
Abstract
Sperm protein 17 (Sp17) is a highly conserved mammalian protein whose primary function is still poorly understood. Immunohistochemistry (IHC) in the human testis reveals the presence of Sp17 in some spermatocytes and abundantly in spermatids. All spermatogonia, Sertoli cells, and Leydig cells appear to be immunonegative for Sp17, whereas some interstitial cells are immunopositive. IHC recognized two distinct populations (immunopositive or not for Sp17) in the ejaculated spermatozoa. Although it will be necessary to clarify why some ejaculated spermatozoa do not contain Sp17, its distribution suggests that this protein may be associated with some phases of germinal cell differentiation.
T
A family of low molecular weight sperm autoantigens (RSA) has been detected in rabbit spermatogenic cells and spermatozoa and was shown to bind the carbohydrate components of the zona pellucida (Lea et al. 1998). A 17-kD mannose-binding protein was the first RSA to be isolated, sequenced, and characterized in rabbit (O'Rand et al. 1988). It is now called sperm protein 17 (Sp17) because it is also found in mouse, monkey, baboon, macaque, and human (HSp17) testis and spermatozoa (Kong et al. 1995; Lea et al. 1996,1998). HSp17 has a high degree of homology (70% over the coding region sequence) with rabbit and mouse Sp17, and 97% homology with baboon Sp17 (Wen et al. 2001).
Because no data are yet available concerning the immunolocalization of HSp17 in human primate testis and ejaculated spermatozoa, and the suggested potential role of Sp17 as a key-target in human immunotherapy (Lim et al. 2001; Chiriva–Internati et al. 2001; Grizzi et al. 2002), this study aimed at investigating the topography of HSp17 in human spermatogenic cells identifiable in testis biopsy specimens, surgical specimens, and ejaculated spermatozoa.
The study was carried out in accordance with the guidelines of the Ethics Committee of the hospital treating the patients, all of whom were fully informed of the possible discomfort and risks of surgical treatment. None of the specimens (n = 10) represented benign or malignant tumors or damaged tissues, as confirmed by the histological analyses independently performed by two pathologists.
The specimens were fixed in 10% neutral buffered formalin (∼12 hr) and embedded in paraffin. The histological analysis was performed on 2-μm hematoxylin and eosin (H&E)-stained sections under a light microscope (Leica DMLA; Milan, Italy). The immunolocalization of HSp17 in human testis and ejaculated spermatozoa was investigated by using a self-produced recombinant Sp17 protein to immunize a BALB/c mouse to produce mouse Sp17 monoclonal antibodies (MAbs) from the immunized splenocytes. The successful generation of mouse Sp17 MAbs was first confirmed by Western blotting analyses demonstrating their ability to bind the recombinant Sp17 but not a control recombinant protein produced in an identical manner (Chiriva–Internati et al. 2001; Lim et al. 2001; Grizzi et al. 2002). Both of the recombinant proteins were derived from Escherichia coli and contained a C-terminal 6-His tag. The antibodies are mouse IgG1 subclass.
A further Western blot using this antibody was made on a testis homogenate and sperm lysate to demonstrate the specificity of this antibody on the labeled tissues. Testis homogenate and sperm lysate were separated under reducing and denaturing conditions on a 10% polyacrylamide gel, and transferred onto nitrocellulose membranes (Burnette 1981). The membranes were incubated overnight at 4C with the MAb to Sp17 and the immunoreactive bands revealed by the Opti−4CN substrate (Amplified Opti−4CN; Bio Rad, Milan, Italy).
Consecutive 2-μm sections were cut and processed for IHC. After dewaxing and rehydration, the sections were autoclaved for antigen retrieval for 15 min at 121C in a freshly made 1 mM EDTA solution, incubated with 3% H2O2 for 30 min to quench endogenous peroxidase activity, and then treated with either primary antibody raised against HSp17 at room temperature for 2 hr (monoclonal mouse anti-HSp17, dilution 1:100 in PBS) or with 1 mg/ml mouse IgG1 (DAKO; Milan, Italy) as a negative control. This was followed by 30-min incubation with the DAKO Envision system, which is based on a unique enzyme-conjugated polymer backbone that also carries secondary antibody molecules. 3,3′-Diaminobenzidine tetrahydrochloride (Sigma; St Louis, MO) 12.5 mg and 500 μl H2O2 in 50 ml TRIS-buffered saline was used as a chromogen to yield brown reaction products. The nuclei were lightly counterstained with hematoxylin solution (Medite; Bergamo, Italy). Experiments were performed independently three times to confirm the findings. Cytological samples of acrosome-intact ejaculated spermatozoa obtained from four healthy donors with a mean age of 27 ± 5.19 years were independently analyzed by two expert cytologists.
