
Abstract
A monoclonal antibody directed against the amino terminal of rat phosphodiesterase 10A (PDE10A) was used to localize PDE10A in multiple central nervous system (CNS) and peripheral tissues from mouse, rat, dog, cynomolgus macaque, and human. PDE10A immunoreactivity is strongly expressed in the CNS of these species with limited expression in peripheral tissues. Within the brain, strong immunoreactivity is present in both neuronal cell bodies and neuropil of the striatum, in striatonigral and striatopallidal white matter tracks, and in the substantia nigra and globus pallidus. Outside the brain, PDE10A immunoreactivity is less intense, and distribution is limited to few tissues such as the testis, epididymal sperm, and enteric ganglia. These data demonstrate that PDE10A is an evolutionarily conserved phosphodiesterase highly expressed in the brain but with restricted distribution in the periphery in multiple mammalian species.
Keywords
C
PDE10A was identified simultaneously by three laboratories using bioinformatic approaches (Fujishige et al. 1999a,b; Loughney et al. 1999; Soderling et al. 1999). In Northern blots, PDE10A mRNA was highly expressed only in the central nervous system (CNS) and testes. Subsequent studies with an antibody to rat PDE10A localized PDE10A to the medium spiny projection neurons of the rat and cynomolgus macaque striatum (Seeger et al. 2003; Xie et al. 2006). Striatal medium spiny neurons are the principal input site of the basal ganglia, a series of interconnected subcortical nuclei that integrate widespread cortical input with dopaminergic signaling to plan and execute relevant motor and cognitive patterns while suppressing unwanted or irrelevant patterns (Graybiel 2000). Thus, expression of PDE10A in these neurons initially suggested a role for this enzyme in the regulation of information processing by the basal ganglia circuit. Evidence in support of this hypothesis is beginning to emerge from studies with mice in which the gene for PDE10A has been disrupted (Siuciak et al. 2006). Furthermore, these studies suggest that PDE10A inhibitors may provide a novel therapeutic approach to the treatment of psychosis.
Cellular localization and anatomic distribution of PDEs are important steps in understanding their functions and potential therapeutic applications. PDE expression in a particular cell type could suggest potential effects of a PDE inhibitor in vivo. For example, high levels of PDE5 expression in corpus cavernosum and lung are consistent with the therapeutic benefits in treating erectile dysfunction and pulmonary hypertension, respectively (Gopal et al. 2001; Corbin et al. 2005). Tissue distribution of PDE10A expression across different central and peripheral tissues has not been described to date. An important consideration in pursuing PDE10A as a therapeutic target is whether the distribution pattern of the protein observed in rodent is similar in other toxicological relevant species and ultimately in humans. Thus, the aim of the present study was to compare the distribution of PDE10A protein in the CNS and in peripheral systems across mouse, rat, dog, cynomolgus, and human using a previously characterized antibody against rat PDE10A (Seeger et al. 2003; Siuciak et al. 2006; Xie et al. 2006).
Materials and Methods
Tissue Specimens
All animals used in these studies were housed in a facility accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International and used in accordance with an approved animal care and use protocol, the Institute for Laboratory Animal Research Guide for the Care and Use of Laboratory Animals, and all applicable regulations. Tissue specimens were collected from cynomolgus macaque, beagle dog, Sprague Dawley rat, and CD-1 mouse. Monkeys and dogs were euthanized with IV injections of sodium pentobarbital (Nembutal; Abbott Laboratories, North Chicago, IL) followed by exsanguination. Rats and mice were euthanized with carbon dioxide gas and exsanguinated. Animal tissues were harvested and fixed within 48 hr in 10% neutral buffered formalin. All samples were processed according to routine procedures and embedded into paraffin blocks. Tissues evaluated are listed in Table 1.
Human tissue samples were received from the Pfizer Human Tissue Bank, University of Pittsburgh, and the University of Arizona. All human samples used in this study were received with informed consent. Human brains were initially fixed in 4% paraformaldehyde, cut into slabs, and frozen for longterm storage in 30% glycol solution. Some human brain tissues were processed as free-floating sections. All other samples were fixed in 10% neutral-buffered formalin for an unknown interval, processed, and embedded into paraffin blocks.
Brain-specific Regions
The five specific brain regions evaluated from cynomolgus macaque, human, dog, rat, and mouse were the striatum, cortex, substantia nigra, cerebellum, and hippocampus. Rat and mouse brains were evaluated in the coronal plane at 2-mm intervals, whereas dog, primate, and human brains were trimmed to select the regions of interest. Human brain regions of interest were identified within specific brain slabs, trimmed, postfixed briefly in 10% neutral-buffered formalin, and processed into paraffin blocks. In some human cases, 2-cm blocks of specific regions of interest were isolated from slabs, and 40-μm-thick free-floating sections were prepared from blocks of the striatum and substantia nigra. These sections were stored in glycerol containing cryopreservative at −20C until stained.
