
Abstract
High levels of plasma estrogens constitute an endocrine peculiarity of the adult stallion. This is mostly due to testicular cytochrome P450 aromatase, the only irreversible enzyme responsible for the bioconversion of androgens into estrogens. To identify more precisely the testicular aromatase synthesis sites in the stallion, testes from nine horses (2–5 years) were obtained during winter or spring. Paraplast-embedded sections were processed using rabbit anti-equine aromatase, followed by biotinylated goat anti-rabbit antibodies, and amplified with a streptavidin-peroxidase complex. Immunore-activity was detected with diaminobenzidine. Immunofluorescence detection, using fluoroisothiocyanate-conjugated goat anti-rabbit antibodies, was also applied. Specific aromatase immunoreactivity was observed intensely in Leydig cells but also for the first time, to a lesser extent, in the cytoplasm surrounding germ cells at the junction with Sertoli cells. Interestingly, the immunoreactivity in Sertoli cells appears to vary with the spermatogenic stages in the basal compartment (with spermatogonia) as well as in the adluminal one (with spermatids). Relative staining intensity in Leydig and Sertoli cells and testicular microsomal aromatase activity increased with age. The present study in stallions indicates that in addition to Leydig cells, Sertoli cells also appear to participate in estrogen synthesis, and this could play a paracrine role in the regulation of spermatogenesis.
S
The stallion is commonly considered to be a seasonal breeder, with a breeding season extending in general from March to July in the northern hemisphere (April to July in Normandy, France). However, unlike other seasonal breeders that cease spermatozoa production during winter, stallions continue to produce spermatozoa throughout the year. Moreover, the stallion breeding season is characterized by an increase in testicular size and weight and in Leydig, Sertoli, and germ cell numbers (mainly type A and B spermatogonia). During this time, the daily production of sperm (Johnson 1985) and of plasma and seminal sexual steroids, such as estradiol and testosterone (Berndtson et al. 1974; Lemazurier et al. 2002a), are also enhanced.
In addition to seasonal variations, both testicular and hormonal characteristics of stallions also change with age. Horse puberty begins at 1–1.5 years of age, depending on the breed, and spermatogenesis is completed by 2 years of age (Clemmons et al. 1995). Leydig and Sertoli cell numbers are age-dependent in stallion. Adult values for daily sperm production (Johnson et al. 1991; Johnson 1995) and Sertoli cell number are reached by 4 years of age (Johnson 1995). A simultaneous increase of plasma gonadotropins (LH, FSH) and testosterone levels with age has also been reported (Johnson and Thompson 1983; Johnson et al. 1991). Similarly, a rise in intratesticular content of testosterone has been noted with age (Johnson et al. 1991).
Aromatase is the only enzyme responsible for the irreversible bioconversion of androgens to estrogens. This enzyme is considered to be crucial and rate-limiting for the estrogen/androgen balance in the body, and thus important for estrogen-dependent processes such as bone maturation (Lemazurier et al. 2002b) and reproduction, even in males (Hess et al. 1997). Thus, this enzyme has been immunolocalized in the horse testis solely in Leydig cells (Eisenhauer et al. 1994; Almadhidi et al. 1995; Nagata et al. 1998). The seasonand age-related cellular and hormonal changes observed in stallion testes should not, however, exclude the possibility of other sources of estrogens. Aromatase was initially detected only in rat Leydig cells; however, it was later localized in several species in both Sertoli and germinal cells (for review see Carreau et al. 2001).
The aim of the present study was to better define the testicular distribution of aromatase in the stallion.
Materials and Methods
Animals and Sampling
Testes from five postpubertal (2-year-old) and four adult (3–5-year-old) stallions were obtained from a veterinary clinic (Veterinary Clinic; Crèvecoeur-en-Auge, Normandy, France) after castration under general anesthesia. This was performed both during the breeding (April to June, n = 6) and non-breeding (February and March, n = 3) seasons. Tissues were immersed in NaCl 0.9% or in Eagle's minimum essential medium (EMEM) and transported to the laboratory on ice. In the laboratory, each testis was cut transversely into two equivalent pieces and either used immediately for immunohistolocalization or stored at −70C for microsomal preparation.