To investigate the immunocytochemical expression of HSp17, washed spermatozoa were cytocentrifuged for 1 min at 800 rpm on a glass slide using a Cytospin-2 centrifuge (ALC pk130; Milan, Italy) and then fixed with Biofix (Bio-Optica; Milan, Italy). The cells were permeabilized with 0.5% Triton X-100 (Sigma), 0.1% sodium citrate in PBS at 4C for 15 min, followed by treatment with either a primary antibody at RT for 2 hr or with 1 μg/ml mouse IgG1 (DAKO) as a negative control. This was followed by 30-min incubation with the DAKO Envision System. 3,3′-Diaminobenzidine tetrahydrochloride (Sigma) 12.5 mg and 500 μl H2O2 in 50 ml of TRIS-buffered saline was used as a chromogen to yield brown reaction products. Experiments were performed three times to confirm the results. Almost 1000 spermatozoa were counted under a light microscope and the number of those immunopositive for HSp17 was expressed as the mean percent (±SD) of all spermatozoa.
Western blotting analysis has confirmed the successful generation of mouse Sp17 MAbs by showing their binding to a recombinant Sp17 but not to a control recombinant protein (data not shown). Figure 1 shows the Western blotting analysis on a testis homogenate and spermatozoa lysate, which demonstrates the specificity of the antibody on the tissue being labeled.