Tissues evaluated for PDE10A protein expression
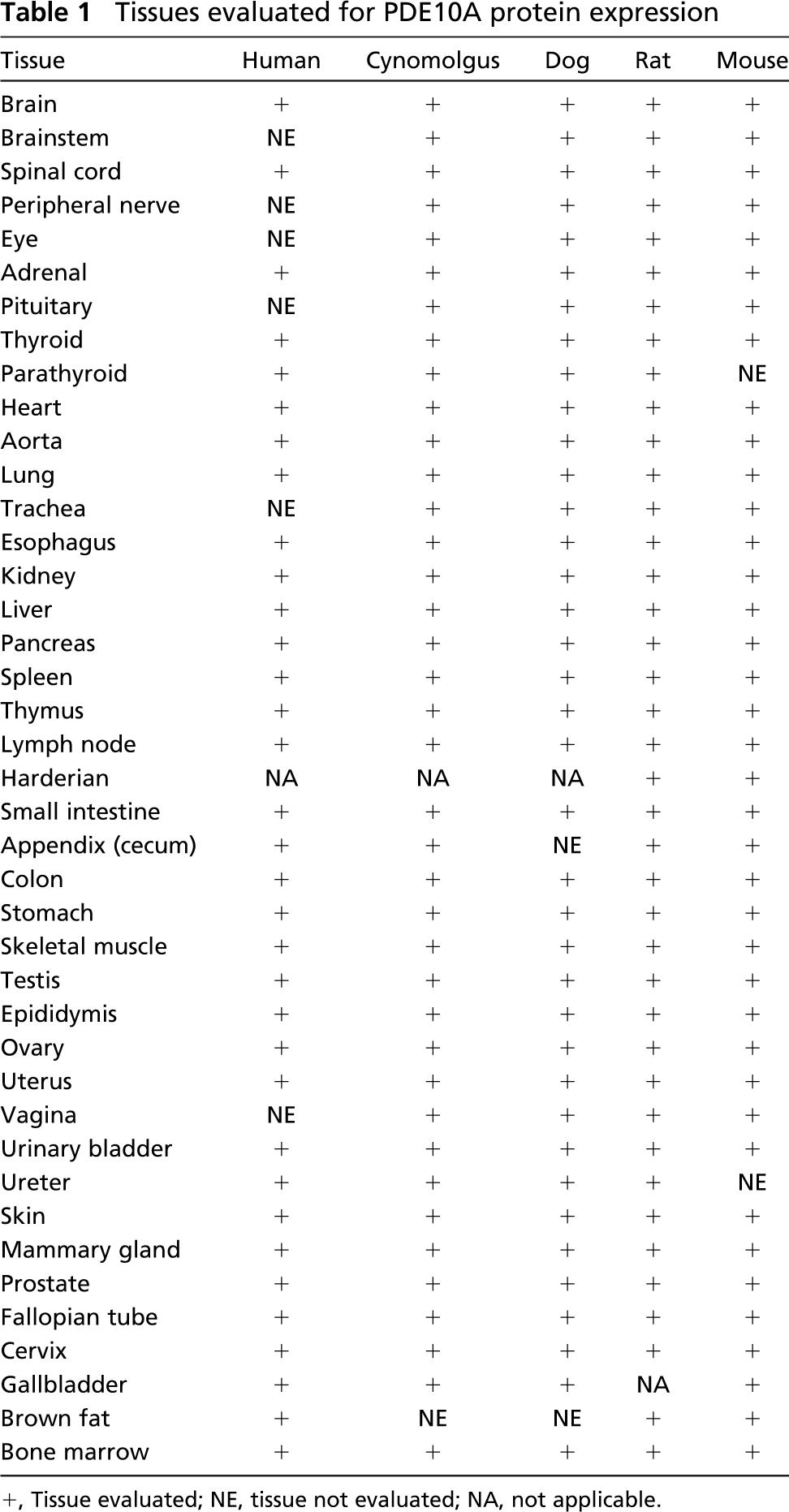
+, Tissue evaluated; NE, tissue not evaluated; NA, not applicable.
Characteristics of Monoclonal Antibody Clone 24F3.F11 Directed Against Rat PDE10A
PDE10A, clone 24F3.F11, is a mouse monoclonal antibody generated against a recombinant rat PDE10A peptide. The 24F3.F11 antibody has been previously described (Seeger et al. 2003). Briefly, using extracts from rat striatum and SF9 insect cells transfected with the rat PDE10A gene, the 24F3.F11 clone recognized a single band of 89 kDa not present in mock-transfected SF9 cells. This band was not recognized by 24F3.F11 when the antibody was preincubated with excess recombinant rat PDE10A protein. In situ hybridization demonstrated that PDE10A transcript and PDE10A immunoreactivity (PDE10A-ir) were present in similar locations (neurons of striatum, cerebral cortex, hippocampus, and cerebellum). The 24F3.F11 clone did not recognize recombinant and/or native PDE1A, PDE1B, PDE1C, PDE2, PDE3A, PDE3B, PDE4A, PDE4B, PDE5, PDE7A, PDE8A, PDE9A, or PDE11 protein in Western blots. Siuciak et al. (2006) demonstrated using Western blots that PDE10A protein of ~80-90 kDa was present in striatal extracts of wild-type mice but missing in striatal extracts of PDE10A knockout mice. Xie et al. (2006) demonstrated in extracts of cynomolgus striatum that the 24F3.F11 antibody recognizes a band identical in size to rat striatal PDE10A protein. BLAST (Altschul et al. 1997) and other available sequence data for the N terminus of the PDE10A protein (the antigenic region for 24F3.F11) demonstrate that the rat sequence is 96% identical with human, 98% with mouse, 89% with dog, and ~90% with primate PDE10A sequences, indicating that the 24F3.F11 antibody can recognize PDE10A protein from these mammalian species.
Immunohistochemistry (IHC)
Antibody binding to its cognate target sequence was verified by immunoadsorption of 24F3.F11 with PDE10A rat full-length protein. 24F3.F11 was incubated with a 25X concentration of rat PDE10A protein overnight at 4C on a rotator. The mixture was then centrifuged at 16,000 × g for 20 min at 4C to spin out the antibody-antigen complexes. The resultant supernatant was applied to brain, testis, epididymis, and enteric ganglia from cynomolgus macaque, dog, and rat. IHC was carried out in parallel with non-immunoadsorbed 24F3.F11 antibody using the method described below.
Representative tissue sections were cut at 5 μm and adhered to glass slides. All PDE10A IHC detections were performed on an automated stainer (DakoCytomation; Carpinteria, CA) using a standard avidin-biotin detection as previously described (Seeger et al. 2003). Optimal antibody concentrations for PDE10A IHC were determined by running a titration series on rat striatum and selecting the dilution that represented the best signal-to-noise ratio. For human, cynomolgus macaque, dog, and rat, 24F3.F11 was used at 1.2 μg/ml, whereas 4.8 μg/ml was used for mouse tissues. Sections were pretreated with Citra Antigen Retrieval (Biogenex; San Ramon, CA) for 20 min in a vegetable steamer. In each automated IHC run, a brain section from rat, dog, cynomolgus macaque, or humans was treated with a mouse IgG1 isotype antibody (DakoCytomation) as a negative control. To account for potential background resulting from using a mouse monoclonal antibody on mouse tissues, two slides were cut for each mouse tissue evaluated. One slide was stained with 24F3.F11, whereas the other slide received mouse IgG1 isotype as a negative control. PDE10A staining intensity was evaluated subjectively on a five-point scale: —, no staining; 1+, minimal staining clearly differentiated from the counterstained surrounding tissue; 2+, mild staining easily visible without obscuring the tissue morphology; 3+, moderate staining with relatively dense dark brown chromagen; 4+, marked staining that obscured cell morphology.
Free-floating sections of human striatum and substantia nigra were also stained with 24F3.F11. Sections stored in glycerol preservative were thawed to room temperature, rinsed in Tris-buffered saline (TBS) plus and 0.1% Triton X-100 (TBST), and incubated for 15 min in 0.6% methanolic peroxide to quench endogenous peroxidase. After further washes in TBST, sections were incubated for 30 min in TBS containing 1.5% horse serum and 0.2% Triton X-100 (TBST blocking buffer). Sections were then incubated for 24 hr at 4C on a rotator in 24F3.F11 at a concentration of 0.06 μg/ml in the same TBST blocking buffer. For detection, sections were washed with TBST and incubated for 30 min in Envision+ Polymer goat anti-mouse HRP (DakoCytomation), washed in TBST, and developed in DakoCytomation Liquid DAB+. Sections were not counterstained but mounted on slides, allowed to dry overnight, and then coverslipped.