Chemicals and Tissue Processing
All chemicals and reagents were obtained from Sigma (St Quentin Fallavier, France) except when stated. Fragments of testicular parenchyma, 0.5 × 0.5 × 1 cm, were taken from the center and periphery of testes, fixed in Bouin's solution or 4% paraformaldehyde (PFA) in Dulbecco's PBS, pH 7.4, for 24 hr at 4C, dehydrated in ethanol, and embedded in paraplast. Tissue blocks were serially sectioned at 5–7 μm and attached to poly-L-lysine-coated slides. Slides were then dried at 37C for 48 hr before immunohistochemistry.
Immunohistolocalization
The preparation and specificity of primary rabbit anti-equine aromatase antibodies were reported previously (Almadhidi et al. 1995). Antibodies were raised in the laboratory after equine testicular aromatase purification on an immunoaffinity column (Séralini et al. in press). Slides were deparaffinized with xylene, rehydrated in graded alcohols, and washed in dd H2O and finally in PBS. They were then treated with 3% H2O2 in PBS for 60 min to inactivate endogenous peroxidase activity. After a brief wash in PBS, slides were heat-treated for antigen retrieval using citrate buffer (10 mM, pH 6) in a microwave oven at maximal power (Tecvis: 1250 W, operating at a frequency of 2450 MHz) three times for 3 min, and washed in PBS (Lan et al. 1995; Shi et al. 1997). To eliminate nonspecific binding, slides were incubated with 5% normal goat serum in PBS twice for 30 min. They were then incubated overnight with the primary antibodies diluted 1:1000 in PBS containing 1% BSA in a moist chamber at 4C. This was followed by three 5-min washes in PBS, after which slides were subjected to two different protocols, either with immunofluorescence or immunoperoxidase. For immunofluorescence, sections were incubated with fluoroisothiocyanate (FITC)-conjugated goat anti-rabbit IgG (dilution 1:60) for 1 hr in a moist and dark chamber, and then washed in PBS (three times for 5 min), and then in PBS containing 0.1% Tween, and finally in distilled water (2 min). Slides were then mounted with Fluoprep (BioMérieux; Charbonnières Les Bains, France) and microscopically observed and photographed as previously described (Almadhidi et al. 1995). For immunoperoxidase, sections were incubated with biotinylated goat anti-rabbit IgG (dilution 1:800) for 1 hr and then incubated with streptavidin-peroxidase complex (dilution 1:500) for 30 min after three 5-min washes with PBS containing 0.1% Tween. After serial washes in PBS, the reaction on the slides were developed with diaminobenzidine (DAB) in H2O2 for 4–6 min, counterstained with Harris hematoxylin, dehydrated in alcohol, and mounted with DPX. All the above incubations were carried out at room temperature except when stated. In controls, normal rabbit serum or PBS alone was applied instead of primary antibodies. In addition, primary antibodies were preadsorbed for 48 hr with 23 nmol/ml purified equine aromatase and this preparation was also used as control (Almadhidi et al. 1995). A total of 150 tubules were precisely analyzed per testis, in three serial sections from five different pieces.
Microsomal Preparations, Aromatase Activity Assays, and Data Analyses
Microsomes from each testis were prepared by successive ultracentrifugations as previously described (Dintinger et al. 1989). Protein concentrations in the preparations were determined according to Bradford (1976). Microsomal aromatase activity was evaluated by measuring 3H2O released from 100 nM [1β-3H]-androstenedione substrate (NEN, Les Ulis, France; specific activity 25.9 Ci/mmol) at 37C for 15 min. The method was adapted from Thompson and Siiteri (1974) as previously described (Dintinger et al. 1989). Six samples for each microsomal preparation were evaluated and the results were statistically analyzed by Student's t-test. All values are expressed as mean± SD.
Results
Both Bouin and PFA fixatives induced similar immunoreactivity characteristics. However, Bouin's solution was better than PFA for conservation of cell morphology. Therefore, all results presented here were obtained from tissue fixed in Bouin's solution.
Aromatase was immunolocalized in both Leydig and Sertoli cells; Leydig cells were immunostained very much more strongly than Sertoli cells (Figures 1–3). In the latter, immunostaining appears heterogeneously in the cytoplasm surrounding germ cells. However, labeling in the peripheral membrane of germ cells is not excluded. In all testes examined, staining intensity was consistent between serial sections or between staining methods (Figures 1A–1C). Higher magnification views indicate that staining was localized only in the cytoplasm of immunopositive cells; nuclei were negative (Figures 1B–1D). All other cell types in the tubular, peritubular, and interstitial compartments were immunonegative.
