
Abstract
Using confocal microscopy and morphometry, we analyzed the expression of connexin26 (Cx26) and ZO-1 in rat cochlea during the postnatal period to elucidate spatiotemporal changes in gap junctions and tight junctions during auditory development. We also studied changes in these junctions in experimental endolymphatic hydrops in the guinea pig. In the adult rat cochlear lateral wall, Cx26 was detected in fibrocytes in the spiral ligament and in the basal cell layer of the stria vascularis, whereas ZO-1 was detected in the apical surfaces of marginal cells and in the basal cell layer. During postnatal development, Cx26 expression increased mainly in the spiral ligament, whereas ZO-1 expression increased in the basal cell layer. The morphometry of Cx26 showed a sigmoid time course with a rapid increase on postnatal day (PND) 14, whereas that of ZO-1 showed a marked increase on PND 7. In experimental endolymphatic hydrops, the expression of Cx26 significantly decreased, whereas there were no obvious changes in the expression of ZO-1. These results indicate that gap junctions and tight junctions in the cochlea increase in a different spatiotemporal manner during the development of auditory function and that gap junctions and tight junctions in the cochlea are differentially regulated in experimental endolymphatic hydrops.
(
Keywords
T
Gap junctions form intercellular channels and mediate direct transfer of low molecular weight (less than about 1000 Daltons) metabolites, ions, and small molecules between neighboring cells. Because some second messengers in signal transduction can pass through gap junction channels, gap junctional intercellular communication is considered to play an important role in tissue homeostasis, growth control, and cell differentiation (Paul 1995; Bruzzone et al. 1996; Goodenough et al. 1996; Kumar and Gilula 1996). Gap junctional channels are composed of hexagonal arrangements of oligomeric proteins called connexins (Cxs) (Beyer et al. 1990). Thus far, more than 15 rodent genes for connexins have been cloned and shown to be expressed in a tissue- or cell-specific manner (White and Paul 1999).
The tight junction is another intercellular junction that plays two distinct roles in selective permeability barrier functions. One is as a barrier that seals neighboring cells together in a sheet to prevent leakage of molecules between them, and the other is to prevent diffusion of membrane proteins, including ion channels and glycolipids, between the apical and basolateral regions of the plasma membrane. Several proteins associated with tight junctions have been identified. These include not only the integral proteins claudin and occludin but also the peripheral proteins ZO-1, ZO-2, and symplekin (Anderson and Van Itallie 1999; Tsukita and Furuse 2000).
Gap junctions and tight junctions were detected in the cochlea by electron microscopy (Jahnke 1975; Reale et al. 1975; Nadol et al. 1976; Forge 1984; Spicer and Schulte 1996), and the immunolocalization of connexin26 (Cx26) in the cochlea has been reported (Kikuchi et al. 1995). The characteristics of electric intercellular coupling in the organ of Corti have been studied during the past two decades (Santos–Sacchi 2000). However, the crucial roles of intercellular junctions in auditory functions were not proved until a mutated Cx26 gene was revealed to be one of the genes responsible for human hereditary nonsyndromic sensorineural deafness (Kelsell et al. 1997). The discovery of mutations in Cx26 genes in human hereditary deafness is evidence of a critical role of gap junctions in hearing. However, many important questions about the roles of intercellular junctions in auditory function remain to be clarified. For example, only a limited amount of information exists on when and between what cells gap junctions and tight junctions develop in the cochlea during the postnatal development of endocochlear potential. There have only been a few recent papers on the rat (Lautermann et al. 1999), mouse (Xia et al. 1999), and gerbil (Souter and Forge 1998). This is because simultaneous observation of gap junctions and tight junctions in a wide area of the cochlea was not done. Furthermore, only a few reports are available on what changes in gap junctions and tight junctions take place in the cochlea in patients who suffer from hearing loss, such as Meniere's disease (Jahnke et al. 1985; Ichimiya et al. 1994; Nadol et al. 1995). Comparison of the changes in gap junctions and tight junctions between normal auditory development and hearing loss provides a valuable opportunity for us to understand their roles in physiological and pathological conditions.
In this study we performed confocal and morphometric analyses on the expression and localization of Cx26 and ZO-1 in the cochlea during the postnatal period and in an experimental model of endolymphatic hydrops to elucidate spatiotemporal changes in cochlear gap junctions and tight junctions not only during normal development of auditory function but also in the pathological condition. For the developmental study, we used a rat species in which the endocochlear potential has been shown to dramatically increase during the postnatal period, especially from 11–14 days after birth (Bosher and Warren 1971; Rybak et al. 1992). Guinea pigs, in which endolymphatic hydrops was experimentally induced by endolymphatic duct and sac obliteration, were used for the hydrops model.
Materials and Methods
Animals
Postnatal Development. A total of 35 Wistar rats free of middle ear infection were used for assessment of developmental changes in gap junctions and tight junctions in the cochlea. Five rats were sacrificed at each of the following postnatal days (PNDs): 1, 4, 7, 11, 14, 21, and 70, under deep pentobarbital anesthesia. The animals were perfused transcardially with freshly prepared 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4, 4C). Temporal bones were removed bilaterally and the cochleas were dissected from them and placed in the same fixative at 4C for 3 hr. The cochleas were then decalcified in 0.12 M EDTA at room temperature (RT) for a week. After washing in PBS, pH 7.4, the cochleas were rinsed as follows: (a) 10% sucrose in PBS for 3 hr; (b) 15% sucrose in PBS for 3 hr; (c) 20% sucrose in PBS for 3 hr. The specimens were infused with Tissue-Tek OCT compound (Miles; Elkhart, IN), frozen in disposable vinyl specimen molds (Tissue-Tek Cryomold; Miles) by partial immersion in liquid nitrogen, so that the axis of the cochlea was parallel to the bottom of the mold, and stored at −80C. For comparison between immunohistochemistry of ZO-1 and that of occludin, adult rat cochlear lateral walls were frozen without fixation and decalcification.