Western Blotting
Western blot analysis was performed on extracts of striatum and cerebral cortex from rat and cynomolgus macaque as described by Xie et al (2006). Briefly, rat extracts were prepared by homogenization in lysis buffer B (50 mM Tris—HCl, pH 7.5, 250 mM NaCl, 5 mM EDTA, pH 8.0, 0.1% Nonidet P-40, and complete protease inhibitors), whereas cynomolgus brain regions were homogenized in lysis buffer from Roche Immunoprecipitation kit (Roche; Indianapolis, IN) (50 mM Tris at pH 7.5, 150 mM NaCl, 1.0% Nonidet P-40, 0.5% sodium deoxycholate with complete protease inhibitor cocktail tablets). Lysates were centrifuged at 12,000 × g for 10 min at 4C, and supernatants were stored at −80C until analysis. One μg of striatal and 10 μg of cortical extracts were loaded onto a 4-12% NuPAGE Bis-Tris gel (Invitrogen; Carlsbad, CA). Blots were probed with 24F3.F11 at a concentration of 1.2 μg/ml. As a loading control, blots were simultaneously probed with rabbit anti-actin (Sigma; St Louis, MO) at a dilution of 1:2000.
Results
PDE10A Is Highly Expressed in Mammalian Striatum
The striatum from all species examined was intensely labeled for PDE10A (Figure 1). Staining was present in the caudate nucleus and putamen with no staining of adjacent white matter (corpus callosum and internal capsule). Within the striatum, the neuropil exhibited dense and diffuse PDE10A-ir. Additionally, individual neurons in the caudate nucleus and putamen exhibited strong cytoplasmic PDE10A-ir (Figure 2). The pattern of PDE10A-ir observed in all species was consistent with previous rat studies (Seeger et al. 2003). In each case, cells positive for PDE10A comprised the majority of striatal neurons. Because ~90% of striatal neurons are medium spiny neurons, this suggests that PDE10A is highly expressed in this neuronal population across all mammalian species examined.
PDE10A Is Expressed in Substantia Nigra and Other Distinct Brain Regions
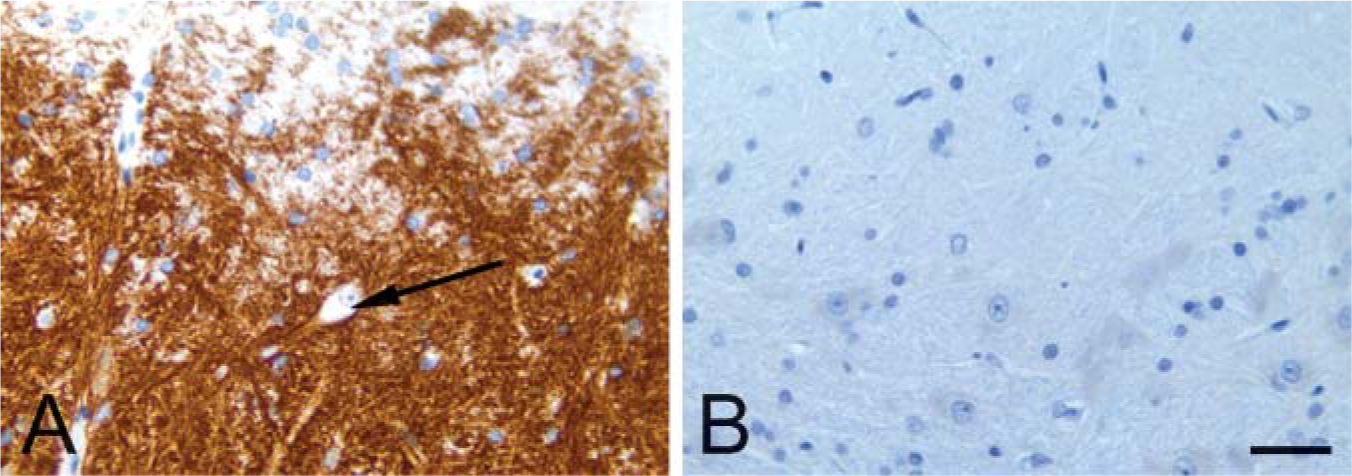
The medium spiny neurons project to both the globus pallidus and the substantia nigra (Graybiel 1990). PDE10A-ir was pronounced in the globus pallidus (Figures 1D and 1E) and the substantia nigra (Figure 3). In the substantia nigra and median forebrain bundle, PDE10A-ir was prominently expressed in fibers and terminals. Staining in the substantia nigra was confined to the pars reticulata and not present in the pars compacta. In contrast to the striatum, neuronal cell bodies in the substantia nigra were devoid of PDE10A-ir in all species examined (Figure 4). PDE10A-ir was restricted to large nerve fiber tracts and gray matter neuropil and the ventromedial white matter bundles within the putamen and globus pallidus. This demonstrates that PDE10A-ir is present in the axons of the striatal medium spiny neurons as they course toward the terminal regions of the globus pallidus and substantia nigra.
