
Abstract
Gap junctions (GJs) have been shown to play a role in tumor progression including a variety of keratinocyte-derived and non-keratinocyte-derived skin tumors. Here we show that the synthesis of the GJ proteins connexin 26 and connexin 30 (Cx26 and Cx30) is induced in keratinocyte-derived epithelial skin tumors whereas there is either no change or a downregulation of Cx43. Cx26, Cx30, and Cx43 are absent in non-epithelial skin tumors. Further, Cx26 and Cx30 are induced in the epidermis adjacent to malignant melanoma but absent in the epidermis adjacent to benign non-epithelial skin lesions (melanocytic nevi and angioma). The keratinocyte-derived skin tumors are very heterogeneous regarding the Cx26/Cx30 pattern in the epidermis at the periphery of the tumors. We did not observe any difference in the localization of the very similar proteins Cx26 and Cx30 but a variation in intensity of immunoreactivity. As the staining patterns of Cx26 and Cx30 antibodies are not identical to those of CK6, a marker for hyperproliferation, and CK17, a marker for trauma, we discuss that the induction of these gap junctional proteins exceeds a reflection of reactive hyperproliferative or traumatized epidermis. We further discuss the putative roles of these gap junctional proteins in tumor progression.
Keywords
C
In the skin, GJIC has been hypothesized to be involved in the regulation of keratinocyte growth, differentiation, and migration (Pitts et al. 1988; Salomon et al. 1993; Brissette et al. 1994; Goliger and Paul 1995), as well as in keratinocyte-melanocyte interaction (Hsu et al. 2000; Li et al. 2002). The importance of epidermal GJIC has been demonstrated by the identification of mutations in genes coding for Cx in autosomal epidermal diseases (Richard et al. 1998; Maestrini et al. 1999; Wilgoss et al. 1999; Kelsell et al. 2000; van Steensel et al. 2002).
In multicellular organisms, homeostasis is governed by either endocrine or paracrine communication via soluble factors including hormones, growth factors, and cytokines or by intercellular communication via cell-cell and cell-matrix adhesion and GJIC (reviewed in Haass and Herlyn 2005; Haass et al. 2004, 2005). GJs have been shown to play a role in progression of a variety of tumors (reviewed in Yamasaki et al. 1999; Carystinos et al. 2001; Naus 2002; Haass et al. 2004). Available data for human skin tumors are limited: Wilgenbus et al. (1992) investigated Cx26 and Cx43 in basal cell carcinoma (BCC) but did not distinguish between the highly homologous Cx26 and Cx30. Tada and Hashimoto (1997) showed that Cx43 is present in the cytoplasm and in poorly developed GJ in BCC and squamous cell carcinoma (SCC). Transfection of Cx43 into a Cx43-deficient melanoma cell line suppressed anchorage-independent growth in colony-forming efficiency assays, suggesting that Cx43 plays an important role as a tumor suppressor in malignant melanoma (Su et al. 2000). Ito et al. (2000) did not find Cx26 in human melanomas within the epidermis but found an upregulation of Cx26 in the areas of melanomas invading into the dermis; they did not distinguish between Cx26 and Cx30. Hsu et al. (2000) showed that melanocytes, but not melanoma cells, were able to communicate with keratinocytes via GJIC; instead, melanoma cells communicate among themselves and with fibroblasts.
Here we investigated various forms of BCC, SCC, Bowen's disease, and malignant melanoma using specific antibodies for Cx26, Cx30, and Cx43, the latter known to be expressed ubiquitously in normal epidermis. BCC is a locally destructive tumor that does not metastasize and is generally considered “semi-malignant.” SCC of the skin is a malignant tumor that metastasizes in ∼5% of the cases. Bowen's disease is an intraepidermal carcinoma in situ that turns into an SCC/Bowen's carcinoma after invasion through the basement membrane. Melanoma is the most aggressive malignant skin tumor. In contrast to the above-listed tumors, melanoma is of mesenchymal origin. Previously we investigated Merkel cell carcinoma (MCC), a highly malignant neuroendocrine cutaneous carcinoma, using antibodies to Cx26, Cx30, and Cx43 (Haass et al. 2003). For comparison we investigated benign hyperkeratinocytic lesions (seborrheic keratoses, actinic keratoses, and common warts) and benign melanocytic lesions (melanocytic nevi). To show that changes of the Cx expression pattern in the epidermis adjacent to the above-mentioned lesions are not related to the space these lesions occupy within the skin, we also investigated the epidermis adjacent to angiomas. Moreover, we used cytokeratins 6 and 17 (CK6 and CK17) as markers for epidermal hyperproliferation and trauma, respectively. We show the different characteristics of “keratinocytic” vs “non-keratinocytic” hyper/neoplasias regarding the expression of Cx. In the tumor-surrounding epidermis, we demonstrate striking differences among tumors, which seem to correlate to their malignancy.
Materials and Methods
Tissues, Antibodies, and Nuclear Dyes
Samples of human BCC (solid: n=6; superficial: n=5; infiltrative: n=4), SCC (n=14), Bowen's disease (n=6), actinic keratoses (n=5), seborrheic keratoses (n=5), common warts (n=5), malignant melanoma (n=8), melanocytic nevi (n=8), and angiomas (n=5) were obtained from our Department of Dermatology and Venerology, Hamburg, Germany (for MCC see Haass et al. 2003). In agreement with the local medical ethics committee (OB-008/04), samples were used after diagnostic procedures had been completed. All patients gave informed consent. Normal human skin was obtained during the routine removal of epidermal cysts and tumors; the samples were localized at least 2 cm from any lesion. Murine liver was taken from euthanized C57/Bl6 mice. Immediately after excision, all tissue samples were transferred to 3.5% formalin and embedded in paraffin.