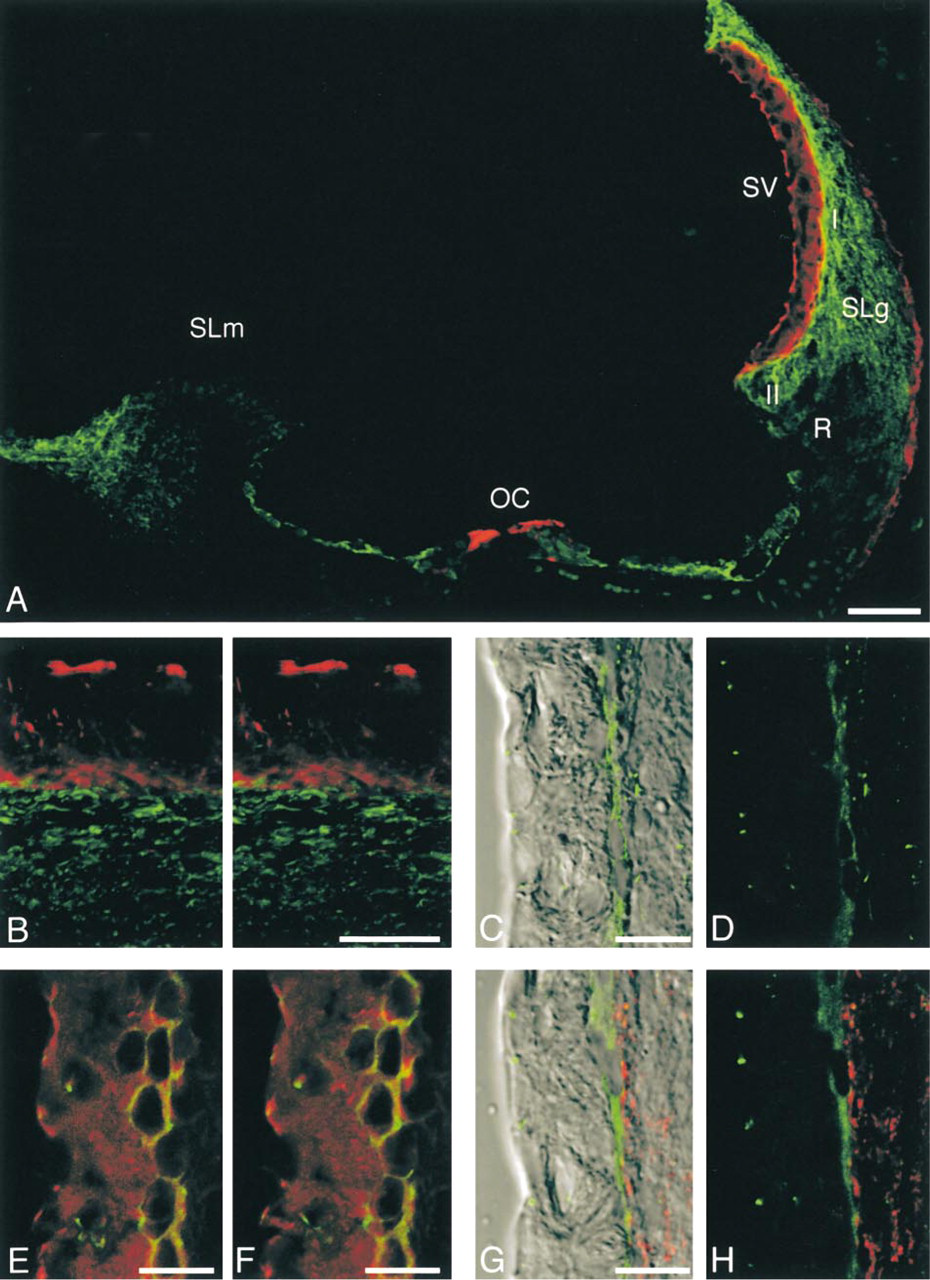
Immunohistochemical localization of Cx26, ZO-1 and occludin in the adult (70-postnatal-day) rat cochlea. (
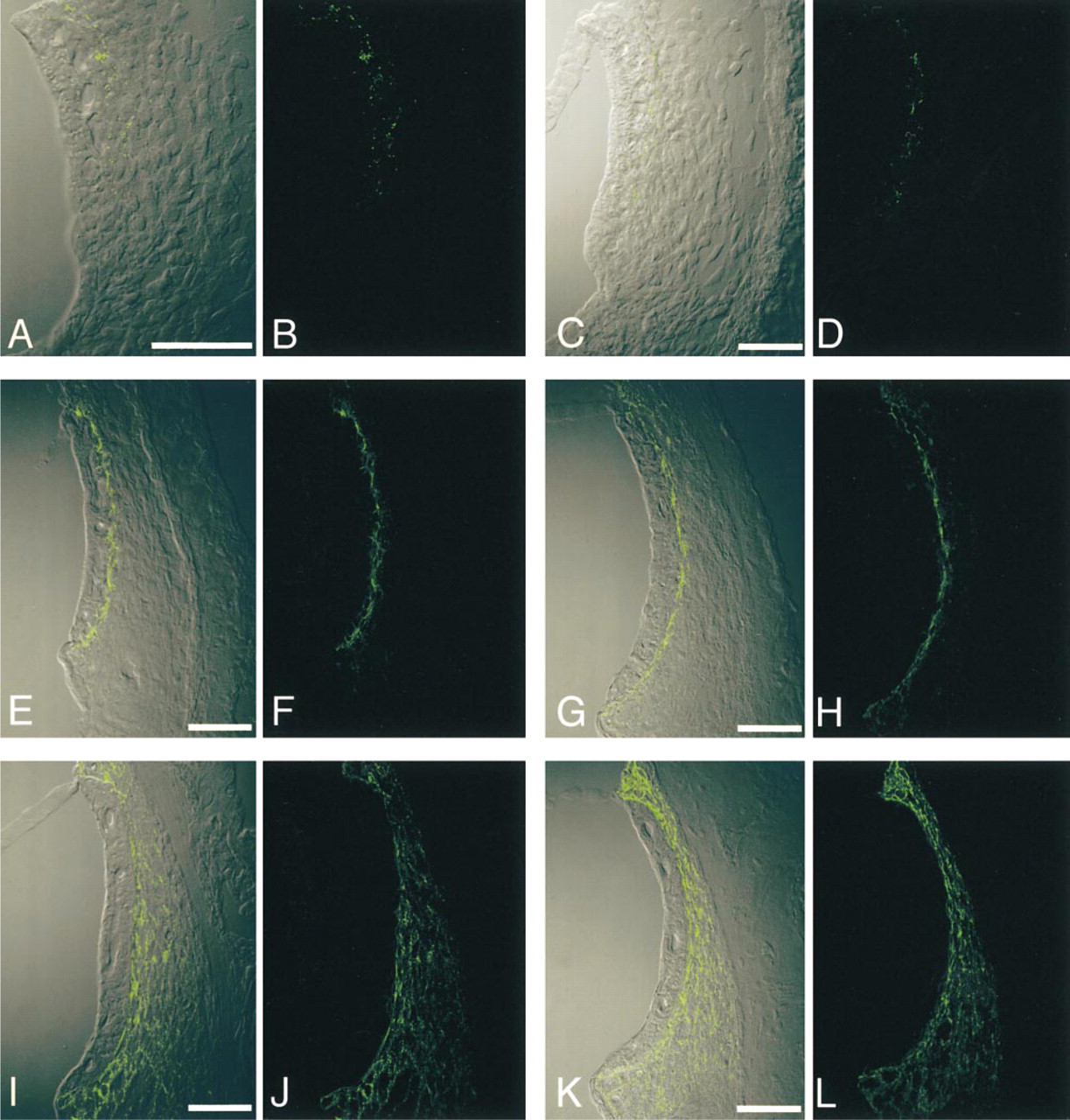
Immunohistochemical localization of Cx26 in the rat cochlear lateral wall at various postnatal days (PND). Confocal images of Cx26 with DIC imaging (
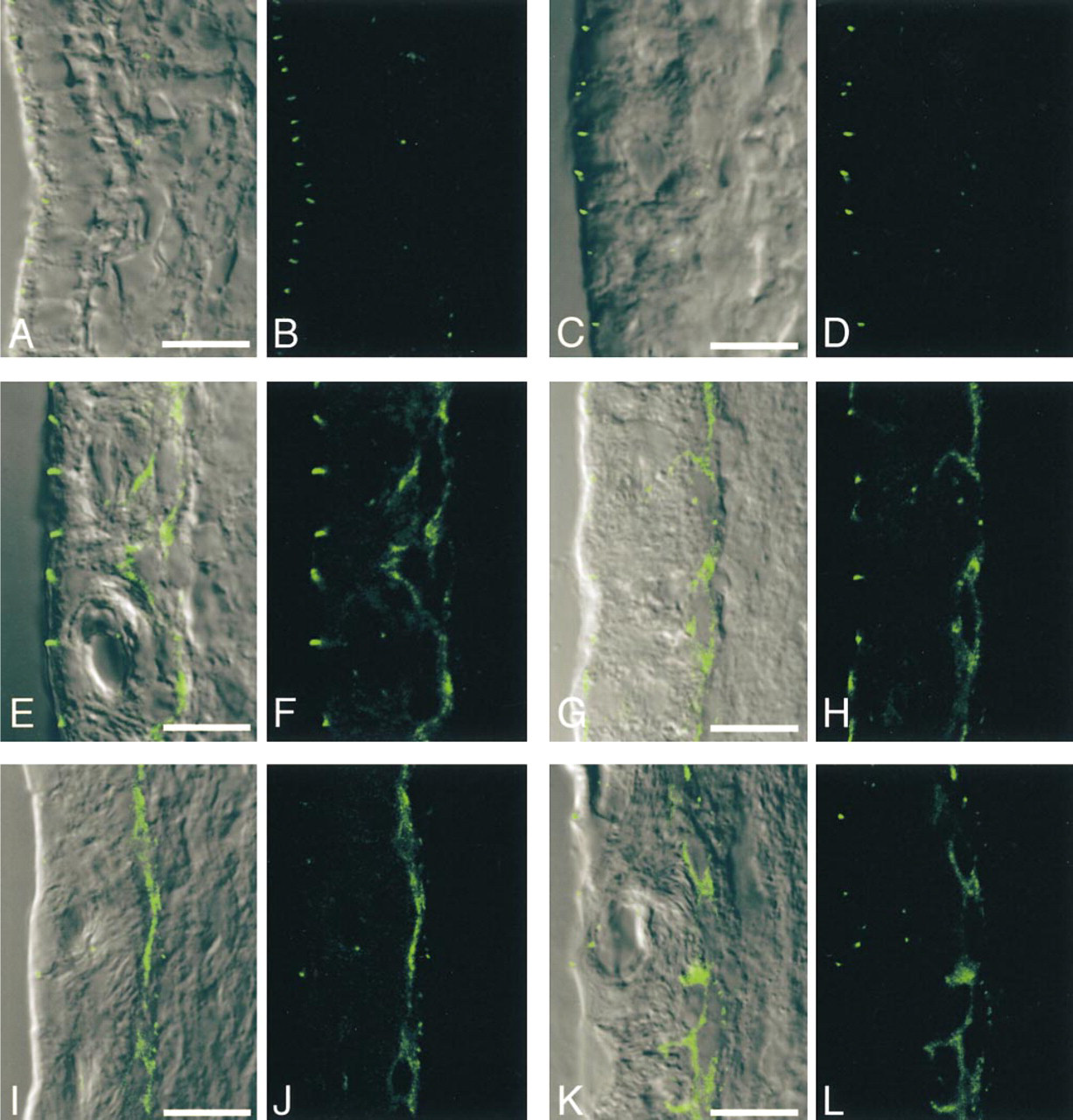
Immunohistochemical localization of ZO-1 in the rat cochlear lateral wall at various postnatal days (PND). Confocal images of ZO-1 with DIC imaging (
To confirm the relationship between maturation of endocochlear potential, and development of gap junctions and tight junctions, neonatal and adult guinea pigs, in which species endocochlear potential is mature at birth (Raphael et al. 1983), were used. Five albino guinea pigs at PND 1 and five adult albino guinea pigs at PND 70 were sacrificed