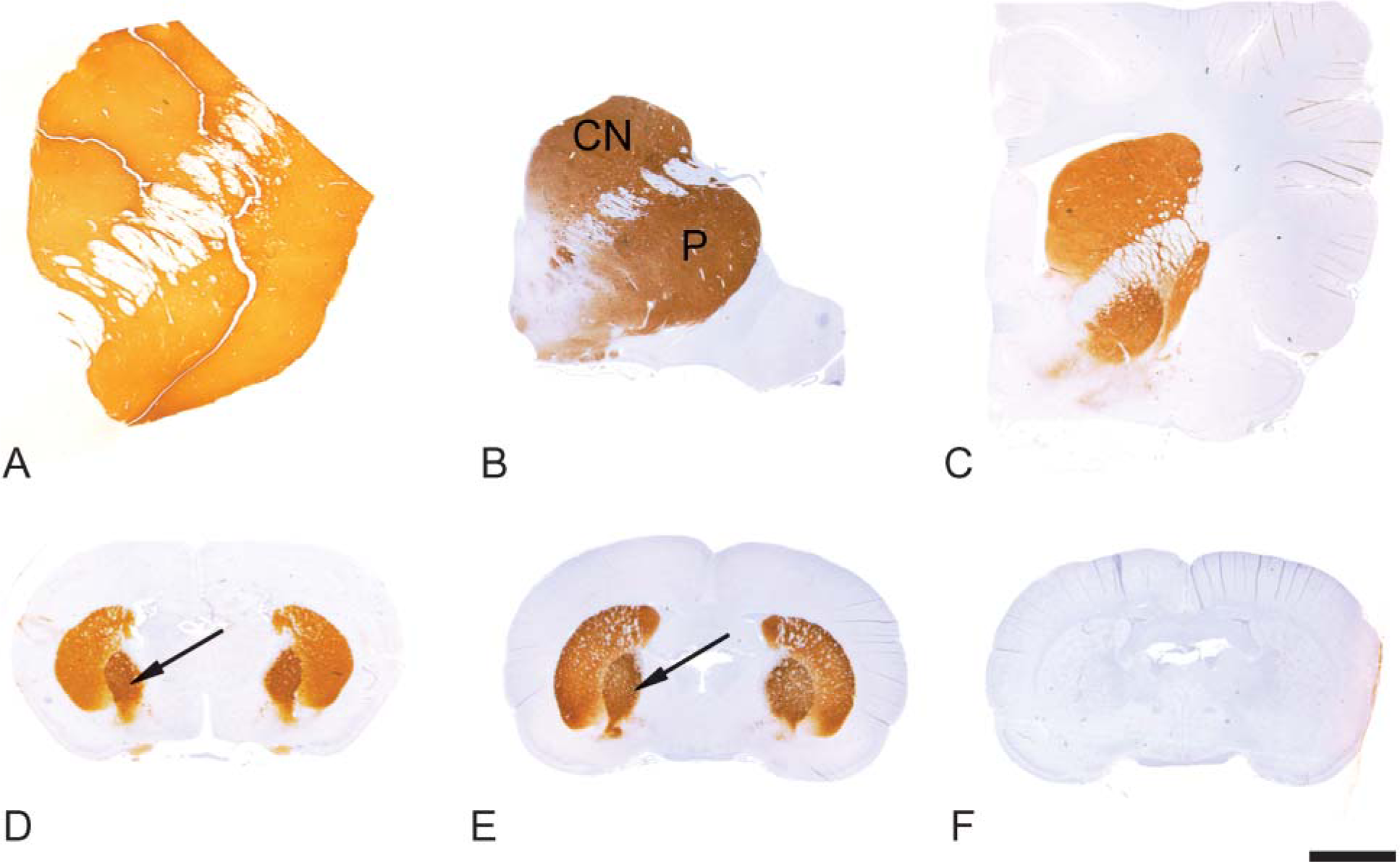
Immunohistochemical staining of mammalian striatum with anti-PDE10A. (
Outside the striatum and substantia nigra, PDE10A-ir was confined to individual neuronal nuclei. Multiple brain regions including the hippocampal pyramidal cell layer, dentate gyrus, cerebellar granular layer, and various cortical regions as well as individual neurons within the brainstem and spinal cord exhibited nuclear PDE10A localization in all species. In these widely distributed areas, PDE10A-ir was restricted to nuclei with no detectable signal in neuronal cytoplasm, glial cells, or neuropil. Immunoadsorption with excess rat PDE10A protein abolished the nuclear staining (data not shown). The intensity of nuclear PDE10A-ir in individual CA1-CA4 hippocampal pyramidal neurons varied across species (Figure 5). The number of PDE10A neuronal nuclei in mice was less than in other species, and the staining in these neurons was very faint in comparison to the other species examined.
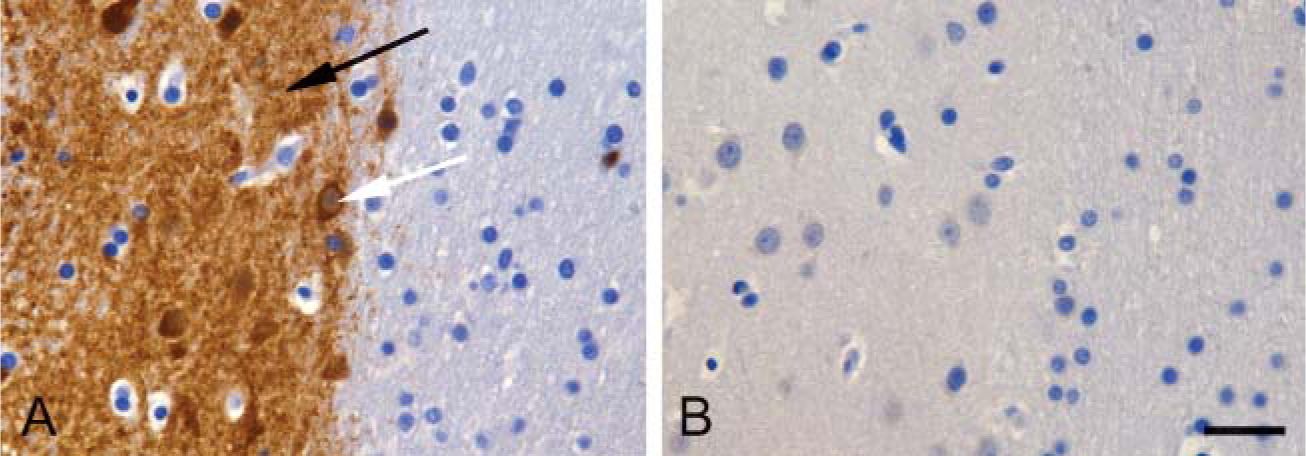
PDE10A staining of striatal neuropil (black arrow) and neurons (white arrow) of cynomolgus macaque. The region depicts the border between the putamen and the corpus callosum. (
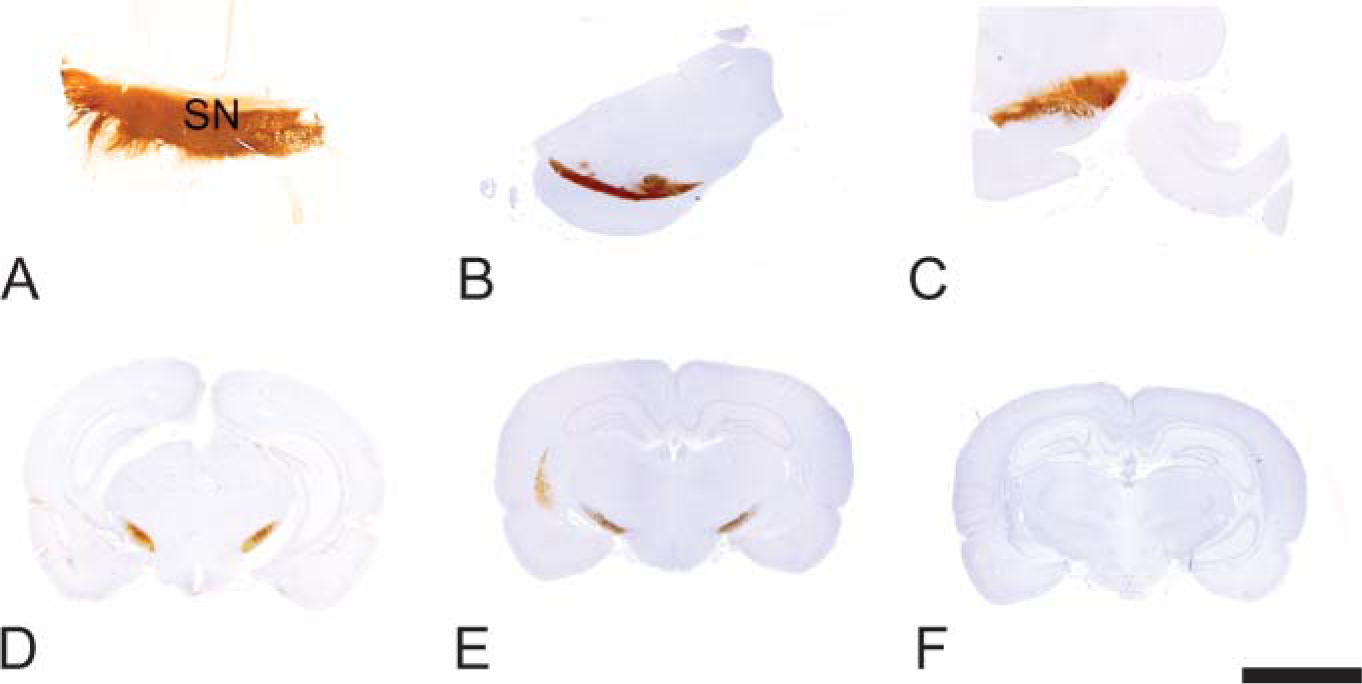
Immunohistochemical staining of mammalian substantia nigra with anti PDE10A. (
PDE10A Expression in Peripheral Tissues
Outside the CNS, PDE10A mRNA was previously shown to be expressed in human, rat, and mouse testis (Fujishige et al. 1999a,b; Soderling et al. 1999). In the current study, male reproductive tissues from all species were run in parallel to assess their expression of PDE10A protein. PDE10A-ir was detected in seminiferous tubules and epididymal sperm of dog and cynomolgus macaque