Antibodies specific for Cx30 (#71–2200), and Cx43 (#71–0700) were purchased from Zymed Laboratories (San Francisco, CA), for CK6 (Ks6.KA12) and CK17 (Ks17.E3) from Progen Biotechnik GmbH (Heidelberg, Germany), and for Melan A (A103) from DAKO (Glostrup, Denmark). Antibodies specific for Cx26 (gpCx26) were designed in our laboratory and produced by Peptide Specialty Laboratories GmbH (Heidelberg, Germany) by immunizing guinea pigs with KLH-coupled peptides highly specific for Cx26 (residues 102–114 of the Cx26 peptide: KKRKFIKGEIKSE) (Meyer et al. 2002; Brandner et al. 2004). These antibodies were affinity purified with the corresponding peptide as previously described (Haass et al. 1996). The specificity of gpCx26, especially with regard to cross-reactions with Cx30 antigen, has been shown previously (Meyer et al. 2002; Brandner et al. 2004). We also used other polyclonal (#71–0500; Zymed Laboratories) and monoclonal (Clone CX-12H10, #13–8100; Zymed Laboratories) antibodies against Cx26 for some stainings. Both antibodies confirmed our results but proved to be much weaker than gpCx26. Moreover, they showed unspecific cytoplasmic staining, and both are known to cross-react with Cx30 and, in the case of the polyclonal antibody, with an unknown 45- to 50-kDa protein (see data sheets to CX-12H10 and #71–0500, Zymed Laboratories). Therefore, in this report we show results using gpCx26.
Nuclear staining was performed with DAPI (4′,6-Diamidino-2′-phenylindole-dihydrochloride; Boehringer Mannheim, Mannheim, Germany).
Immunofluorescence Microscopy
Paraffin sections of 5-μm-thick formaldehyde-fixed tissues were deparaffinated and rehydrated. Antigen retrieval was performed by microwave oven heating (three times for 5 min, 600 W) in 10 mM sodium citrate buffer (pH 6.0) and followed by mild trypsinization with 0.001% trypsin for 10 min at 37C. For the Cx26 and Cx30 antibodies, this was followed by blocking the tissues for unspecific binding sites with 0.1% Triton X-100 and 2% dry milk powder in PBS and for the CK6, CK17, and Melan A antibodies with 0.1% Triton X-100, 2% dry milk powder, and 2% normal goat serum in PBS. Primary antibodies were applied for 1 hr at room temperature (gpCx26: 1:500; Cx30: 1:50; Cx43: 1:100; CK6: 1:15; CK17: 1:30; Melan A: 1:30 in PBS), followed by washing the samples three times for 10 min in PBS. Afterwards, Cy2 (1:50 in PBS) and/or Cy3-coupled (1:500 in PBS) secondary antibodies (Dianova; Hamburg, Germany) were applied for 30 min at room temperature, followed by another washing step as above. For negative controls we applied the Cy2- or Cy3-coupled secondary antibodies only. Finally, the slides were washed with ddH2O and coverslips mounted with Fluoromount (Southern Biotechnology Associates, Inc.; Birmingham, AL). An Axiophot II microscope (Carl Zeiss; Jena/Oberkochen, Germany), a CCD Camera (Hamamatsu Photonics; Hamamatsu City, Japan), and Openlab 2.0.4 software (Improvision; Coventry, UK) were utilized to visualize and evaluate the stained sections.