under deep pentobarbital anesthesia and the cochlea specimens were prepared and stored at − 80C in a way similar to that used for the rats, except for 3-week decalcification in 0.12 M EDTA at RT.
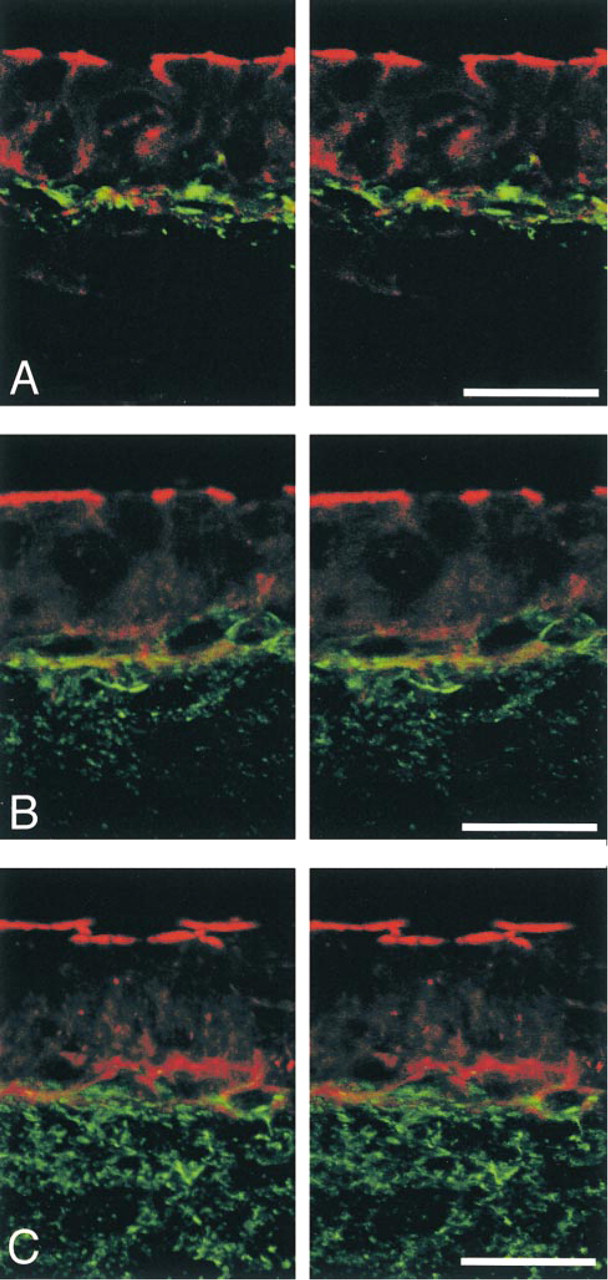
Stereoimages of double-labeling for Cx26 and F-actin in the rat cochlear lateral wall at various postnatal days (PND). (
Experimental Endolymphatic Hydrops
Three albino guinea pigs weighing approximately 300 g were used. Experimental endolymphatic hydrops was induced by endolymphatic duct and sac obliteration as described by Kimura and Schuknecht (1965). Under pentobarbital anesthesia (35 mg/kg body weight), the endolymphatic duct of the right ear of each animal was identified by drilling down from the posterior fossa and the duct and sac were blocked with bone wax. The left ear of each animal served as an unoperated control.