and in the testis of mouse, with no detectable immunoreactivity in testes or epididymides of rat or human (Figure 6). Dog testis showed moderate specific PDE10A-ir in round and elongated spermatids and to a lesser degree in the more mature spermatocytes. In the cynomolgus macaque, testis and epididymal sperm exhibited the same pattern of PDE10A-ir seen in the dog. PDE10A-ir in the mouse testis was similar to cynomolgus macaque but was absent in mouse epididymal sperm. In the epididymides of dog and cynomolgus macaque, sperm midsections, tails, and distal cytoplasmic droplets were positive for PDE10A-ir. Less-mature sperm in the head of the epididymis contained PDE10A in tails and distal droplets, whereas more-mature sperm in the tail of the epididymis only expressed PDE10A in the distal droplets.
In the majority of the peripheral tissues PDE10A-ir was absent, demonstrating that PDE10A is not widely expressed outside the CNS. PDE10A-ir was not observed in major organs (liver, kidney, lung, spleen, or heart) of any species. With the exception of cynomolgus macaque urinary bladder epithelium, the few tissues that were positive exhibited a nuclear PDE10A staining pattern. In cynomolgus macaque, PDE10A immunoreactivity was also observed in neuronal nuclei within the wall of the urinary bladder and in the cytoplasm of the apical layers of bladder urothelium (Figures 7A and 7B). Nuclear PDE10A staining was noted in some endocrine organs (pituitary of cynomolgus macaque and rat and thyroid and parathyroid of rat). Pituitary staining was confined to epithelial cells of the anterior lobe (rat) or anterior and intermediate lobes (cynomolgus macaque), whereas thyroid PDE10A-ir was present in C-cells (rat). With the exception of the