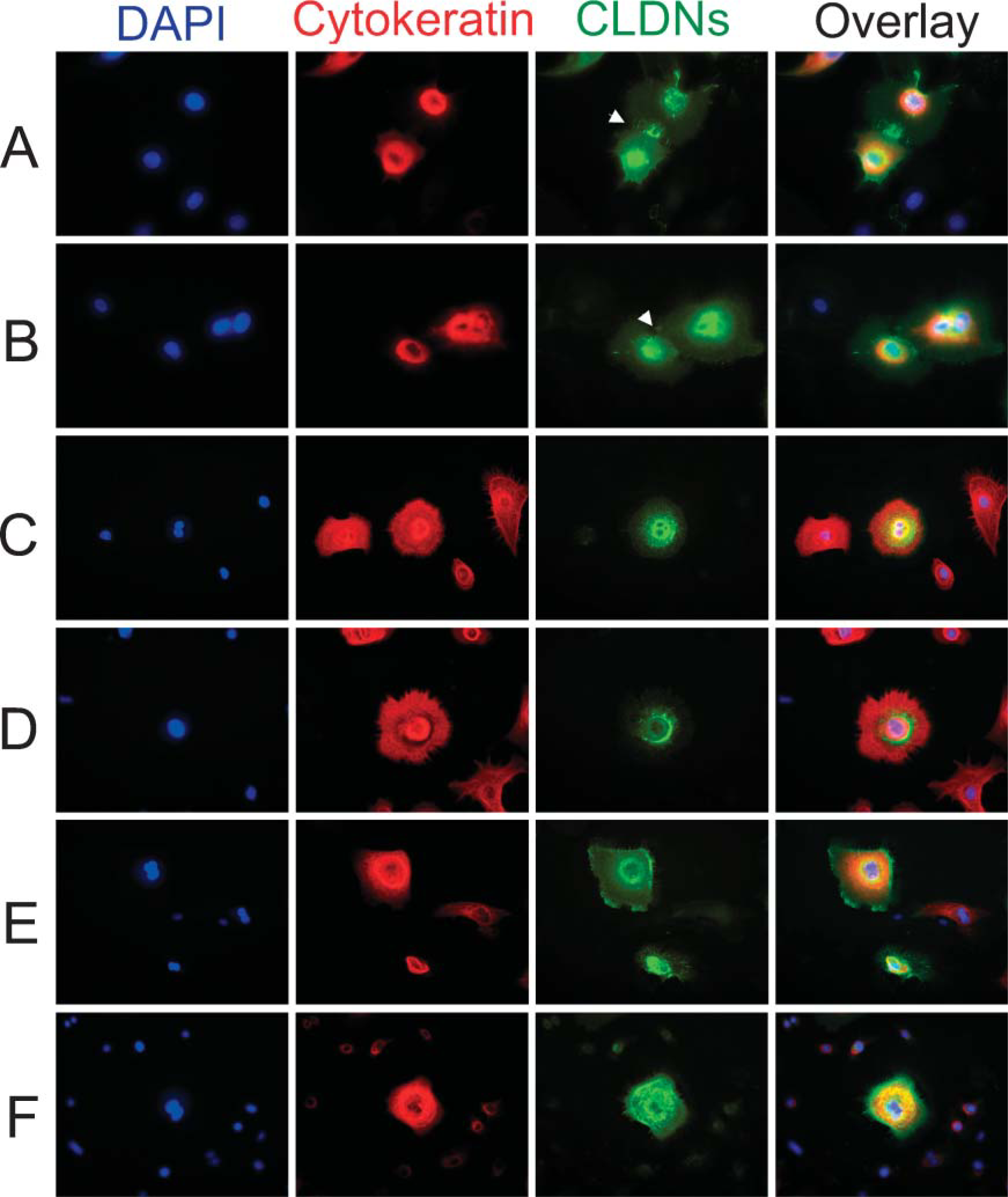
Expression of CLDNs in primary TECs.