Results
Localization of Cx26, Cx30, and Cx43 in Human Interfollicular Epidermis
In human interfollicular epidermis, Cx30 (Figure 1) and Cx26 (Figure 1) were usually not detectable. Only in some cases there was a weak staining for both proteins in the stratum granulosum (for Cx26 see Figure 1). Cx43 was present in all layers with a clearly weaker staining in the stratum basale (Figure 1D). Previously, the expression of Cx26 in human liver has been shown (Traub et al. 1989), which we used as positive control for our antibody (Figure 1E). The antibody has previously been characterized (Brandner et al. 2004).
Localization of Cx26 and Cx30 in Skin Tumors
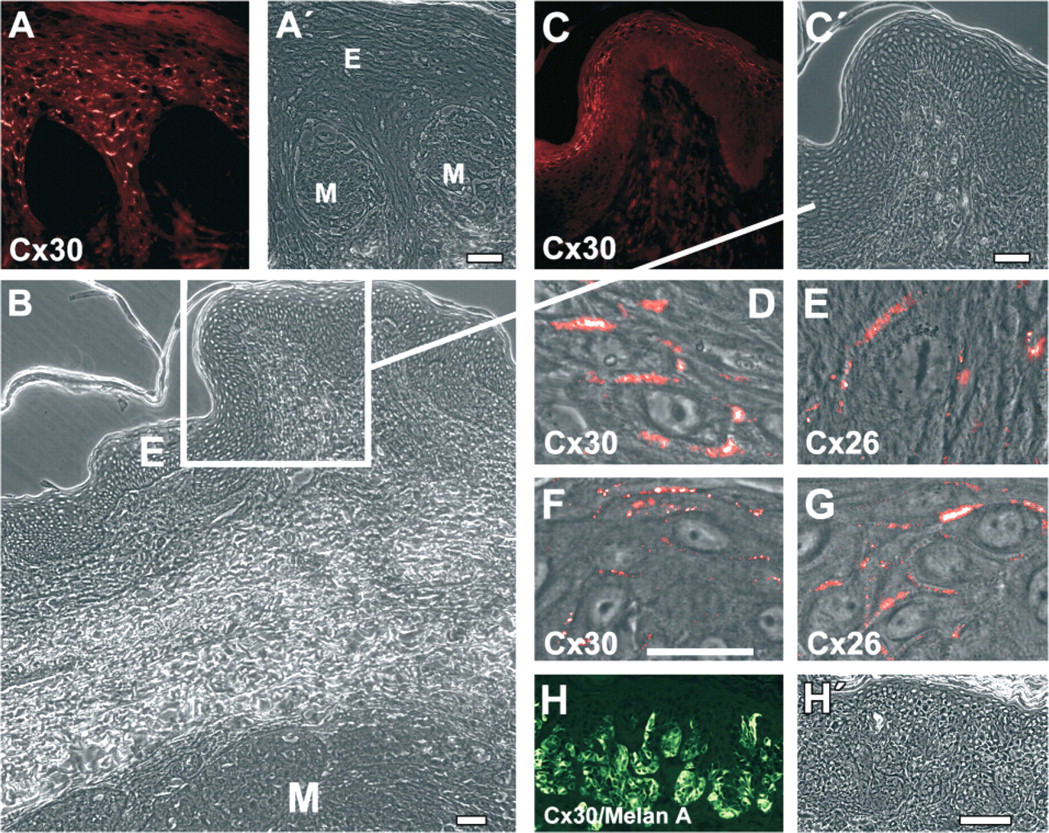
In contrast to human interfollicular epidermis, Cx26 and Cx30 were present in keratinocyte-derived tumor cells, i.e., of nodular, superficial, and infiltrative BCC (Figures 22-A2), SCC (Figures 22-I2), Bowen's disease (Figures 2M and 2N), seborrheic keratoses (Figures 3A and 3B), common warts (Figures 3C and 3D), and actinic keratoses (Figures 3E and 3F). BCC showed a heterogeneous staining for Cx26 and Cx30: there were tumor areas with intense staining (Figures 2A, 2B, and 2D in part) and others with absent or rare staining (Figures 2C and 2D in part). In the latter, the staining was often restricted to basal cells of the BCC (Figures 2C and 2D). Cx26 and Cx30 were localized to the cell-cell borders in the typical punctuate pattern (Figures 22-E2). In SCC, differentiated areas showed strong reactivity to the Cx26 and Cx30 antibodies, whereas dedifferentiated, i.e., more invasive, areas showed weak reactivity (Figures 2I and 2J). Also in SCC, Cx26 and Cx30 showed a typical GJ staining at the cell-cell borders (Figures 2K and 2L). The reactivity in Bowen's disease was weak (Figures 2M and 2N) and in Bowen's carcinoma barely detectable (data not shown). Also, in seborrheic keratoses, common warts, and actinic keratoses there were areas positive for Cx26—again at the cell-cell borders—and others that were negative (Figure 3). Cx26 considerably colocalized with Cx30 in the keratinocyte-derived tumors, but there were some structures heterogeneously distributed within the tumor reacting more intensively with the Cx26 antibodies and others with the Cx30 antibodies, respectively (Figure 4). Neither Cx26 nor Cx30 was detected in primary malignant melanoma (Figure 4B and Figure 5), melanoma metastasis (data not shown), and melanocytic nevi (Figure 5H). They also were not detectable in angioma (data not shown).