mouse, individual enteric ganglion neurons along the length of the digestive tract (esophagus to colon) exhibited nuclear PDE10A-ir, whereas mucosal epithelia, submucosa, glands, and smooth muscle were negative. PDE10A nuclear staining in pituitary and intestinal ganglia of cynomolgus macaque are illustrated in Figures 7C and 7D. A summary of the PDE10A-ir with the 24F3.F11 clone is shown in Table 2 for human, cynomolgus macaque, dog, rat, and mouse tissues.
Rat substantia nigra demonstrating strong PDE10A staining of fibers and terminals, with no staining in neurons (black arrow). (
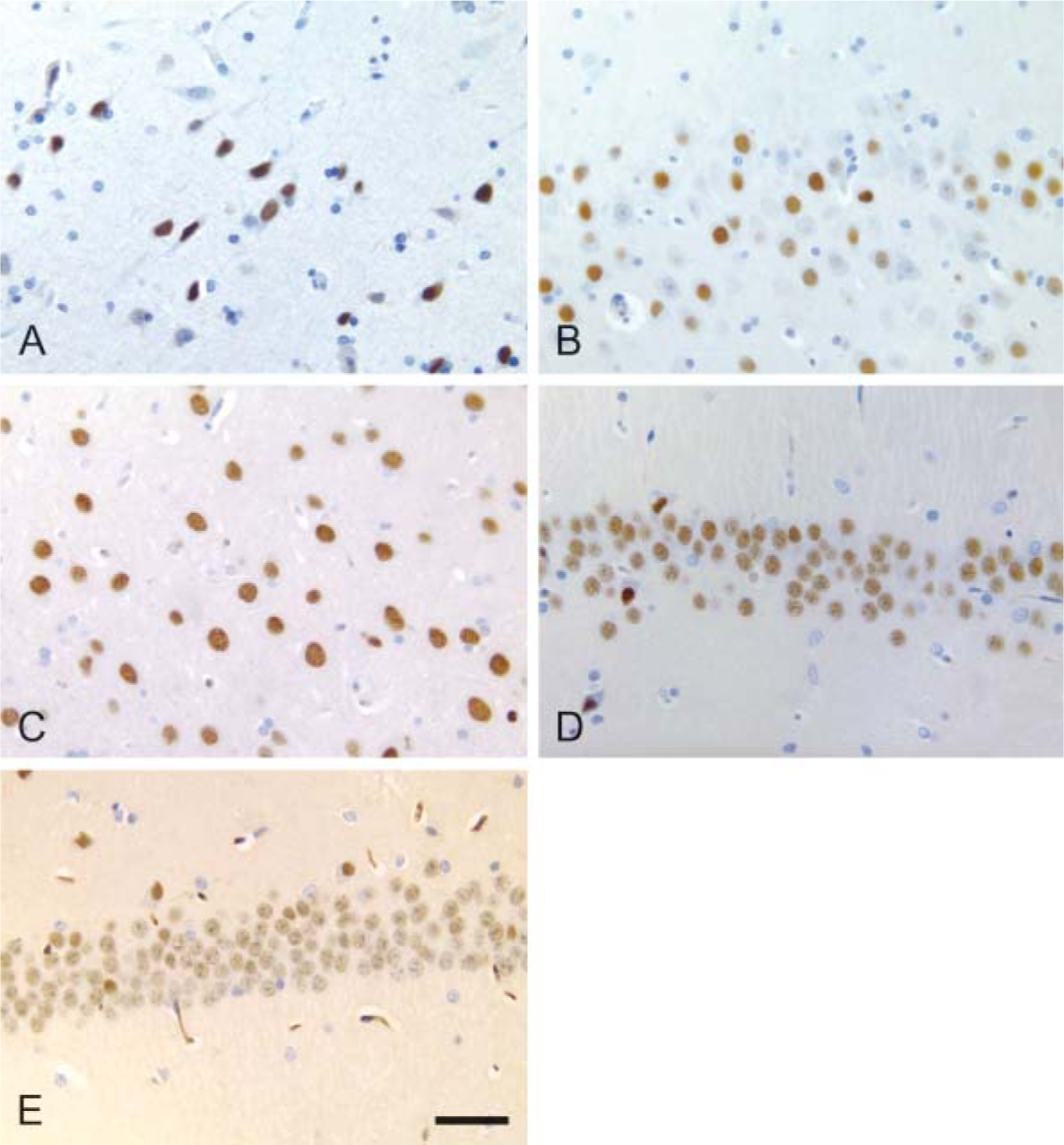
Hippocampal pyramidal neurons exhibit distinct nuclear PDE10A staining in all species. Note the variation in packing density of pyramidal neurons across species. Most hippocampal neurons are positive for PDE10A, and occur in varying intensity across species. Cynomolgus macaque, dog, and rat show strong nuclear PDE10A-ir, human is variable, and mouse is faint. (
Western Blot Analysis Demonstrates PDE10A Protein in Cynomolgus Macaque Striatum and Cerebral Cortex
To demonstrate that neuronal nuclear PDE10A-ir corresponds to PDE10A, Western blotting was performed on cynomolgus macaque and rat striatum and cerebral cortex. A single band of 89 kDa was observed in cynomolgus striatum and cerebral cortex that comigrated with comparable bands in rat (Figure 8). Previously, PDE10A protein was demonstrated in extracts of rat cerebral cortex by immunoprecipitation (Seegar et al. 2003). The current work demonstrates PDE10A protein in cynomolgus cerebral cortex, supporting the hypothesis that neuronal nuclear staining seen in all species examined is PDE10A.
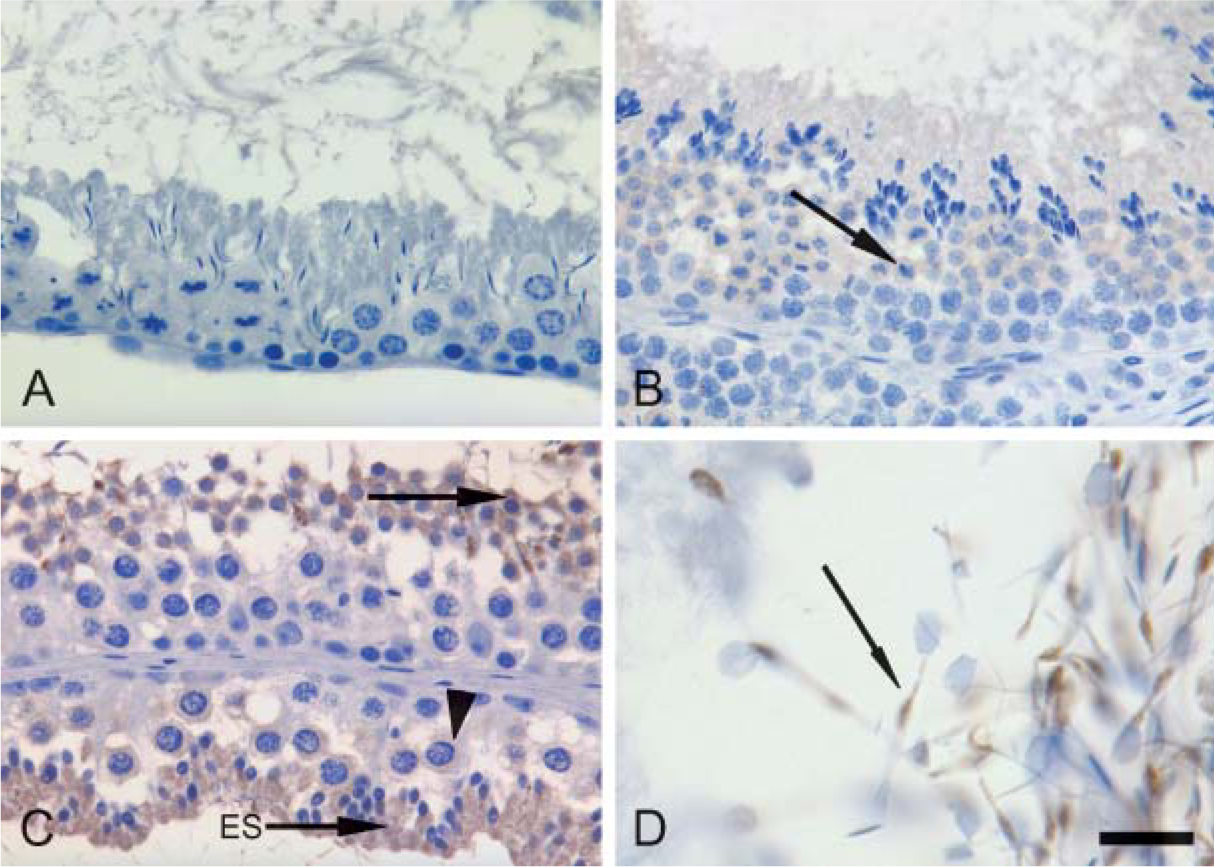
PDE10A expression varies in intensity in mammalian testis. (