At 3 months after the operation, brainstem-evoked responses were recorded. Under pentobarbital anesthesia, the head was stabilized by a clamp and a sound source was placed in the external auditory meatus. All recordings were made in an electrically shielded soundproof chamber. Evoked responses were recorded between a needle electrode placed into the skull at the vertex and in the pinna of the test ear. A needle electrode in the contralateral pinna served as a ground. Thresholds were measured for clicks and tone pips. The tone pips were presented with a two-cycle rise and fall time, with no plateau, at 1, 2, 4, and 8 kHz. At each frequency the threshold was separately measured for the unoperated left and then the operated right ear. Stimulus intensities were decreased in 5 dB steps until no responses could be detected visually, and the threshold was defined as the minimal intensity at which a response was evoked. After recording, the animals were sacrificed in the anesthetized condition. Cochlea specimens from not only the experimental ears on the right side but also the left control ears were prepared and stored at −80C in a way similar to that used for the rats, except for 3-week decalcification in 0.12 M EDTA at RT. All animal experiments described in this study adhered to the standards detailed in “Principles of Laboratory Animal Care” (NIH publication no. 85 23, revised 1985), and also to the “Rules and Regulations of Animal Research, Kyoto Prefectural University of Medicine.”
Immunohistochemistry
Serial 10-μm frozen midmodiolar sections were cut with a cryostat microtome (Bright 5030 microtome), placed on silane-coated slides (Matsunami; Osaka, Japan), air-dried, and fixed in absolute ethanol at −20C for 15 min. After incubation in PBS with 5% skim milk for 15 min at RT to block nonspecific labeling, primary antibodies, such as a rabbit polyclonal antibody against a Cx26-specific peptide (amino acid residues 101–119; Kuraoka et al. 1993) (1:2000 dilution), another rabbit polyclonal antibody against Cx26 (1:200 dilution; Zymed Laboratories, South San Francisco, CA), a rabbit polyclonal antibody against ZO-1 (1:200 dilution; Zymed), a cytoplasmic protein associated with the tight junction, or a rabbit polyclonal antibody against occludin (1:100 dilution; Zymed), a tight junction-specific integral protein, were applied overnight at 4C. After three 5-min washes with PBS, incubation with fluorescein isothiocyanate (FITC)-labeled anti-rabbit antibodies (1:200 dilution; DAKO, Copenhagen, Denmark) was performed for 1 hr at 37C. Following the final rinses in PBS, sections were mounted in Vectashield (Vector Laboratories; Burlingame, CA). All antibody dilutions were made in PBS containing 1% bovine serum albumin (Sigma; St Louis, MO) and 0.1% sodium azide with 0.1% Triton X-100. In some sections, Texas Red-X-labeled phalloidin (1:200 dilution; Molecular Probes, Eugene, OR) was used together with one of the first antibodies to visualize filamentous actin (F-actin). Some other sections were double-labeled for Cx26 and ZO-1 by utilizing the mouse anti-Cx26 (1:200 dilution; Zymed) and the rabbit anti-ZO-1 antibodies in combination with FITC-conjugated anti-rabbit immunoglobulin (used as above) and Texas Red-conjugated anti-mouse immunoglobulin (1:200 dilution; Vector). As negative controls, the first antibodies were omitted under otherwise similar conditions for immunofluorescence.
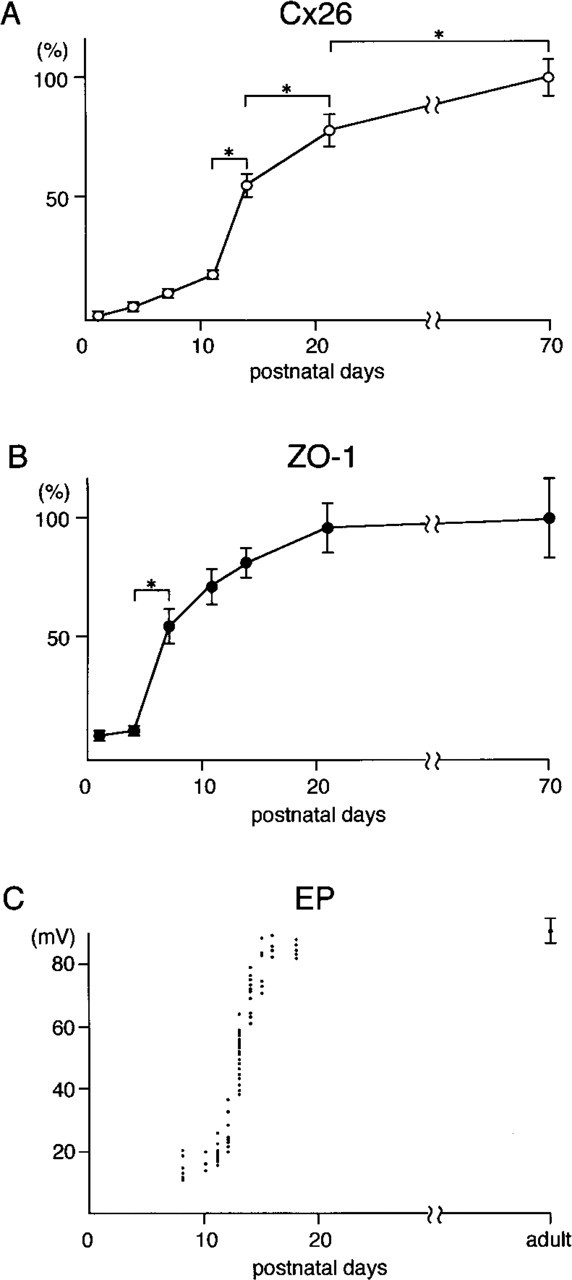
Morphometric data on the expression of Cx26 (
Confocal Laser Scanning Microscopy
The sections were observed with a confocal laser scanning microscope (Olympus FLUOVIEW system; Olympus, Tokyo, Japan) equipped with an oil immersion objective (Plan Apo ×60, NA = 1.4; Olympus). An argon–krypton laser produced excitation bands at 488 nm for FITC, 568 nm for Texas Red, and monochromatic light for differential interference contrast (DIC) images. Fluorescence images were collected with emission filters for 510–550 nm for FITC and 585–610 nm for Texas Red. Simultaneous images (800 × 600 pixels, 12 bits each) of the FITC label and either DIC images or Texas Red label were acquired and stored. Digital images were transferred to a Macintosh computer (Apple Computer) equipped with the Photoshop Program (Adobe System; Tucson, AZ) and printed on an ink-jet printer (PM-750C; Epson, Tokyo, Japan)
Morphometric Analysis
For morphometric analysis, all aspects of tissue processing, immunolabeling, and imaging were strictly standardized. For example, only the sections through the axis of the cochlea were utilized, and to exclude the possibility that variations in immunolabeling at different dates affected the morphometric data, one section each for PND 1, 4, 7, 11, 14, 21, and 70 was included in a single immunolabeling run and all of the sections were analyzed at the same time. Digital images were collected using the ×60 objective lens and a zoom setting of 2 under the same conditions, i.e., laser power = 35 mW, pinhole size = 100 μm, ND filter = 20%, photomultiplier voltage = 600 V, and scanning speed = 8.7 sec/frame. As a result, all digital images showed that the spectrum of label intensities was within the 4096-level scale. Three section planes at 1-μm steps along the z-axis were obtained for the same x–y axis.