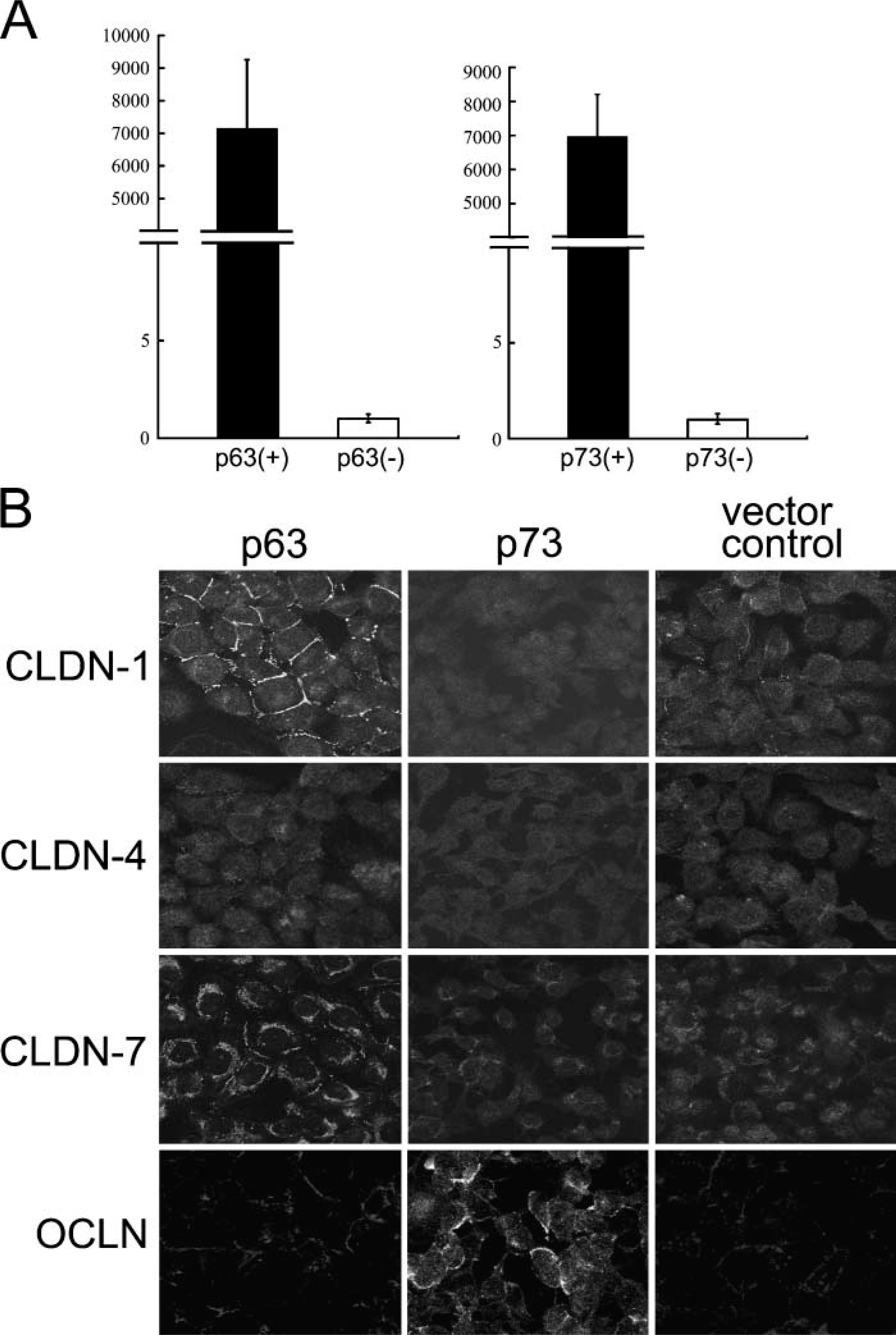
Upregulation of TJ-associated molecules by p53 family members in human TECs.
Discussion
To establish immunosurveillance mechanism supported by effector T cells, thymus generates fully functional T-cell repertoires with immunological self-tolerance properties. This central process of the thymus is mainly achieved by MHC-based negative selection of T cells in the medulla. This study provides the first evidence that p63 and p73, related to the archetypal tumor suppressor p53, participate in the regulation of CLDN-1 and OCLN of TJ-associated molecules, respectively, to help TEC-TEC and possibly TEC-DC connections in the human thymic medulla. This also provides a hint as to why the histological architecture of the thymus is recognized as two major parts: the cortex and medulla. It has been reported that p63 and p73 show epitheliotropic expression in many tissues, suggesting their fundamental roles in the maintenance of epithelial cell identity (Di Como et al. 2002; Green et al. 2003; Kamiya et al. 2004). The functional relevance of such p53 family members of the human thymus is presumed in previous reports that, for example, mutations of the p63 gene cause ectrodactyly-ectodermal dysplasia cleft lip/palate (EEC) and EEC-related syndromes, some of which exhibit histological abnormalities of the thymus and prominent depletion of cells in T-zones of the lymph nodes (Frick et al. 1997; Celli et al. 1999; Brunner et al. 2002). We have recently reported nuclear localization of p63 and p73 in human TECs, which are capable of regulating cell adhesion through ICAM-1/LFA-1 interaction (Ichimiya et al. 2002; Kikuchi et al. 2004). From the point of view of cell adhesion, it is assumed that p63 and p73 play a role in the regulation of T-cell development by supporting intercellular contacts of TECs to other cells in the medulla. The importance of p53-related molecules in the regulation of the expression of Notch receptors, Jagged-1 and Jagged-2, as suggested by studies of human cancer cell lines (Sasaki et al. 2002; Ross and Kadesch 2004), should also be noted because Notch signaling is required to induce T-cell precursors to differentiate into mature T cells in mouse models (Radtke et al. 2004; Schmitt et al. 2004).