Cytoplasmic staining intensity in both Leydig and Sertoli cells was increased noticeably with stallion age. Leydig cells of 3- and 5-year-old stallions were stained more strongly than those of 2-year-old stallions (Figures 2A–2B). Moreover, in aged testes almost all interstitial spaces were filled by clusters of Leydig cells; this phenomenon amplified the signal. Because of their immunoreactivity with the antiserum, two types of Leydig cells were observed; one type reacted less than the other. In 2-year-old stallions, the presence of the less immunoreactive cell type was observed more frequently than in 3- or 5-year-olds. In addition, the 2-year-old stallion testes contained small clusters of Leydig cells in the interstitial spaces, and individual Leydig cells were observed more frequently (Figure 2A). No differences in immunostaining were observed between winter (February) and spring (April to June) whatever the age.
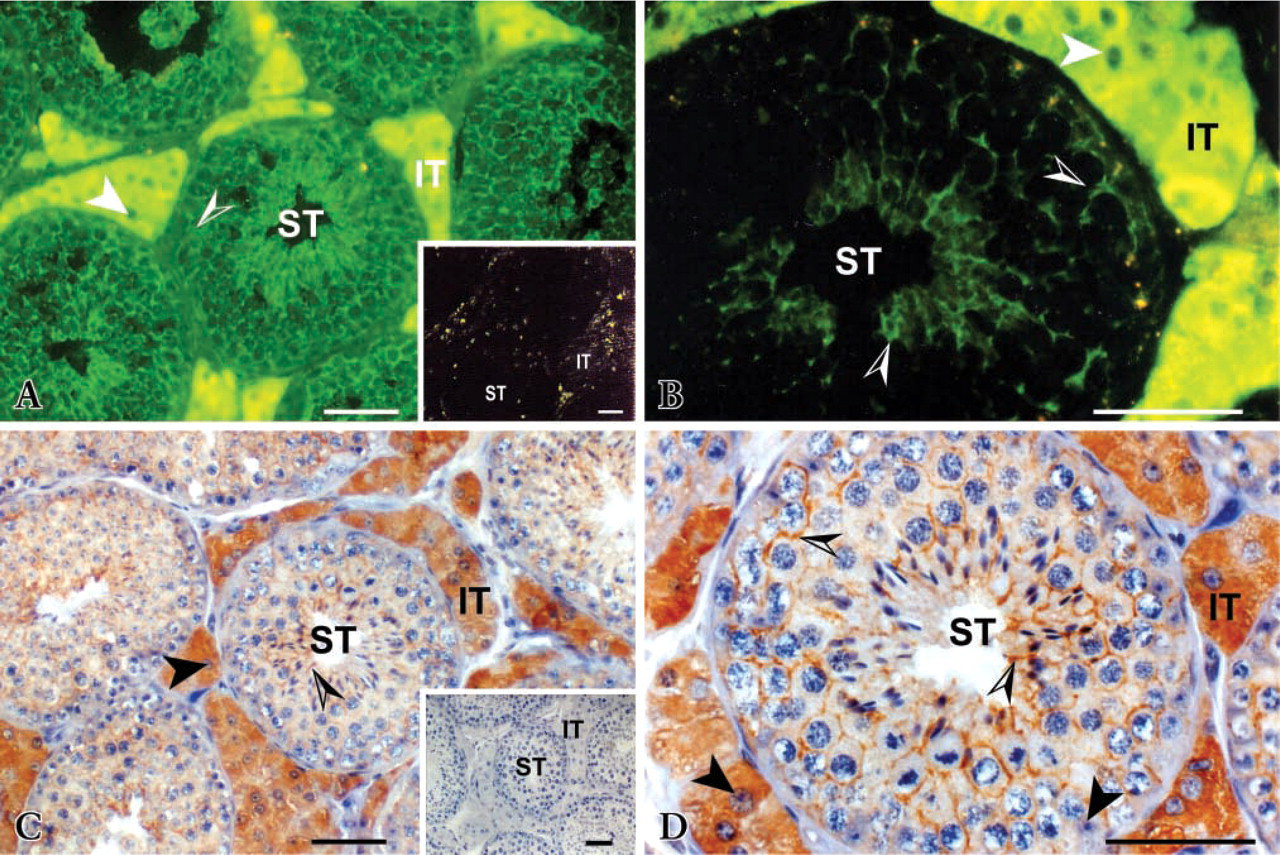
Aromatase immunostaining distribution in 3-year-old stallion testis. (
Although immunoreactivity of Sertoli cells was seen in all spermatogenic stages, some differences across the seminiferous epithelium were observed (Figure 3). In stages III and VI, the intensity of the staining was close to the basal lamina and appeared higher at the junctions of Sertoli cells, i.e., around spermatogonia and zygotene spermatocytes. In contrast, in stages IV and VII the staining was higher in the adluminal section, i.e. around the diakinese spermatocytes and elongated spermatids. In all other stages, I, II, V, and VIII, the staining was distributed uniformly across the tubules within Sertoli cells (Figure 3). The intercellular spaces between germ cells appear to be positive where Sertoli cells support germ cells. There was no difference in both distribution and intensity of the staining between testes collected from April to June in 3-year-old stallions. About 150 tubules were observed per testis.