Concerning Cx26 expression in the cochlear lateral wall, to cover the total cross-sectional area of the stria vascularis and spiral ligament in the second turn of the cochlea, two to four 800 × 600-pixel images were taken by moving slides in the x–y axis, and 6–12 images were acquired for each slide. A single set of images that were taken at the same z-axis and covered the total cross-sectional area of the stria vascularis and spiral ligament was then selected. The digital images were transferred to a Macintosh computer, converted into 8-bit images (256 levels), and analyzed using the NIH Image program (Wayne Rasband, NIH). After the area of the spiral ligament was enclosed manually, a binary overlay was created automatically by a set threshold of 40 on the 255-point gray scale to eliminate the background cell outlines. Then the binary overlaid area in the area of interest was analyzed. Data were first expressed as mean ± SE of overlaid areas (pixel × pixel) per areas of spiral ligament (pixel × pixel) and then converted so that the mean of each datum on PND 70 and of the control ear was 100% for the developmental study and experimental endolymphatic hydrops, respectively. Data were collected from five cochleas of different rats for each PND and from three cochleas of different guinea pigs for experimental endolymphatic hydrops. A total of 41 cochleas were actually analyzed.
Regarding ZO-1 expression in the strial basal cell layer, to cover the total cross-sectional area of the stria vascularis in the second turn of the cochlea, two 800 × 600-pixel images were taken. The same procedure was performed for the transfer and conversion of image data as for Cx26. Morphometric data were first expressed as mean ± SE of ZO-1-positive areas (pixel × pixel) per length of boundary lines (pixel) between the stria vascularis and spiral ligament and then converted so that the mean of each datum on PND 70 and of the unoperated ear was 100% for the developmental study and experimental endolymphatic hydrops, respectively.
Statistical analyses were performed using the StatView program (Abacus Concepts). Data on the rat postnatal development were examined by ANOVA and the Tukey-Kramer test. Data on the guinea pig between PND 1 and PND 70 were compared by t-test. Data on experimental endolymphatic hydrops were examined by paired t-test. Significant differences were defined by p > 0.05.
Results
Cx26 and ZO-1 in Adult Rat Cochlea
Double-labeling for Cx26 and filamentous actin using the anti-Cx26 antibody (Kuraoka et al. 1993) and Texas Red-phalloidin showed that, in the adult rat cochlea (PND 70), Cx26-immunolabeled spots were localized in the cochlear lateral wall, spiral limbus, and supporting cells of the organ of Corti (Figure 1A). In the cochlear lateral wall, Cx26 immunolabeling was detected in the spiral ligament and in the basal cell layer of the stria vascularis. In the spiral ligament, intense Cx26-labeling was localized among fibrocytes beneath the stria vascularis (Type I fibrocytes), infrastrial (Type II fibrocytes), and suprastrial fibrocytes (Type V fibrocytes), whereas lower expression was found among inferior fibrocytes (Type III and Type IV fibrocytes). At higher magnification, Cx26-immunolabeled spots were localized at the lower surfaces of basal cells of the stria vascularis, i.e., between basal cells of the stria vascularis and fibrocytes in the spiral ligament, as well as in the spiral ligament (Figure 1B). The anti-Cx26 antibodies showed the expected punctate labeling of gap junction plaques in adult rat livers (data not shown) that were processed in a similar manner, i.e., transcardial perfusion-fixation, decalcification, and immunolabeling. Identical results were obtained with another rabbit polyclonal antibody against Cx26-specific peptide (Zymed) (data not shown). In the absence of the first antibody, nonspecific fluorescence was barely detected in the adult rat cochlea (data not shown).
In the lateral wall of the adult rat cochlea, immunolabeling for ZO-1 was found mainly in two regions, the apical surfaces of the marginal cells and the basal cell layer (Figures 1C and 1D). In midmodiolar sections, ZO-1-positive spots were localized at the most apical areas of the marginal cells. In the surface preparation, in which the surfaces of the marginal cells were viewed from the inside of the endolymphatic space, hexagonal or hexagonal-like labeling for ZO-1 was observed (data not shown). At the basal cell layer, ZO-1-immunolabeled spots were tightly packed in a linear manner. In addition, small ZO-1-positive spots were found in the vessels and among fibrocytes. The localization of ZO-1 (Figure 1E) in the cochlear lateral wall was similar to that of occludin (Figure 1F), a tight junction-specific integral protein, when serially sliced sections were immunolabeled for ZO-1 and occludin.
To elucidate the localization of Cx26 and ZO-1 in the border zone of the stria vascularis and the spiral ligament in more detail, we performed double-labeling for Cx26 and ZO-1. Double-labeling for Cx26 and ZO-1 showed that, in the adult rat cochlea, they were closely associated at the lower surfaces of basal cells of the stria vascularis, and that Cx26-positive spots were localized at the side of the spiral ligament, whereas ZO-1-positive spots were distributed at the side of the stria vascularis (Figures 1G and 1H).