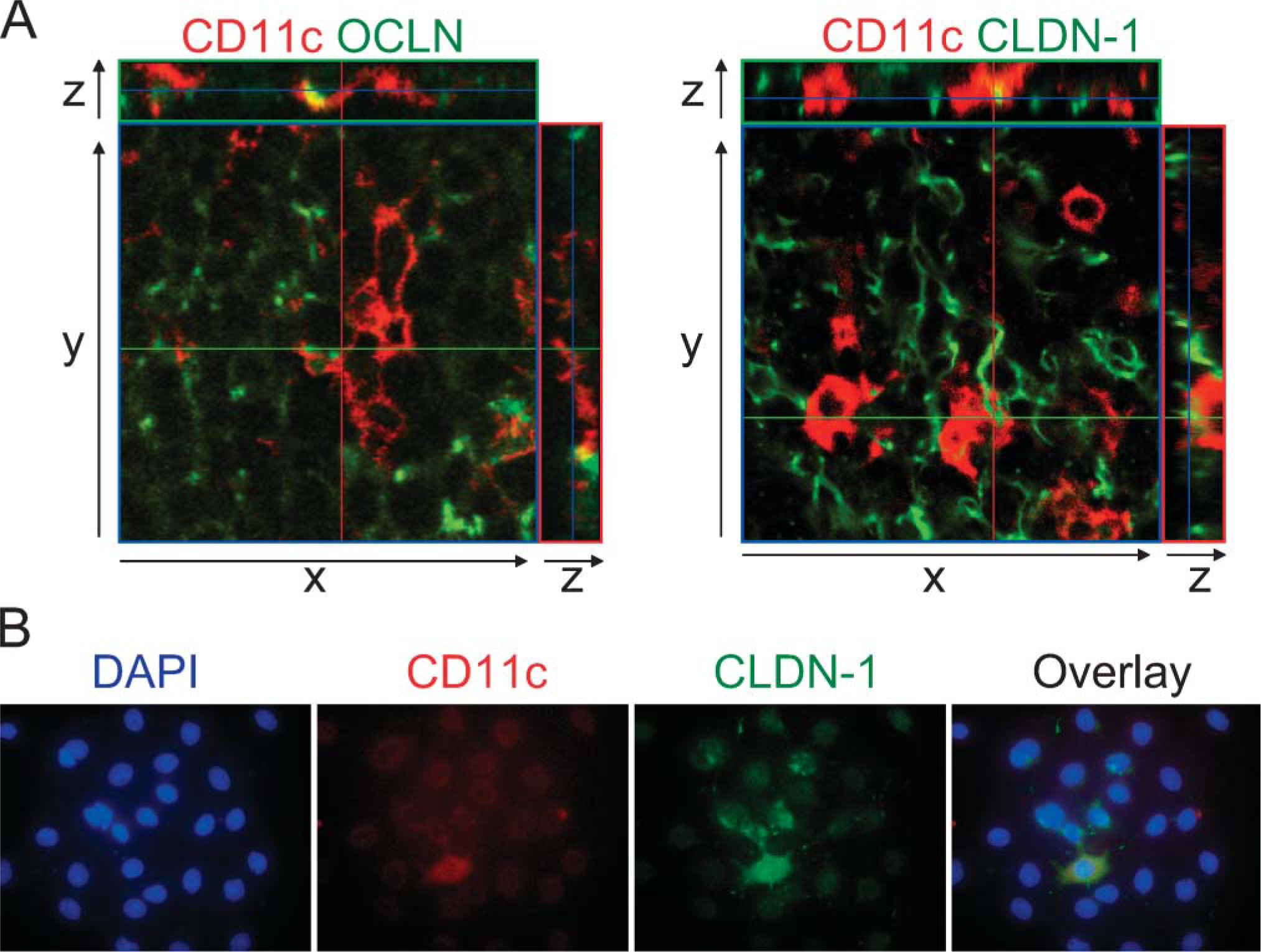
Medullary dendritic cells (DCs) expressing TJ-associated molecules.