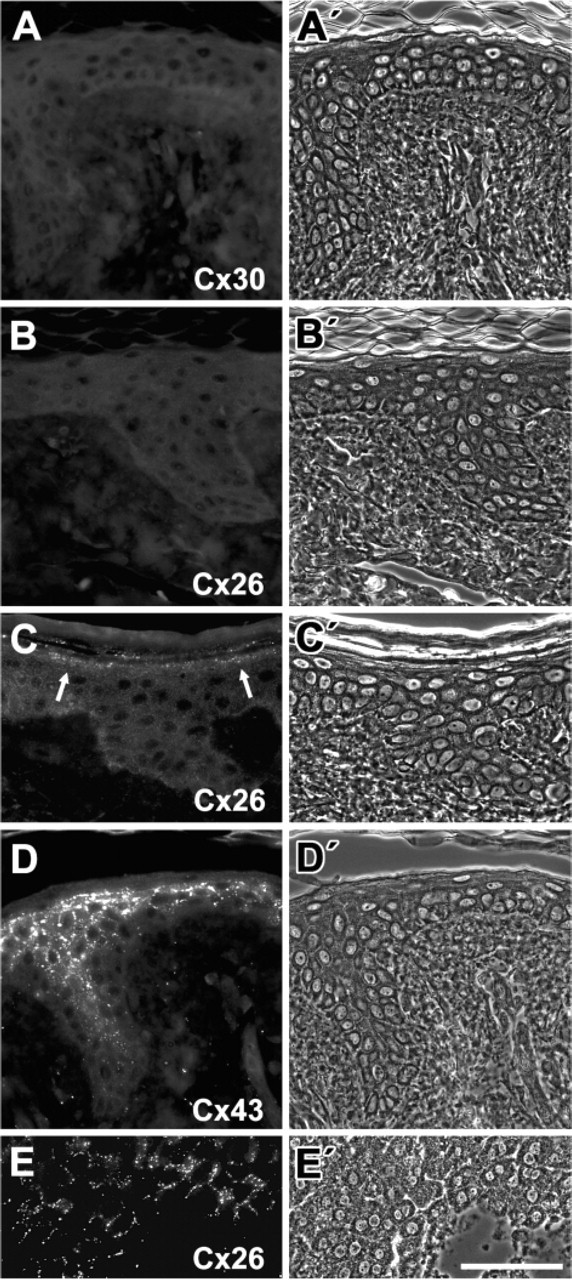
Connexins (Cx) in normal interfollicular epidermis. Immunolocalization of Cx30 (
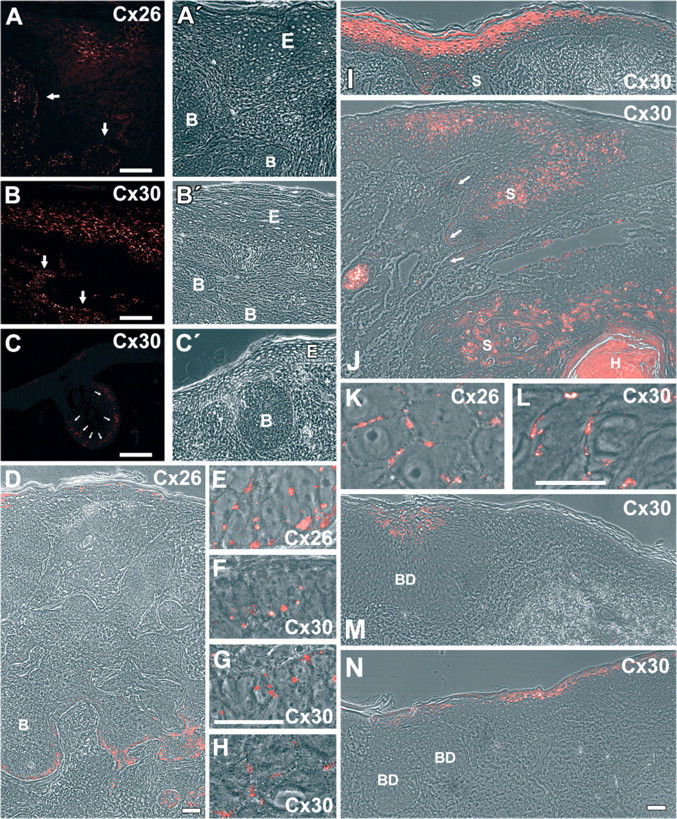
Localization of Cx26 and Cx30 in basal cell carcinoma (BCC), squamous cell carcinoma (SCC), Bowen's disease, and adjacent epidermis. Immunofluorescence microscopy of Cx26 (
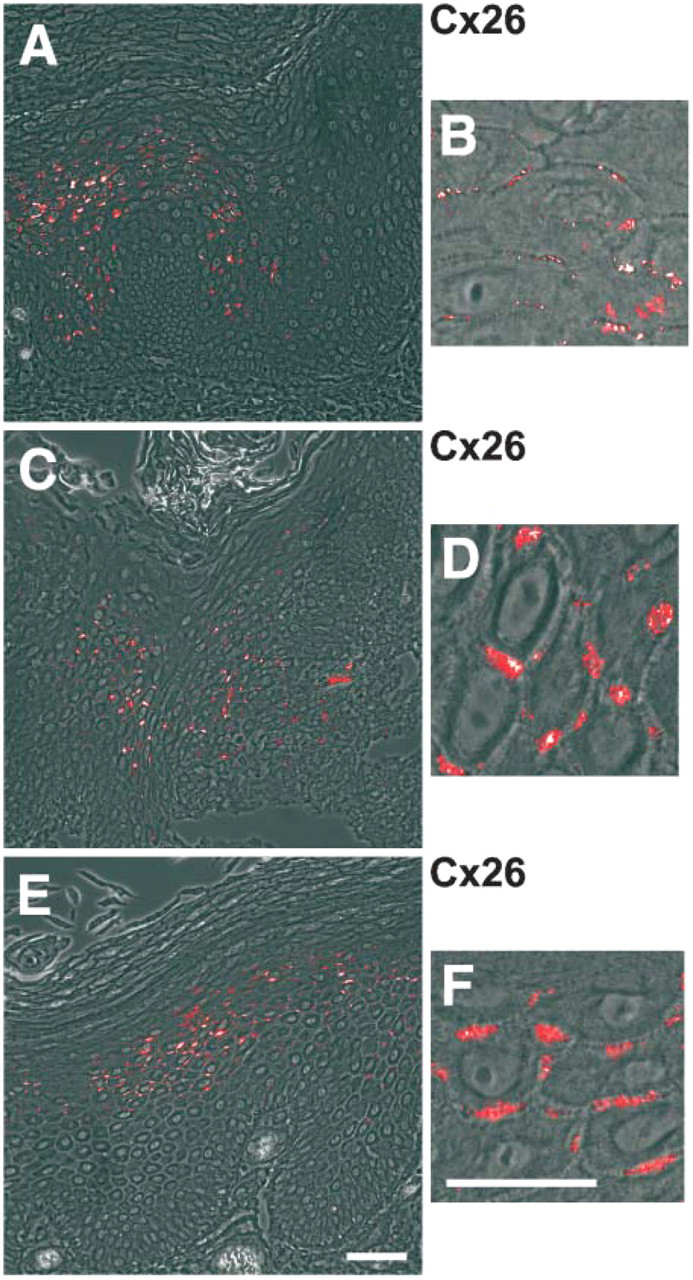
Immunolocalization of Cx26 in seborrheic keratoses (
Localization of Cx26 and Cx30 in Epidermis Adjacent to Skin Tumors
In this report, we defined the epidermis above or around the investigated tumors as “epidermis adjacent to the tumor.” Interestingly, in the epidermis adjacent to BCC, keratinocytes strongly positive for Cx26 (Figures 2A and 2D) and Cx30 (Figure 2) were typically restricted to the contact zone of epidermis and tumor and again showed the typical punctuate GJ pattern at the cell borders (for Cx30, see Figure 2H). Where there was no infiltration of the epidermis, Cx26 and Cx30 usually were absent (Figure 2). However, occasionally a weak reactivity in the stratum granulosum was observed—similar to the weak reactivity in normal epidermis (cf. Figure 1). All investigated types of BCC (nodular, superficial, and infiltrative) showed a similar pattern.
The analysis of SCC and Bowen's disease sections was aggravated by the lack of a clear demarcation of tumor and adjacent epidermis. The highly differentiated regions at the margins of SCC, which strongly resemble “normal epidermis,” usually were positive for Cx26 and Cx30 (Figures 2I and 2J). However, without established SCC markers it remains unclear whether these areas were “activated” epidermis or part of the tumor (for investigation of CK6 and CK17 in SCC, see below). The epidermis in the area of the Bowen's disease was either negative for both antibodies (Figure 2M) or positive in areas with some Bowen cells (Figure 2N).
Surprisingly, Cx26 and Cx30 were present in the keratinocytes of the suprabasal layers of the interfollicular epidermis adjacent to primary malignant melanoma (Figures 4B-4D, Figures 5A, 5D, and 5E) and to melanoma metastasis (Figures 5B, 5C, 5F, and 5G) but not to melanocytic nevi (Figure 5H) and angioma (data not shown). At the light microscopic level, Cx26 significantly colocalized with Cx30 in the epidermis adjacent to malignant melanoma (Figures 44-B4). However, the intensity of immunoreactivity was variable among the different layers of the epidermis: there was a stronger reactivity for Cx26 in the upper suprabasal layers compared with the lower layers (Figures 4B, 4B″, 4C, 4C″, 4D, and 4D″) in contrast to the stronger reactivity of Cx30 in the lower layers compared with the upper suprabasal layers (Figures 4B, 4B″, 4C, 4C″, 4D, and 4D″).