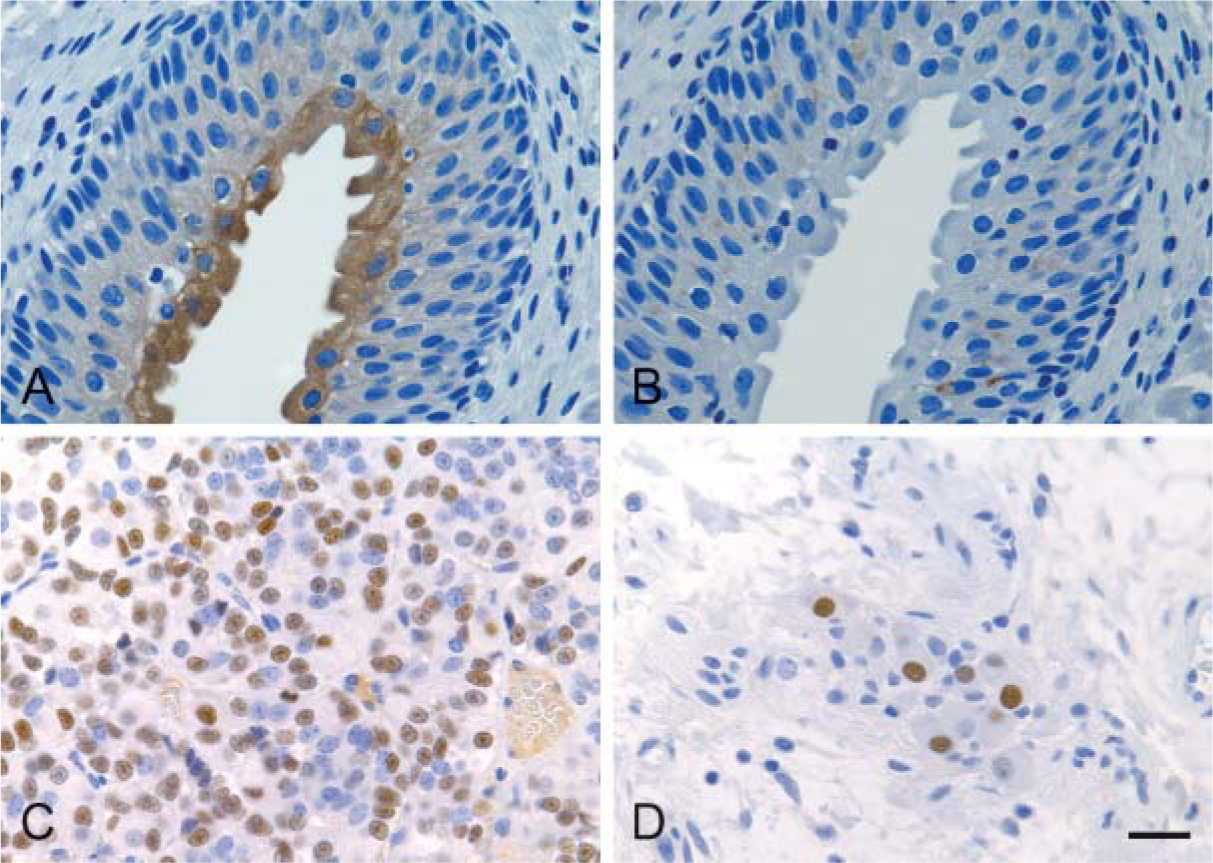
PDE10A is found in discrete areas outside the CNS. Representative images from selected cynomolgus macaque tissues are illustrated. (
Discussion
This study demonstrates that PDE10A is conserved and highly expressed in the striatonigral pathway across several mammalian species. The 24F3.F11 clone demonstrated consistent PDE10A immunoreactivity in the caudate, putamen, globus pallidus, substantia nigra, and fibers of the striatonigral pathway in mouse, rat, dog, cynomolgus macaque, and human. Cellular PDE10A staining was localized in neuronal cell bodies and neuropil of the striatum, efferent nerve fibers originating in the striatum, and neuropil of the substantia nigra. Neuronal nuclear PDE10A protein expression was also demonstrated in many brain regions of all species examined and intestinal ganglion cells of all species except mouse. Brain regions with the most prominent neuronal nuclear staining were cerebral cortex, cerebellum, and hippocampus. Nuclear staining was also seen in rat endocrine organs (pituitary, thyroid, and parathyroid), as well as cynomolgus macaque pituitary and neuronal cells in the urinary bladder wall. Apical cytoplasmic staining was observed in the urinary bladder epithelium in the macaque. In peripheral tissues, additional cytoplasmic PDE10A immunoreactivity was observed only in the testis of mouse, dog, and cynomolgus macaque. Most peripheral tissues were negative for PDE10A-ir. This is in contrast to prior reports demonstrating other PDEs to be abundantly expressed throughout the body (Beavo 1995).
Tissues exhibiting positive labeling for PDE10A
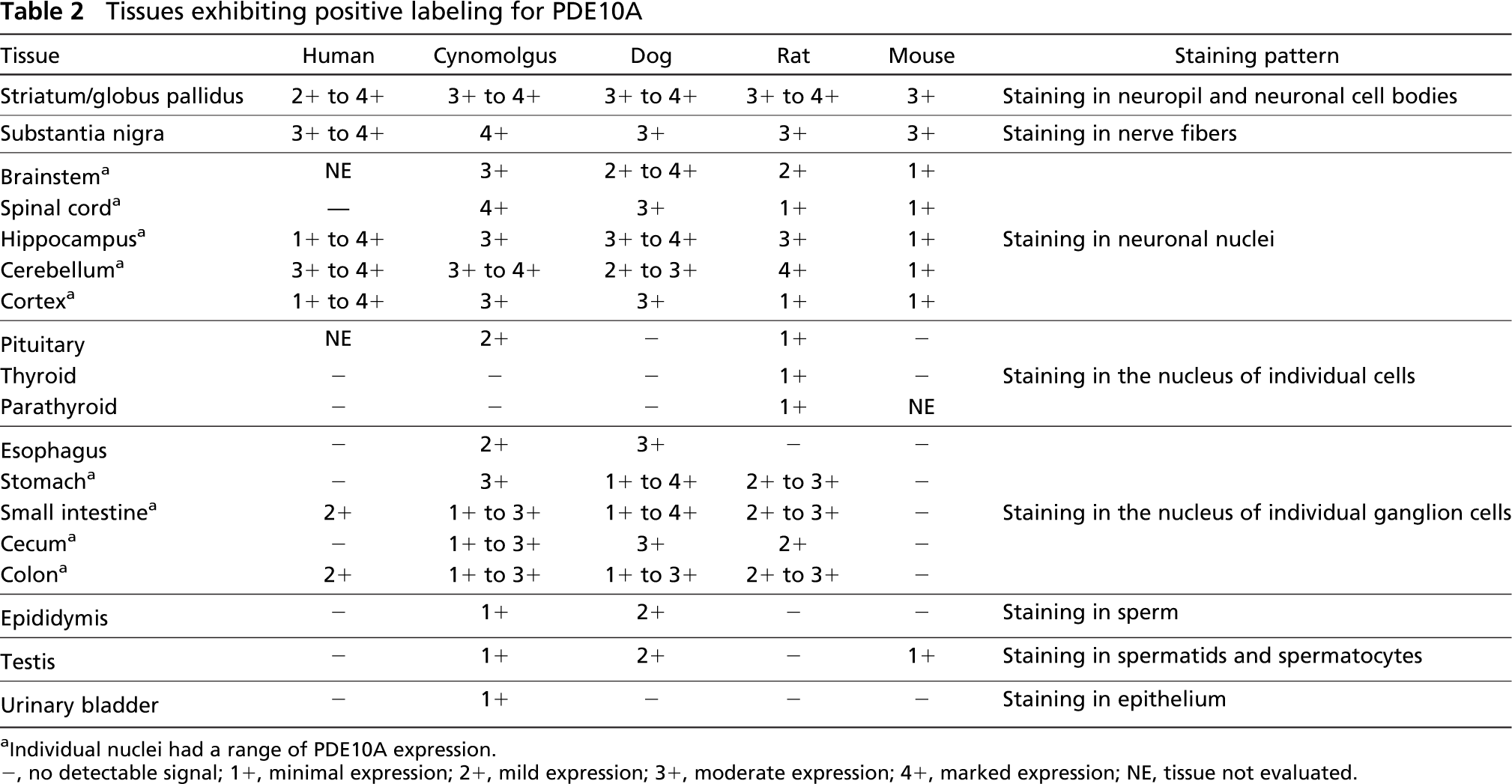
Individual nuclei had a range of PDE10A expression.