Developmental Changes in Cx26 and ZO-1 in Rat Cochlea During the Postnatal Period
On PND 1, the morphology of the rat cochlear lateral wall was different from that of the adult, i.e., the boundary between the stria vascularis and spiral ligament was ill-defined because the proper alignment of the basal cells was absent. Cx26-immunolabeled spots were localized in the border zone between the stria vascularis and spiral ligament and the number was small (Figures 2A and 2B). Immunolabeling for ZO-1 at the apical surface of the marginal cells was detected on PND 1, and the pattern did not change much during postnatal development (Figure 3). On the other hand, in the basal cell layer, only a few ZO-1-positive spots were found (Figures 3A and 3B).
On PND 4, cells at the border region between the stria vascularis and spiral ligament showed a tendency to align, but there was no distinct separation of basal cells and fibrocytes. The localization of Cx26-immunolabeled spots was still limited to the border zone between the stria vascularis and spiral ligament (Figures 2C and 2D). In the basal cell layer, only a few ZO-1-positive spots were found (Figures 3C and 3D).
On PND 7, the alignment of basal cells became clearer and the density of fibrocytes appeared to increase. The distribution of Cx26-immunolabeled spots barely changed and was restricted to the border region (Figures 2E and 2F). Double-labeling for Cx26 and F-actin showed that Cx26 was not localized in the marginal cell layer but was expressed at the border region between the stria vascularis and spiral ligament, and that most Cx26-immunolabeled spots were closely associated with F-actin (Figure 4A). In contrast, a dramatic increase in the expression of ZO-1 occurred in the basal cell layer. ZO-1-positive spots were localized on the cell membrane between overlapping basal cells (Figures 3E and 3F).
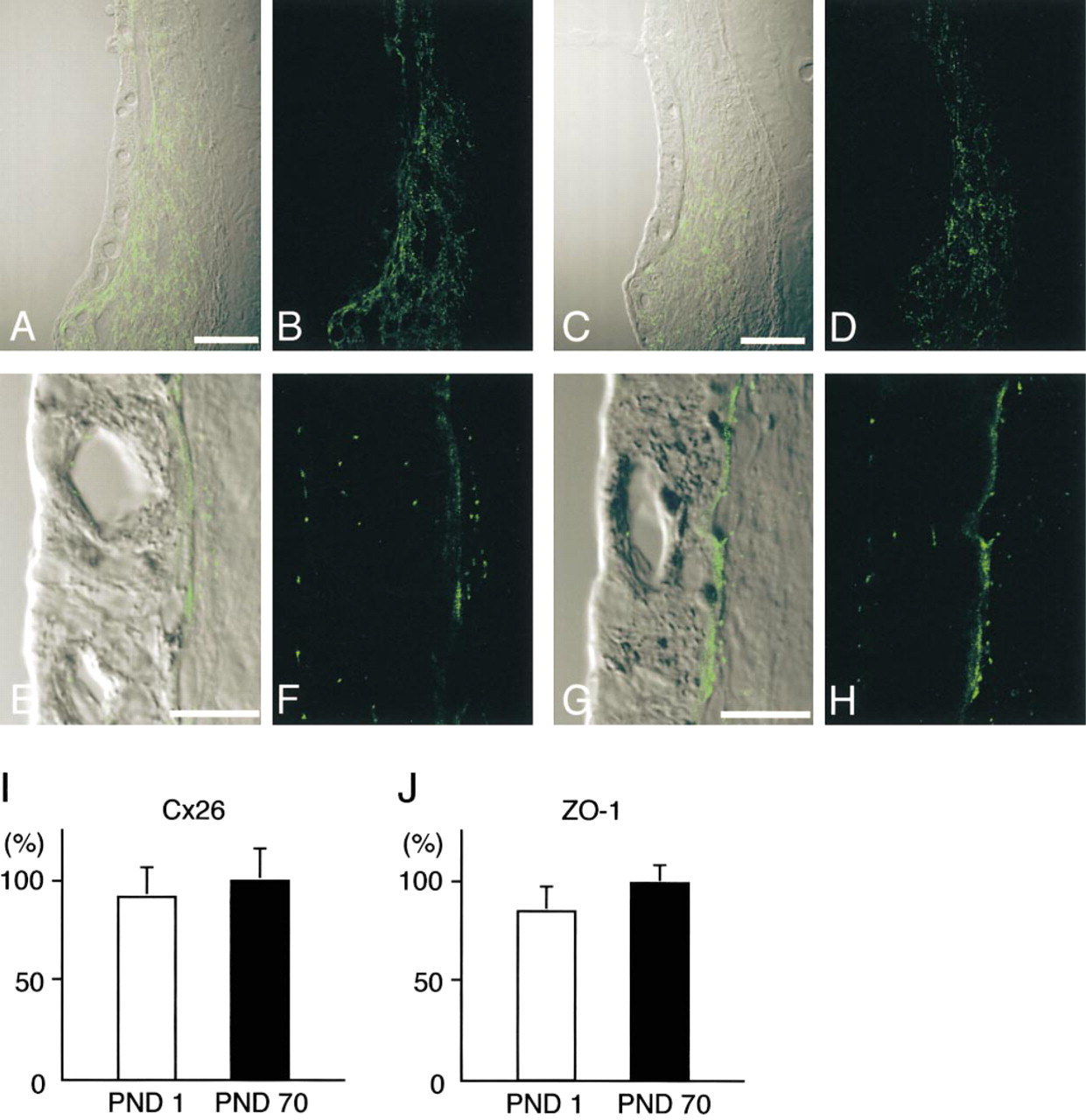
Immunohistochemical localization of Cx26 and ZO-1 in neonatal and adult guinea pig cochleas. Confocal images of Cx26 with DIC imaging (
On PND 11, the basal cells became flatter and aligned more regularly than on PND 7. Afterward, the general morphology of the lateral wall did not change so much. The localization of Cx26-immunolabeled spots extended a little way into the spiral ligament (Figures 2G and 2H). Double-labeling for Cx26 and F-actin demonstrated that Cx26-immunolabeled spots were present not only at the upper and lower surfaces of the basal cells but also on the strial side of the spiral ligament (Figure 4B). ZO-1-positive spots were localized along the basal cell layer (Figures 3G and 3H).