Testicular aromatase activity increased significantly with age between 2 and 3–5 years (Figure 4A; p< 0.001). However, no difference was observed between testes collected in February and April for 2-year-old stallions (Figure 4B).
Discussion
We report here that aromatase is expressed not only in Leydig cells of stallion testis but that immunoreactivity is also present to a lesser extent in the seminiferous tubules, in the cytoplasm surrounding germ cells. Sertoli cells therefore appear positive. It can be considered that both cell types could contribute differently to estrogen synthesis in the stallion. However, these results are different from those of Eisenhauer et al. (1994), Almadhidi et al. (1995), and Nagata et al. (1998), who reported an aromatase immunolocalization essentially in Leydig cells of stallion testes. Almadhidi et al. (1995) immunolocalized aromatase in Leydig cells of 3- to 4-year-old stallion testes collected in February and March, using a polyclonal antibody generated against equine testicular aromatase diluted 1:40. In this study we performed some modifications. First, another preparation of antibodies obtained from an immunoaffinity column was used (Séralini et al. in press) and diluted 1:1000. Second, before being incubated with primary antibody, sections were heated in a microwave oven for antigen retrieval. Application of this technique has been reported to significantly enhance antigen-antibody reactivity and therefore to increase detection sensitivity (Naganuma et al. 1990; Lan et al. 1995; Shi et al. 1997). Finally, the possible existence of nonrelevant endogenous peroxidase activity (pseudoperoxidase) in the tissue, which in turn can give a false-positive reaction when visualized with DAB, was abolished in this study by incubating the sections with 3% H2O2 in PBS after the dehydration step (Streefkerk 1972). With these treatments, high background and false-positive reactivity can be minimized and, most importantly, antigen detection can be optimized. Eisenhauer et al. (1994) and Nagata et al. (1998) immunolocalized aromatase in testes collected from postpubertal, adult, and aged stallions during breeding and non-breeding seasons. Using a standard avidin-biotin complex (ABC) staining, these authors reported that immunoreactivity was detected only in the Leydig cells. However, it should be noted that these researchers used a rabbit anti-human aromatase antibody rather than a specific antibody generated against equine aromatase, as in our study. A species specificity of aromatase, at least at the substrate level, has already been reported (Silberzahn et al. 1988; Gaillard 1991; Amri et al. 1993). Therefore, differences in immunohistochemical protocols and/or use of different antibodies may modify the detection sensitivity. For example, Turner et al. (2002) have shown, using a monoclonal antibody directed to a highly conserved region, that aromatase was present in interstitial tissue and seminiferous tubules of adult rat and marmoset testes. They reported, in agreement with our results, an intense labeling in Leydig cells but also significant immunostaining in the cytoplasm surrounding the heads of elongated spermatids.