Localization of Cx43 in Skin Tumors and Epidermis Adjacent to Skin Tumors
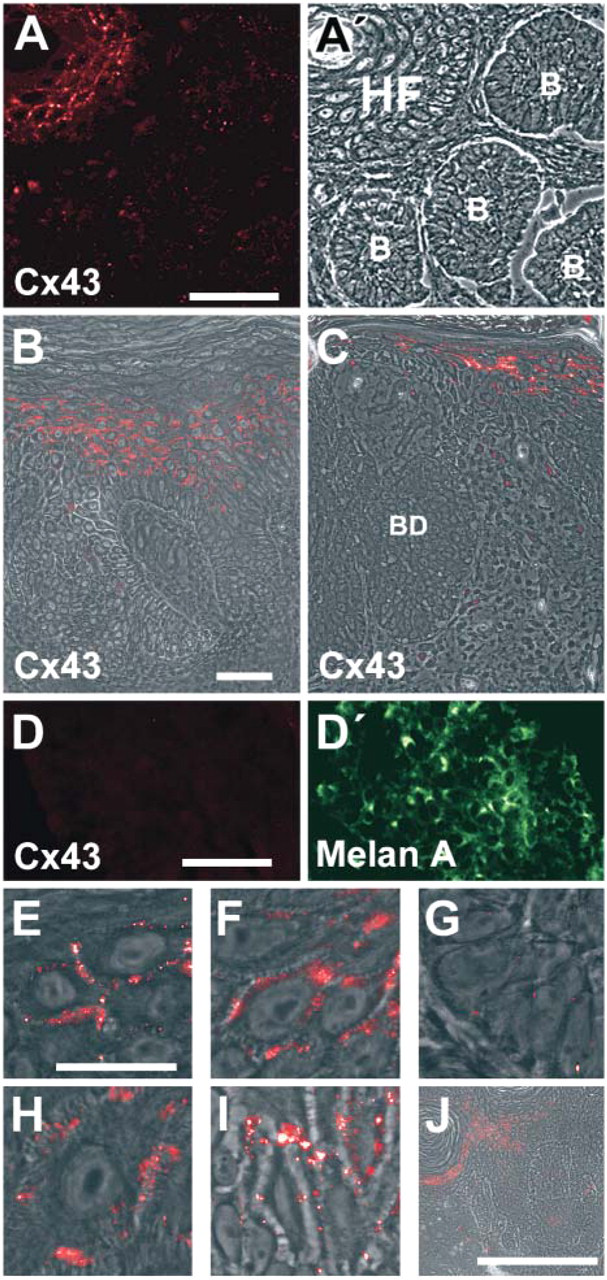
Cx43 was present in BCC. Its staining intensity was similar to the stratum basale in normal epidermis but lower than in suprabasal layers of normal epidermis and in hair follicles (Figure 6). This was mainly due to a low expression of Cx43 within the BCC cells leading to fewer dots in the punctuate GJ pattern at the cell borders (Figure 6G). In addition, in the center of BCC some cells appeared negative for Cx43 (Figure 6). Cx43 was also present in SCC, but not in all areas (Figure 6). It was barely detectable in Bowen's disease (Figure 6) and not detectable in malignant melanoma (Figure 6D). In suprabasal epidermis (Figure 6E), hair follicles (Figure 6F), BCC (Figure 6G), SCC (Figure 6H), and Bowen's disease (Figure 6I) Cx43 showed a typical GJ staining at the cell borders. Cx43 was found in common warts and in seborrheic keratoses (Figure 6J; cf. Table 1). It was expressed throughout the interfollicular epidermis, adjacent to and distant from all investigated tumors (data not shown).
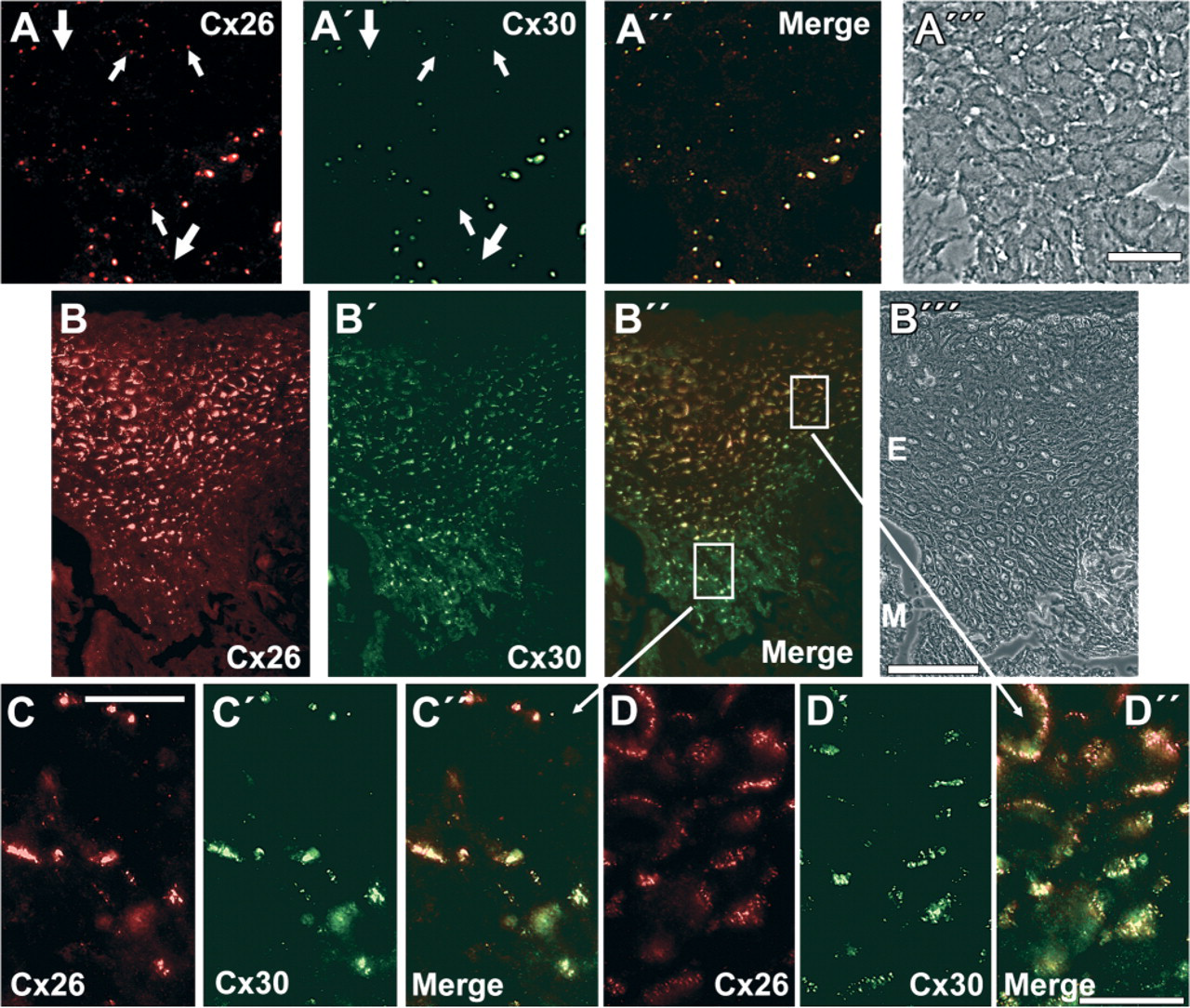
Colocalization of Cx26 and Cx30. Immunolocalization of Cx26 (red;
Relation between Cx and Markers for Hyperproliferation and Trauma
The staining pattern of the hyperproliferation marker cytokeratin 6 (CK6) (Navarro et al. 1995; Porter et al. 1998) was not identical to those of Cx26/Cx30 (see Table 1). In contrast to Cx26 and Cx30, CK6 was negative or detectable only in small cell clusters within BCC (Figures 7A and 7B). Its distribution in the adjacent epidermis was in some cases more restricted (Figure 7) and in others more widespread than Cx26 and Cx30 (data not shown). Moreover, CK6 was absent or barely detectable in epidermis adjacent to MCC (data not shown), whereas Cx26/Cx30 were clearly present (Haass et al. 2003). CK6 and Cx26/30 were present in SCC and adjacent epidermis, but the distribution of CK6 at the margins of the tumor was more widespread (Figure 7). The distribution patterns of Cx26/Cx30 and CK6 were similar in malignant melanoma, i.e., they were absent in malignant melanoma but present in its adjacent epidermis (Figure 7D). However, the localization patterns of CK6 and Cx26/Cx30 were not identical, even though there was a large overlap (Figure 7D). An expression pattern different from Cx26 and Cx30 was also true for cytokeratin 17 (CK17, see Table 1). CK17, which is known to be positive in hair follicles (Moll et al. 1982), is often associated with traumatized epidermis. CK17 was strongly positive in BCC but barely detectable in epidermis adjacent to BCC (Figure 7E), SCC, malignant melanoma, and MCC (data not shown).
Localization of Cx26 and Cx30 in malignant melanoma, melanocytic nevus, and adjacent epidermis. Immunofluorescence microscopy of Cx30 (
Discussion