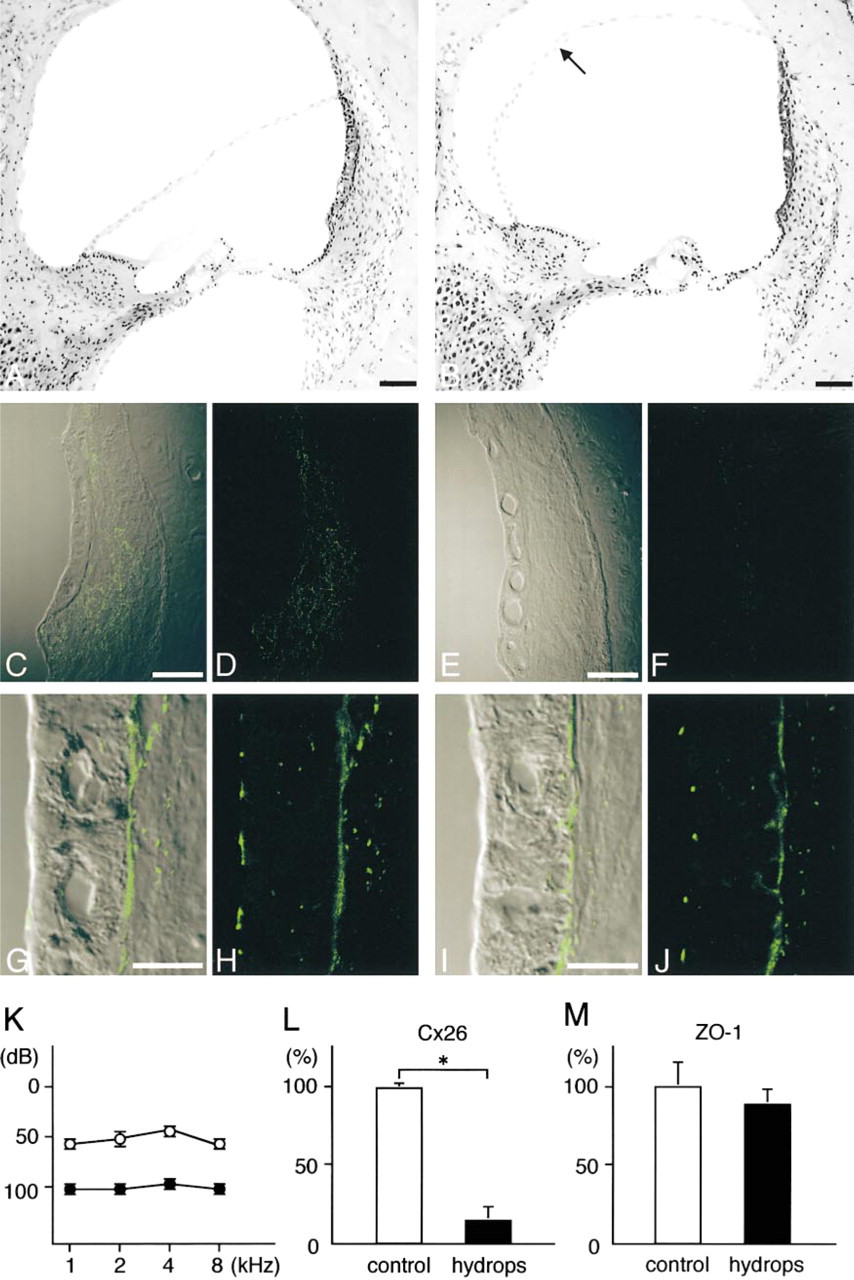
Immunohistochemical localization of Cx26 (
On PND 14, the distribution of Cx26-immunolabeled spots spread dramatically throughout the entire spiral ligament (Figures 2I and 2J). Double labeling for Cx26 and F-actin showed that Cx26-immunolabeled spots were widely distributed in the spiral ligament, including the border with the basal cells in the stria vascularis (Figure 4C). ZO-1-positive spots in the basal cell layer were aligned in a continuous pattern (Figures 3I and 3J).
On PND 21, the density of Cx26-immunolabeled spots moderately increased compared with that of PND 14. The expression pattern of Cx26 (Figures 2K and 2L) appeared similar to that in the adult rat cochlea (PND 70). ZO-1-positive spots formed a linear structure that surrounded basal cells (Figures 3K and 3L).
Morphometric analysis for Cx26 expression during postnatal development of the rat demonstrated that the Cx26-positive area per area of spiral ligament followed a sigmoid time course, i.e., there was a gradual increase from PND 1 to PND 11, a steep rise from PND 11 to PND 14, and a moderate increase from PND 14 to adulthood (Figure 5A). Morphometric analysis demonstrated that ZO-1 expression markedly increased on PND 7 and afterward the increase gradually slowed (Figure 5B).
Cx26 and ZO-1 in the Cochlea of Neonatal and Adult Guinea Pigs
To confirm the relationship between maturation of endocochlear potential, and development of gap junctions and tight junctions, neonatal and adult guinea pigs, in which species endocochlear potential is mature at birth, were analyzed by Cx26 and ZO-1 immunolabeling. On PND 1, Cx26 was already expressed widely in the spiral ligament of the guinea pig (Figures 6A and 6B). There was no apparent difference in the Cx26 expression in the guinea pig cochlea between PND 1 and PND 70 (Figures 6C and 6D). The guinea pigs showed abundant expression of ZO-1 both at the apical surfaces of the marginal cells and in the basal cell layer from PND 1 (Figures 6E and 6F). Morphometric data confirmed that on PND 1, the Cx26-positive area per area of spiral ligament reached the adult level in guinea pigs (no significant difference between PND 1 and PND 70) (Figure 6I), which coincided with the endocochlear potential of neonatal and adult guinea pigs reported by Raphael et al. (1983). Morphometric analysis for ZO-1 expression in the basal cell layer of the cochlea showed no significant difference between neonatal and adult guinea pigs of PND 70 (Figure 6J).
Changes in Cx26 and ZO-1 in the Cochlea of the Guinea Pig with Experimental Endolymphatic Hydrops
Figure 7K shows the brainstem response thresholds for the operated and unoperated ears at 3 months after right endolymphatic duct and sac obliteration. The operated ears had higher thresholds of responses to the stimulus at each frequency, i.e., 1, 2, 4, and 8 kHz, than the unoperated ears in the same animals. Light microscopic examination demonstrated distention of Reissner's membrane in all turns of the operated ears (Figure 7B), whereas the unoperated control ears on the left side had no hydrops (Figure 7A). No severe cell injury was noted in the operated ears, including the stria vascularis and spiral ligament, although mild atrophy of the stria vascularis at all turns and a decrease in the number of spiral ganglion cells at the apical turns (data not shown) were observed.
In the hydropic ears, the number of Cx26-immunolabeled spots decreased in the spiral ligament and in the basal cell layer of the stria vascularis in each turn (Figures 7E and 7F). Morphometric analysis for Cx26 expression in the cochlea (Figure 7L) demonstrated a significant difference between control and hydropic ears (p > 0.05).
In contrast to Cx26, ZO-1 immunolabeling showed no drastic changes in the lateral wall of the hydropic ears, i.e., the expression of ZO-1 did not decrease or increase either on the apical surface of the marginal cells or in the strial basal cell layer (Figures 7I and 7J). Morphometric analysis for the expression of ZO-1 in the basal cell layer of the cochlea (Figure 7M) showed no significant difference between the control and hydropic ears.