In the formation of epithelial networks of the medulla, CLDN-1 and OCLN appear as major constituents of TJ strands, which are initially recognized as common multispanning molecules constructing TJ strands in many different types of epithelium (Tsukita et al. 2001). Therefore, the mTEC network seems to share a common epithelial architecture seen in usual epithelium. Recent studies of the mucosal immune system show that CLDN-1 and OCLN have important roles in the defense mechanism when DCs interact with epithelia of tissues in the lung, intestine, and pharynx (Rescigno et al. 2001; Ichiyasu et al. 2004; Takano et al. 2005). These findings, together with our evidence about the expression profiles of TJ-associated molecules of DCs, suggest that the TJ-mediated connection might be a general moiety to encourage interaction of DCs with epithelial cell compartments. In the cortical epithelial network we failed to detect TJ-associated molecules by our IHC procedures. It was difficult to conclude that there was localization of TJs in the cortex from histological analysis of thymic sections because intercellular contact points between cell surfaces of cTECs were obscured compared with those of mTECs and cTECs due to the cell density and longer cytoplasmic process of cTECs used to achieve epithelial-lymphocyte interactions.

Freeze-fracture replica electron microscopy of the thymus.
Whereas CLDN-1 was expressed in most mTECs, CLDN-4 and CLDN-7 were localized in some mTECs. These observations support the heterogeneity of mTECs and further focus attention on the complexity of the mTEC network. It is well known that mTECs are heterogeneous groups divided into different subpopulations based mainly on ultrastructural morphology (van de Wijngaert et al. 1984; von Gaudecker 1991). There must be a molecule(s) responsible for generating mTEC heterogeneity, although mTECs broadly express both p63 and p73 transcription factors. In accordance with the analysis of p73-transformed TECs suggesting a role of p73 in the regulation of OCLN, OCLN was preferentially localized in medullary and subcapsular epithelia with p63 and p73. However, the expression profile of CLDNs of mTECs was clearly different from that of sTECs. The association of heterogeneous CLDN subtypes with OCLN constitutes the backbone of TJ strands in situ so that TJ strands are recognized as copolymers of OCLN and heterogeneous CLDNs (Green et al. 2003). In terms of the TJ-mediated TEC connection, it might be possible that p73 participates in the regulation of OCLN, whereas CLDNs are under the control of p63 and other factors.
Intercellular communication is canonically achieved by GJs via mediation of transportation of small molecules. On the cell surface of general epithelium, TJs and GJs frequently form a junctional complex. In cells, TJ-associated molecules directly interact with zonula occludens-1 (ZO-1), which is a member of the family of proteins called membrane-associated guanylate kinases. In addition to colocalization of TJs and GJs, there is accumulating evidence that ZO-1 controls connexin 43-mediated communication (Hunter et al. 2005; Laing et al. 2005). Thus, TJs of the medulla might play a role not only in establishing stromal cell connections but also in helping to achieve GJ-mediated intercellular communication as in other epithelial tissues. Connexin 43, which is known to be a widely distributed molecule, transports calcium ions and other signaling molecules such as cyclic AMP, cyclic GMP, and inositol 1,4,5-triphosphate. Even short peptide fragments are translocated through connexin 43 in the tumor cell network against immunosurveillance by cytotoxic T cells (Neijssen et al. 2005). The fact that small dye molecules were readily transferred to neighboring cells of human mTECs probably supports the existence of intercellular communication within the mTEC network. However, further investigations are required to address the question of whether the mTEC network shares self-peptides for their presentation to immature T cells.