In the present study we demonstrate an induction of Cx26 and Cx30 in various forms of BCC and in SCC, whereas there is either no change or a downregulation of Cx43. This confirms previous results for Cx26 and Cx43 in BCC (Wilgenbus et al. 1992) and for Cx43 in BCC and SCC (Tada and Hashimoto 1997). We additionally show that benign lesions, such as seborrheic keratoses and common warts, synthesize Cx26, Cx30, and Cx43 as well (for induction of Cx26 in common warts, see also Lucke et al. 1999). The induction of Cx26 and Cx30 is similar to various conditions characterized by hyperproliferation, e.g., psoriasis, wound healing, normal epidermis treated with retinoic acid, and epidermal equivalents (Goliger and Paul 1995; Masgrau-Peya et al. 1997; Rivas et al. 1997; Labarthe et al. 1998; Lucke et al. 1999; Wiszniewski et al. 2000; Coutinho et al. 2003; Kretz et al. 2003; Brandner et al. 2004). However, the staining patterns of Cx26/Cx30 are different from that of the hyperproliferation marker CK6 in some of these tumors (e.g., CK6 is barely detectable in BCC but more widespread in Bowen's disease), suggesting that Cx26 and Cx30 do not only indicate hyperproliferation.
Cx43 in BCC, SCC, Bowen's disease, malignant melanoma, and seborrheic keratosis. Immunofluorescence microscopy of Cx43 (red) in superficial BCC (
Interestingly, we found a heterogeneous distributionaof Cx26 and Cx30 in different areas of epithelial tumors. In BCC they are concentrated in the basal cell layer. Moreover, we observed an increase of Cx26 and Cx30 in tumor areas deep in the dermis compared with those close to the epidermis, suggesting an increase in proliferating invasive areas of the BCC. In SCC, we observed a correlation of staining reactivity and tumor differentiation. Highly differentiated, spinous-like cells are clearly positive, whereas less-differentiated cells are weakly positive for Cx26 and Cx30. Also in Bowen's disease, Cx26 and Cx30 are found mainly in differentiated areas, whereas they are missing in the highly invasive areas. These data suggest that GJIC via GJ containing Cx26 and Cx30 is absent in the invasive areas of SCC and Bowen's disease.
Thus, there is a contrast with regard to Cx26 and Cx30 staining patterns in the invasive areas of the malignant SCC and Bowen's carcinoma, compared with those of the semi-malignant BCC. Whereas the first are negative for Cx26 and Cx30, the latter are positive. The presence of Cx26 and Cx30 in these areas of the semi-malignant BCC is similar to their presence in benign epithelial tumors, whereas the absence of Cx26 and Cx30 in the respective areas of SCC and Bowen's carcinoma is similar to their absence in other aggressive skin tumors, which are also able to metastasize, e.g., malignant melanoma and MCC (Haass et al. 2003). On the other hand, there are both positive and negative areas for Cx26 and Cx30 in seborrheic keratoses, common warts, and actinic keratoses also, which make a central role of the absence of these proteins in metastasis unlikely.