-, no detectable signal; 1+, minimal expression; 2+, mild expression; 3+, moderate expression; 4+, marked expression; NE, tissue not evaluated.
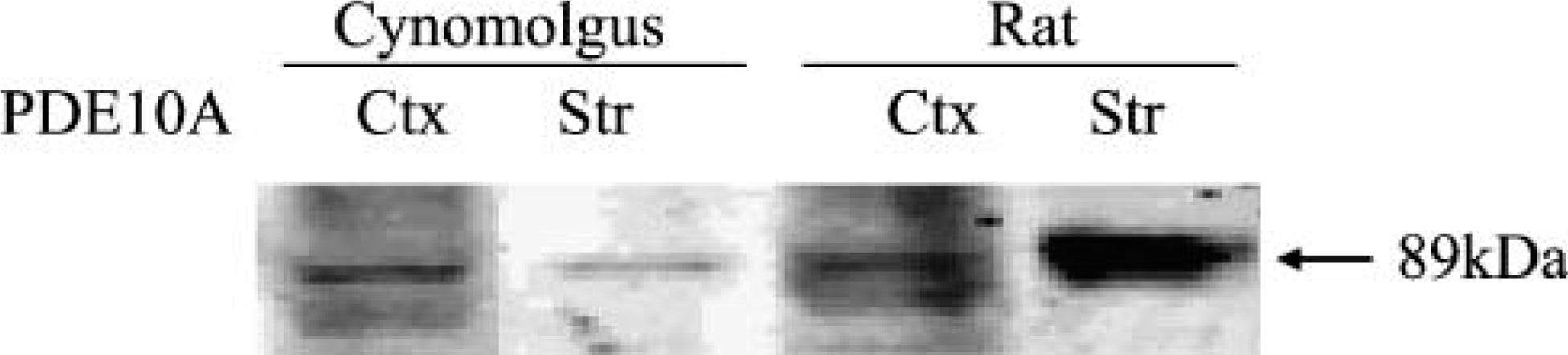
Western blot analysis of PDE10A in rat and cynomolgus macaque brain. 24F3.F11 reveals a single band of 89 kDa in cerebral cortex and striatal extracts from both rat and cynomolgus macaque. To visualize all bands in a single exposure, samples were loaded with different quantities of protein. A total of 1 μg was loaded for striatal extracts, whereas 10 μg was loaded for cerebral cortex extracts. Ctx, cortex; Str, striatum.
This is the first report of the distribution of PDE10A-ir within neuronal nuclei across several mammalian species. Several lines of evidence suggest that the nuclear localization of PDE10A-ir is specific for PDE10A: 1) PDE10A mRNA is prominently expressed in neurons of the rat cerebral cortex, cerebellum, and hippocampus (Seeger et al. 2003). These are the same areas that show prominent nuclear PDE10-ir. 2) PDE10A protein of 89 kDa is present in rat and cynomolgus macaque cerebral cortex and striatum (Seeger et al. 2003 and this study). 3) The abolition of nuclear staining via immunoadsorption with excess recombinant rat PDE10A protein also demonstrates that the nuclear staining is not an artifact. 4) The 24F3.F11 clone was previously shown not to cross-react with other PDE enzymes (Seeger et al. 2003).
PDE10A mRNA has been reported in testis and in spermatozoa of humans and rats (Fujishige et al. 1999a,b; Soderling et al. 1999; Baxendale and Fraser 2005). In this study, intensity of PDE10A-ir was most prominent in dog testis (spermatocytes and spermatids), less intense in cynomolgus and mouse testis, and below the level of detection in rat and human testis. The role of PDE10A in the testis or sperm production and maturation is not known; however, viability and intact reproductive function in PDE10A knockout mice suggests that PDE10A is not essential (Siuciak et al. 2006).
Expression of PDE10A in human tissues has been demonstrated in Northern blots of striatum with weaker expression in thyroid, testis, and pituitary (Fujishige et al. 1999a). With the exception of PDE10A-ir in enteric neurons, we did not detect PDE10A immunoreactivity in other human peripheral tissues. There are several possible explanations for this discrepancy: 1) false negative results due to challenges inherent in working with human postmortem tissues (such as lack of control over postmortem interval and duration of fixation); 2) protein expression below the level of IHC detection; and 3) lack of overlap in protein and mRNA expression in tissues. Additional work is needed to determine the significance of PDE10A in human tissues.
In summary, the present study demonstrates that PDE10A protein is expressed at much higher levels in the mammalian striatum than in other tissues. This suggests a significant, evolutionarily conserved role for this enzyme in the regulation of information processing through the mammalian basal ganglia. In fact, studies in PDE10A knockout mice suggest that PDE10A is involved in regulating the activity of the basal ganglia circuit to modulate behavioral activity (Siuciak et al. 2006). PDE10A is also expressed by neurons throughout the mammalian brain, but the distribution of the protein in non-striatal neurons is restricted to the nuclear region. This suggests a second and most likely distinct function for PDE10A. Outside the CNS, PDE10A expression is low and variable across species, including in testis where high levels of mRNA are expressed. The function of PDE10A in peripheral tissues has not been elucidated.
Footnotes
Acknowledgements
We thank Alan Opsahl for assistance in preparing the figures used for this study. We also thank Mary Payette, Jill Long, and Renee Huynh for performing immunohistochemistry.