Discussion
This study shows that gap junctions and tight junctions in the cochlear lateral wall increase in different spatiotemporal manners during postnatal development of rat auditory function. We found that Cx26 expression increased mainly in fibrocytes in the spiral ligament, whereas ZO-1 expression increased in the basal cell layer of the stria vascularis but did not change at the apical surfaces of the marginal cells of the stria vascularis. Our morphometric analysis demonstrated that Cx26 expression was rapidly augmented on PND 14, whereas ZO-1 expression was augmented on PND 7.
Our results also indicated that gap junctions and tight junctions in the cochlear lateral wall were differentially regulated during experimental endolymphatic hydrops of the guinea pig. The evidence showed that Cx26 expression significantly decreased in the operated ears compared with the unoperated ears in the same animals, whereas there were no obvious differences in ZO-1 expression between them.
Our present work has confirmed the observations of recent studies on rats (Lautermann et al. 1999) and mice (Xia et al. 1999), which showed changes in the distribution patterns of connexins during the postnatal period. However, our study includes several new observations that provide a more complete understanding of spatiotemporal changes in gap junctions in the cochlear lateral wall during the development of auditory function.
First, we found a sigmoidal increase in the Cx26 expression in the cochlear lateral wall by confocal microscopy and morphometry. It was reported that postnatal development of endocochlear potential also shows a sigmoidal time course, as indicated in Figure 5C (quoted from Bosher and Warren 1971). The rapid increase in the Cx26 expression from PND 11 to PND 14 coincided with the rapid rise in endocochlear potential. Furthermore, our data showed that even after PND 14, Cx26 expression increased gradually. This result differed from those of previous studies. Lautermann et al. (1999) reported that beginning from PND 12, an adult-like staining pattern was reached for Cx26 and Cx30 in the rat cochlea. Xia et al. (1999) found no obvious change of Cx26 expression in mouse fibrocytes after PND 15. The discrepancies between our present results and those of other studies are likely due to differences in study methods. Our confocal and morphometric analyses made it possible for us to detect subtle changes in Cx26 expression. The gradual increase in Cx26 after PND 14 agreed with that in endocochlear potential during the same period reported for the rat (Bosher and Warren 1971; Rybak et al. 1992).
Second, new observations were made by comparison between rats and guinea pigs, also indicating a close relationship of Cx26 expression with endocochlear potential development. This is because in the guinea pig, in which species endocochlear potential is mature at birth (Raphael et al. 1983), Cx26 was already expressed widely in the spiral ligament on PND 1 and there was no apparent difference in the Cx26 expression between PND 1 and PND 70.
As far as we know, this is the first report on ZO-1 in the cochlea. ZO-1 has been reported to localize not only in tight junctions but also in adherens junctions, depending on the cell type (Itoh et al. 1993). To confirm that ZO-1 is specifically localized in tight junctions in the cochlea, serially sliced sections were immunolabeled for ZO-1 and occludin, a tight junctionspecific integral protein. Because the localization of ZO-1 in the cochlear lateral wall was similar to that of occludin, it is reasonable to consider that immunolabeling for ZO-1 reflects the presence of tight junctions.
Our developmental study using rats and guinea pigs showed that ZO-1 expression in the basal cell layer of the stria vascularis was closely related to hearing development, suggesting involvement of tight junctions in auditory function. The evidence demonstrated that in rats, in which species endocochlear potential develops dramatically during the postnatal period, especially from days 11 to 14, ZO-1 expression markedly increased on PND 7. After this, the increase gradually slowed. In contrast, in the guinea pig, in which species endocochlear potential is mature at birth, ZO-1 was already expressed strongly in basal cells on PND 1 and there was no apparent difference in ZO-1 expression between PND 1 and PND 70. Our morphometric data on ZO-1 in rats are consistent with those on the increase in tight junction strands on lateral membranes of basal cells in gerbils obtained by freeze-fracture analysis (Souter and Forge 1998). Considering the two-cell model for the generation of the endocochlear potential, the early development of tight junctions at the apical surfaces of the marginal cells and the following development in the basal cells are essential events for the maturation of inner ear function.
The different spatiotemporal development of gap junctions and tight junctions in the rat cochlear lateral wall during postnatal development of auditory function suggests that maturation of these two intercellular junctions is independently regulated. However, we cannot exclude the possibility that during development these two junctions interact with each other, especially in the basal cells of the stria vascularis. This is because both junctions exist next to each other in some parts of basal cells and because a close relationship between connexins and tight junction-related proteins, including ZO-1 (Giepmans and Moolenaar 1998; Toyofuku et al. 1998) and occludin (Nagaoka, et al. 1999), has been reported.
Different regulation of gap junctions and tight junctions in the cochlear lateral wall has also been demonstrated by the present data on experimental endolymphatic hydrops in the guinea pig. Cx26 expression significantly decreased in the operated ears, whereas there were no obvious changes in ZO-1 expression between the operated and control ears. These results indicate that hearing loss in this model is associated with dysfunction of gap junctions rather than of tight junctions. One possible explanation for the difference may be that fibrocytes in the spiral ligament are more seriously affected than other cells by experimental endolymphatic hydrops, and consequently the expression of Cx26 in these fibrocytes is reduced. This explanation is supported by the results reported by Ichimiya et al. (1994) and by Nadol et al. (1995), who demonstrated selective decreases of immunoreactivity of these fibrocytes, not only for various enzymes such as Ca2+-ATPase and aldehyde dehydrogenase but also for Cx26 3 months after endolymphatic duct obliteration. Our data on ZO-1 in hydrops are consistent with those obtained by the freeze-fracture studies of Jahnke et al. (1985).
In conclusion, using confocal microscopy and morphometry, we have shown that gap junctions and tight junctions in the cochlear lateral wall increase in different spatiotemporal manners during postnatal development of rat auditory function, and that gap junctions and tight junctions in the cochlear lateral wall are differentially regulated during hearing loss induced by experimental endolymphatic hydrops in the guinea pig. Because mutations in multiple connexin genes other than Cx26 have been found in human hereditary deafness (Xia et al. 1998; Grifa et al. 1999) and at least 18 members of the claudin (tight junction-specific integral protein) family have been identified (Tsukita and Furuse 2000), a significant number of molecules associated with gap junctions and tight junctions surely contribute to the auditory system under physiological and pathological conditions.
Footnotes
Acknowledgements
Acknowledgments
We gratefully acknowledge the generous gift of the polyclonal anti-connexin26 antibody from Dr Y. Shibata (Kyushu University School of Medicine; Fukuoka, Japan). We thank Mr Kim Barrymore for advice on English usage.