In malignant melanoma there is a lack of Cx43 and no induction of Cx26 and Cx30. This is identical to MCC, another highly aggressive tumor in the skin (Haass et al. 2003), and to invasive areas of SCC and Bowen's disease (see above). Interestingly, there is no difference regarding the presence of Cx between malignant melanoma and melanocytic nevi. In contrast to these in vivo data, synthesis of Cx43 has been shown in cultured melanocytes and melanoma cell lines as well (Hsu et al. 2000). However, cell culture effects may play an important role in these differences. Our data confirm the downregulation of Cx43 in melanoma in gene microarrays (Su et al. 2000). The presence of Cx26 in the cytoplasm of cells in advanced human melanoma (Ito et al. 2000) was seen in our experiments only when we used the antibody used by Ito and colleagues, which also shows a strong cross-reaction with the cytoplasm of outer root sheath cells of hair follicles. Moreover, the antibody against Cx26 used by Ito and colleagues is cross-reactive with Cx30 (see data sheet to CX-12H10, #13–8100; Zymed Laboratories), whereas the antibodies used in this study are highly specific for Cx26 and Cx30, respectively.
Immunoreactivities to Cx26, Cx30, Cx43, CK6, and CK17 antibodies in interfollicular epidermis, skin tumors, and their adjacent epidermis
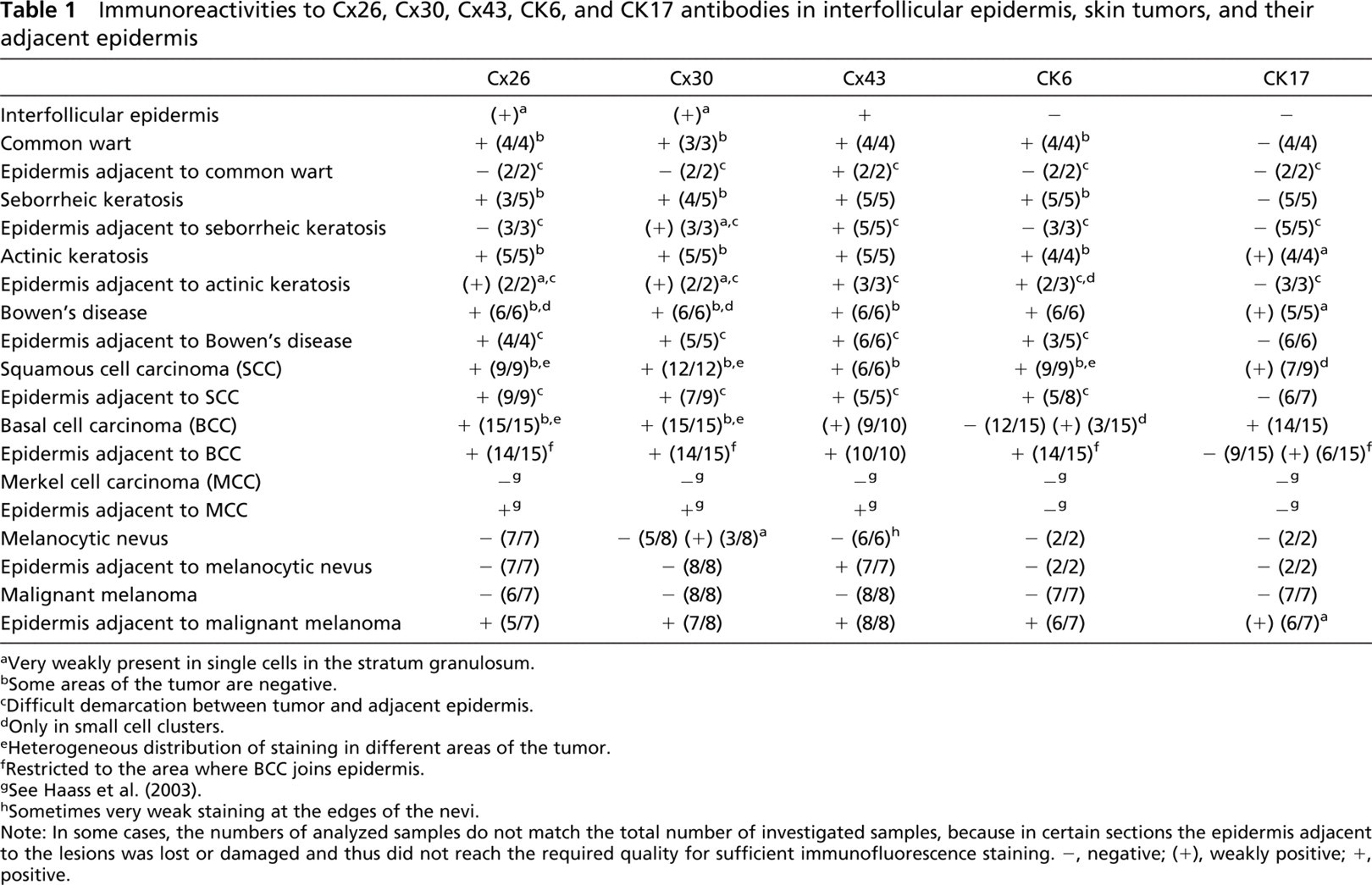
aVery weakly present in single cells in the stratum granulosum.
bSome areas of the tumor are negative.
cDifficult demarcation between tumor and adjacent epidermis.
dOnly in small cell clusters.
eHeterogeneous distribution of staining in different areas of the tumor.
fRestricted to the area where BCC joins epidermis.
gSee Haass et al. (2003).
hSometimes very weak staining at the edges of the nevi.
Note: In some cases, the numbers of analyzed samples do not match the total number of investigated samples, because in certain sections the epidermis adjacent to the lesions was lost or damaged and thus did not reach the required quality for sufficient immunofluorescence staining. -, negative; (+), weakly positive; +, positive.