Although a large number of tubules were highly immunopositive for HSp17 (Figure 2A), some seminiferous tubules were negative (Figure 2B). At higher magnification (Figures 2C and 2D), all of the spermatogonia and Sertoli cells were immunonegative for HSp17, but it was weakly observed in the cytoplasm of some spermatocytes (Figure 2E) and strongly present in the cytoplasm of early and late spermatids (Figure 2F). All tails of the spermatozoa identified in the lumen of the seminiferous tubules were also strongly positive. Although all of the observed Leydig cells were immunonegative, some interstitial cells were immunopositive for HSp17 (Figure 2A). The IgG controls were always negative.
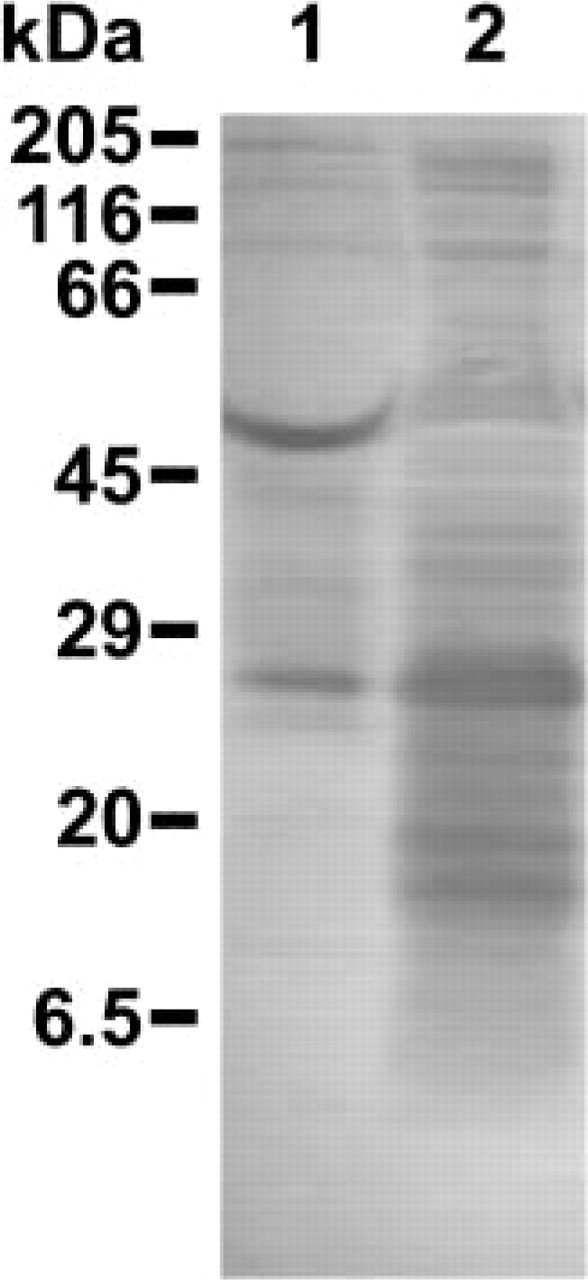
Western blotting analysis of human testis homogenate (Lane 1) and sperm lysate (Lane 2). Immunoreactive bands were obtained with the affinity-purified mouse anti-rHSp17 (dilution 1:1000). Reference molecular weights are indicated at left. According to Lea et al. (1998), a triplet of proteins