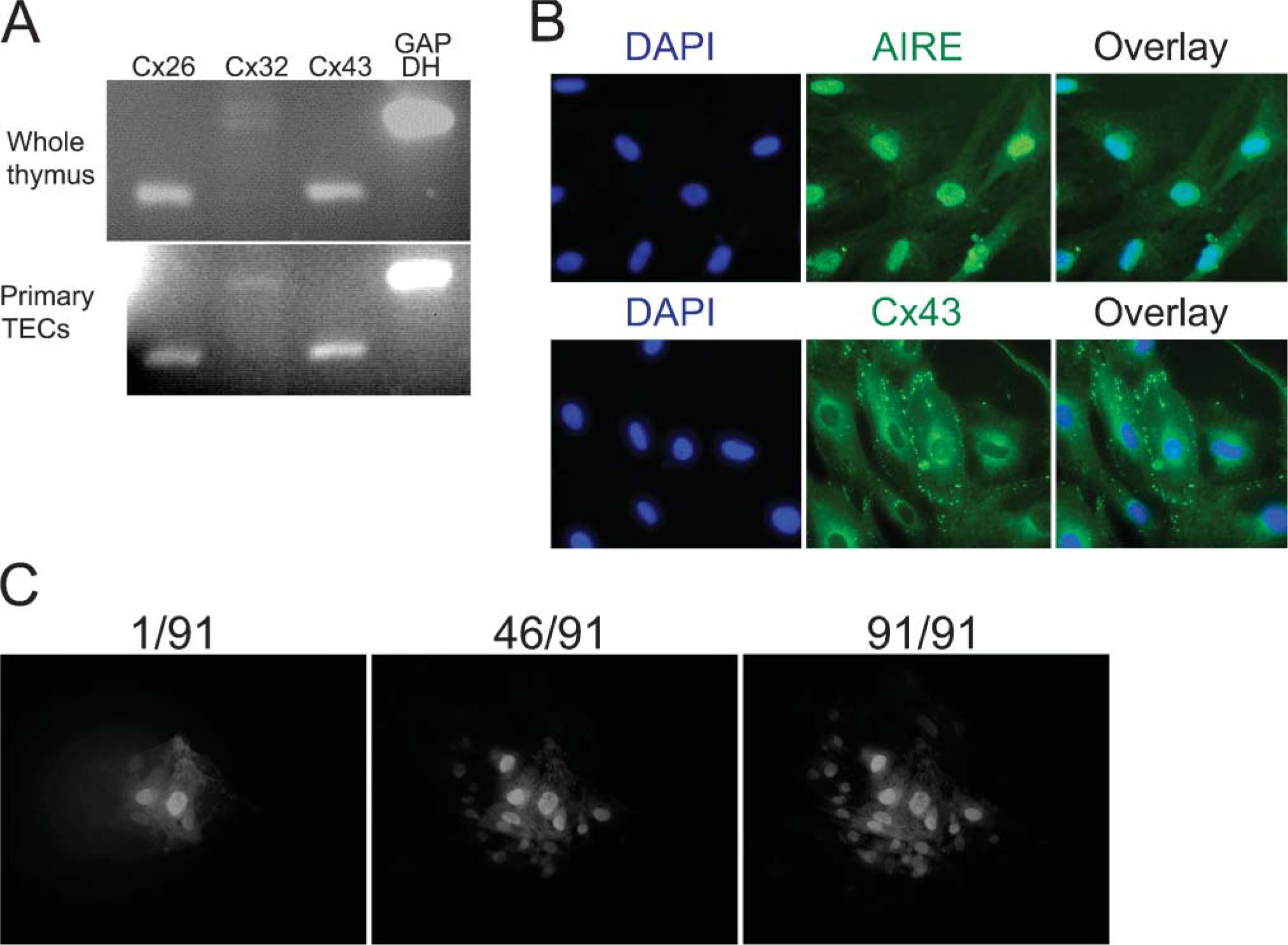
Small molecules transferring HTS91 mTECs with GJ-associated molecules.
In summary, this study demonstrates that p53 homologous transcription factors take part in the regulation of TJ-mediated connection to form the TEC meshwork of the human thymic medulla. Within the network, DCs are also connected through TJs, probably to perform their important role in thymic selection. At present, we cannot fully explain the reason why p63 is involved in such an anomaly associated with failure of T-cell development. However, our findings imply a role for p53 family members in the maintenance of the mTEC scaffold required for establishment of the microenvironment for human T-cell selection.
Footnotes
Acknowledgements
This work was supported by a Grant-in-Aid from the Japan Society for the Promotion of Science for Scientific Research (C16590286 to SI).
The authors thank Sachiko Kimura and Shigeaki Yokoyama (Hokkaido Children's Hospital and Medical Center, Hokkaido, Japan) for thymic tissues, María L. Toribio (Universidad Autónoma de Madrid, Madrid, Spain), and Laura Crísa (Scripps Research Institute, La Jolla, CA) for cell lines, Gerry Melino (University of Rome ‘Tor Vergata’, Rome, Italy) for plasmid DNAs and Ismo Ulmanen (National Public Health Institute, Helsinki, Finland) for AIRE antibodies. The authors also thank Jacques Neefjes (The Netherlands Cancer Institute, Amsterdam, The Netherlands) for instruction on microinjection techniques.