Probably the most surprising result in the present study is the distinct induction of Cx26 and Cx30 in normal interfollicular epidermis adjacent to the highly aggressive malignant melanoma and MCC (Haass et al. 2003), whereas there is no induction adjacent to the investigated benign lesions (melanocytic nevi and angioma) and a restricted induction adjacent to the semi-malignant BCC. The evaluation of epidermis adjacent to tumors, which often comprises the whole epidermis, e.g., SCC, Bowen's disease, seborrheic keratoses, and common warts, is very difficult as there is no clear demarcation between tumor and epidermis. Without established immunohistochemical markers for SCC and Bowen's disease, it remains unclear whether the restricted Cx26 and Cx30 expression in the epidermal area around SCC and Bowen's disease is “activated” epidermis or part of the tumor. In both cases, Cx26 and Cx30 might be valuable as markers for the completeness of the surgical excision of above-mentioned tumors. Interestingly, the epidermis-like structures adjacent to seborrheic keratoses and common warts are negative or only faintly positive in the granular layer. Summarizing the data derived from epidermis adjacent to non-keratinocyte-derived and keratinocyte-derived skin tumors, the broad and intense induction of Cx26 and Cx30 in the adjacent epidermis may correlate to the invasiveness and thus malignancy of a tumor. These results are independent from inflammatory cells around the tumors: there is no difference between melanomas that were highly inflamed and others in which there was no detectable inflammation. Moreover, there is only a weak induction of Cx26 and Cx30 in inflamed BCC (data not shown). Furthermore, Cx26 and Cx30 do not seem to be only markers for hyperproliferation in the adjacent epidermis, as the expression pattern of CK6, a hyperproliferation-associated marker (Navarro et al. 1995; Porter et al. 1998) is not identical to that of Cx26 and Cx30. CK17, a cytokeratin present in hair follicles (Moll et al. 1982) and frequently accompanying traumatized epidermis, also shows a different expression pattern. Therefore, the expression of Cx26 and Cx30 is not a marker for trauma.
Thus, one might hypothesize that the induction of Cx26/Cx30 in the epidermis adjacent to different malignant skin tumors may play a role in their ability for metastasis or invasion and therefore may be used as a marker for the tumor's malignancy. Moreover, the extent of induction in the epidermis may give a hint for the extent of influence of the tumor on its surrounding tissue and might be useful for the selection of the safety margins for the removal of these tumors. The induction of Cx26/Cx30 within skin tumors is probably not a marker for the malignancy. To clarify the relation between the induction of Cx in tumors and/or tumor-surrounding epidermis and the malignant transformation and to answer the question whether the inhibition of this induction might affect tumor progression will be challenging tasks for the future.
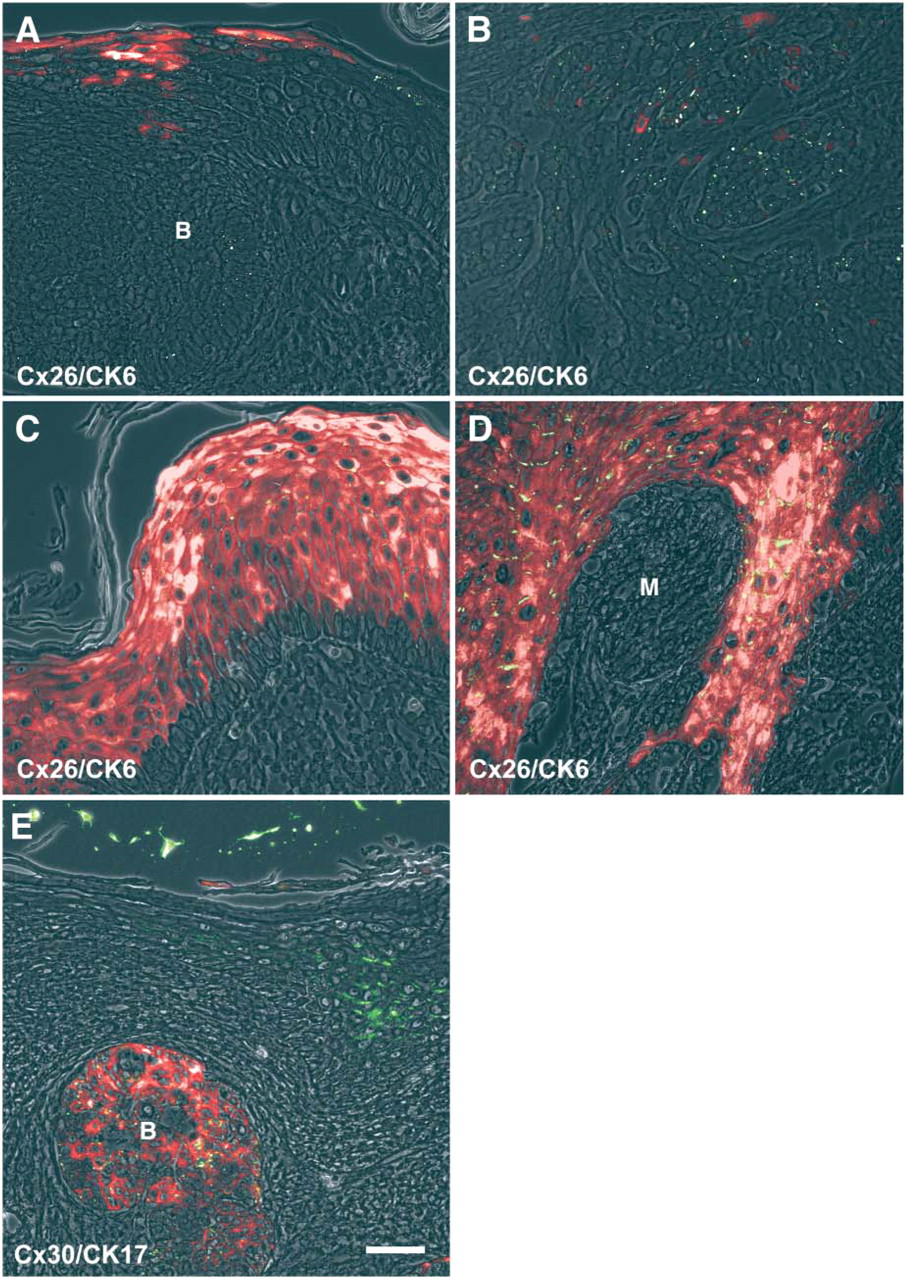
Costaining of Cx26 and Cx30 with markers for hyperproliferation and trauma in various skin tumors. Immunolocalization of CK6 (red) and Cx26 (green) in superficial BCC and its adjacent epidermis (
Footnotes
Acknowledgements
We thank Pia Houdek (Hamburg, Germany) for excellent technical assistance. We also gratefully acknowledge Drs. Roland Moll (Marburg, Germany), Peter von den Driesch (Stuttgart, Germany), Meenhard Herlyn, and Steven Kazianis (Philadelphia, PA) for stimulating discussions.