at 24.5, 22.6, and 22.1 kD were detectable. Additional bands of 20 and 19.1 kD were also detected in Lane 2. A multimeric band was strongly detected at 53.7 kD in Lane 1 and faintly detectable in Lane 2. It is still unclear how the native protein is modified to produce the triplet (22.1– 24.5 kD) or what function is served by the existence of the different forms. Lea et al. (1998) stated that the native human Sp17 exists as a triplet of proteins with a unique nucleotide and deduced amino acid sequence.
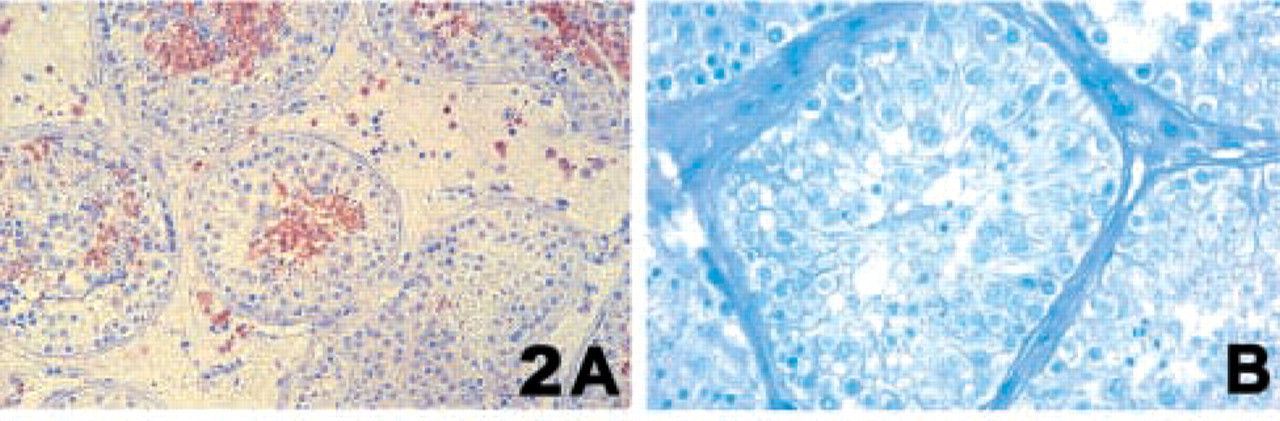
Immunolocalization of HSp17 in human testis (
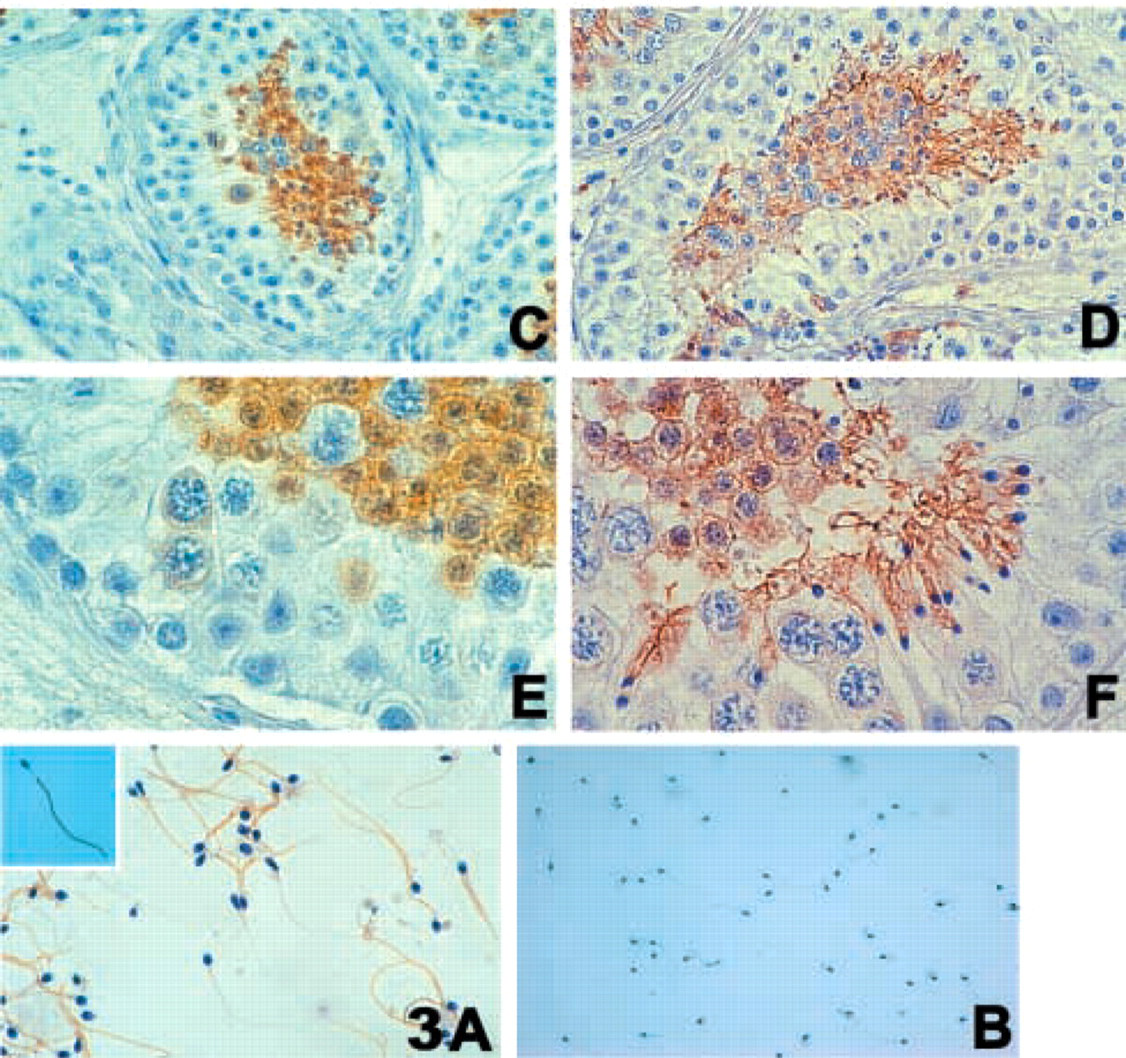
(
Two distinct HSp17 populations, one immunopositive and the other immunonegative (Figure 3A), were recognized in the ejaculated spermatozoa obtained from healthy donors. The immunopositive spermatozoa accounted for 90 ± 5% of all spermatozoa.
Observed using light microscopy (final magnification X1000), HSp17 was clearly detected throughout the principal piece of the flagellum, but the intermediate piece, head, and acrosome vesicle appeared to be immunonegative (Figure 3A, inset). Control IgG was always negative (Figure 3B).
To achieve fertilization, human spermatozoa and oocytes are characterized by the presence of specific molecules that mediate the complex steps of their interactions (Primakoff and Myles 2002). The first interaction between mammalian spermatozoa and oocyte occurs at an oocyte-specific extracellular matrix called the zona pellucida. A large number of specific binding molecules have been widely described and, in rabbit, a family of molecules (RSAs) has been structurally and functionally characterized (O'Rand et al. 1988). These low molecular weight rabbit sperm antigens include a protein of 22–24 kD known as Sp17.
Although Kong et al. (1995) and Adoyo et al. (1997) have recently detected Sp17 in mouse and baboon testis, its presence in human primate testis and ejaculated spermatozoa has not yet been investigated. The present study was driven by the increased number of investigations on Sp17 as a key target for human immunotherapy (Chiriva–Internati et al. 2001; Lim et al. 2001; Grizzi et al. 2002). Although Sp17 was originally described as a protein involved in the rabbit fertilization process, the immunolocalization ascertained in our study indicates a heterogeneous distribution of HSp17 in human testis and a regional localization in a subpopulation of ejaculated spermatozoa. These findings suggest that HSp17 may be associated with the differentiation processes of germinal cells from spermatocytes to completely mature spermatozoa. This hypothesis is suggested by the fact that the very small number of seminiferous tubules immunonegative for HSp17 seemed visually characterized only by the presence of spermatogonia and Sertoli cells (Figure 2B). During spermiogenesis, the most important changes involve the formation of the flagellum, the condensation of the nucleus, the formation of the acrosome, and the elimination of excess cytoplasm. Because Sp17 has been suggested to be a fundamental factor in the molecular aspects of sperm-to-ovum binding (O'Rand et al. 1988) and the tail in the penetration of the zona pellucida (Primakoff and Myles 2002), the presence of this protein throughout the principal piece of the flagellum indicates a synergy of these two components in the fertilization process. However, because some healthy human ejaculated spermatozoa do not label HSp17, further studies will be necessary to clarify the functional significance of the two different populations and why one does not label HSp17.
Kong et al. (1995) have shown that Sp17 mRNA is not present in any of the examined mouse somatic tissues (heart, brain, spleen, lung, liver, skeletal muscle, and kidney) but is specific to the testis. However, emerging evidence indicates that in humans it may be more widely expressed than previously thought, and that its expression is highest in the testis (Lim et al. 2001; Chiriva–Internati et al. 2001). This is in accord with our findings that some interstitial cells seemed to be immunopositive for HSp17.
Lacy and Sanderson (2001) have demonstrated an aberrant expression of normal testicular proteins, such as HSp17, on the surface of malignant lymphoid cells (including B- and T-lymphoid cell lines), and on the surface of primary cells isolated from B-lymphoid neoplasm. HSp17 and other testicular proteins have recently been collectively classified as a new family of tumor antigens called cancer-testis (CT) antigens (Lim et al. 2001). Wen et al. (2001) have reported that HSp17 plays a role in cell–cell adhesion and/or cell migration in transformed, lymphocytic, and hematopoietic cells, possibly as a result of its interaction with extracellular heparan sulfate. The widely aberrant expression of HSp17 in cancer tissues and its restricted expression in normal tissues suggest that HSp17 may be a key target for immunotherapy without damaging the spermatogonia, which are the first cells involved in the spermatogenic process.
In conclusion, previous investigation in baboons (Adoyo et al. 1997) and these exciting new findings in human primates with their intriguing clinical perspectives suggest further electronic microscopy- and biochemistry-based experimental studies. Moreover, it is necessary to elucidate the possible roles played by the highly conserved HSp17 (Frayne and Hall 2002), in an attempt to determine why HSp17 is not present in some human ejaculated spermatozoa.
Footnotes
Acknowledgements
Supported by the National Institute of Health/National Cancer Institute (RO1 CA 88434), the Cancer Treatment Research Foundation, the Mary Kay Ash Charitable Foundation, and the Fondazione “Michele Rodriguez” Istituto Scientifico per le Misure Quantitative in Medicina.
We are very grateful to Prof Massimo Roncalli of the Pathology Unit, the core facility of the Laboratory for Reproductive Biology of the Istituto Clinico Humanitas, Dr Nicoletta Gagliano of the Department of Human Anatomy, University of Milan, and Dr Ruth Tapparo for their help and expertise.