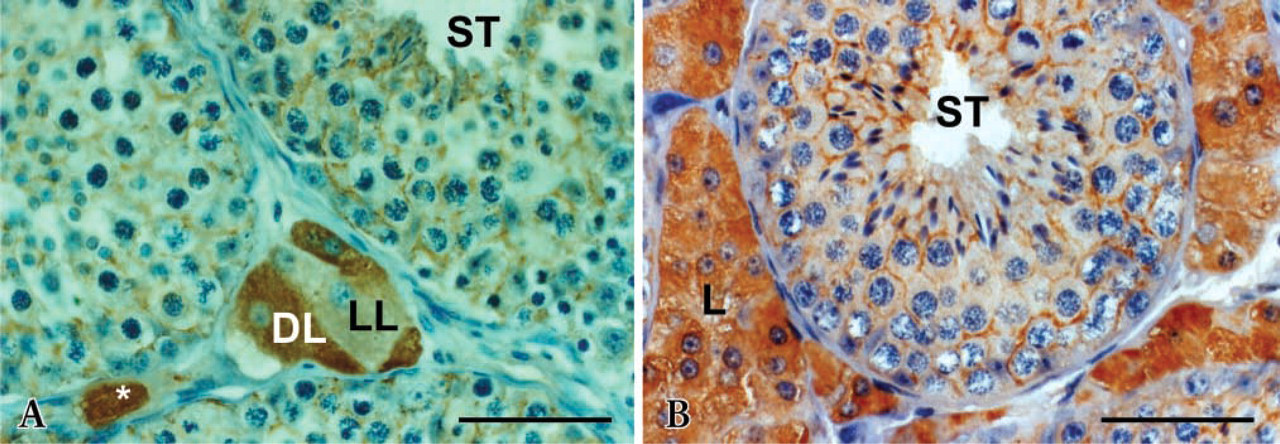
Age-related immunoreactivity in Leydig and Sertoli cells in stallion testes. (
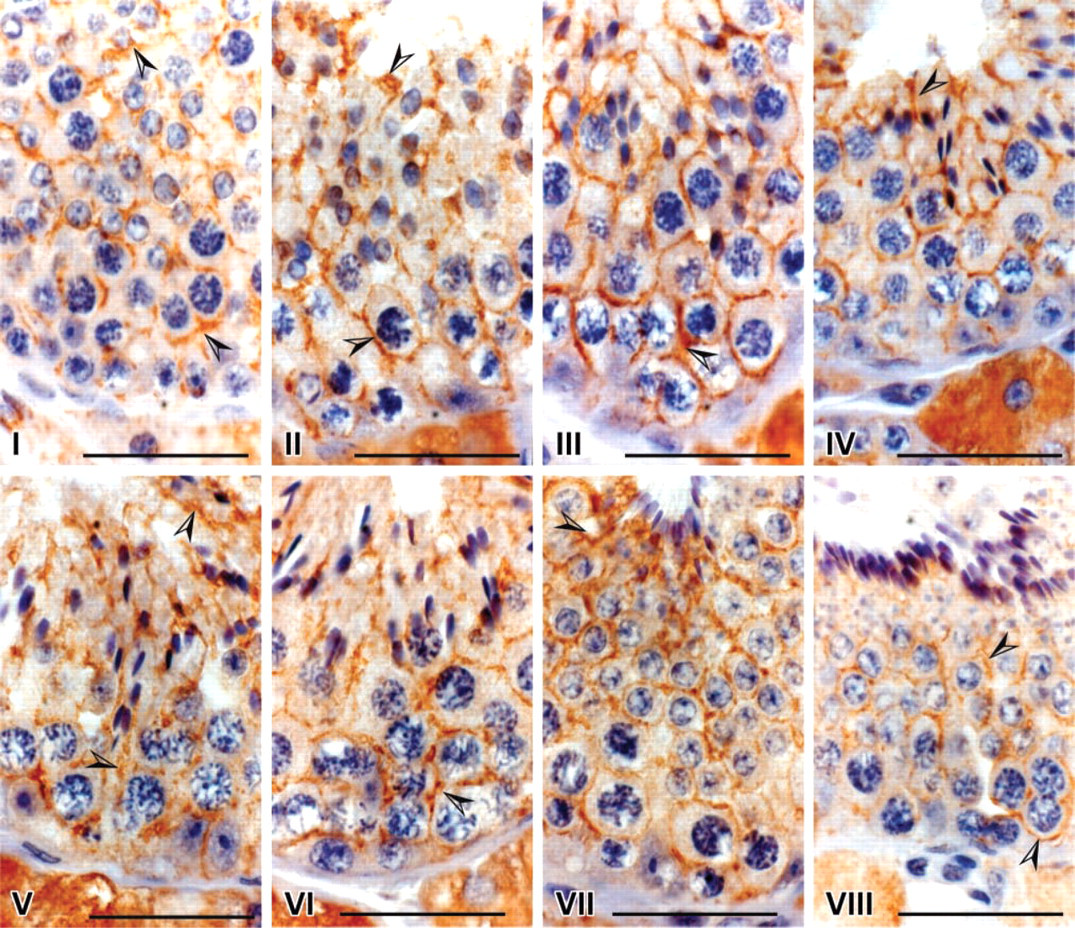
Spermatogenic stages-related immunoreactivity distribution in Sertoli cells of 3-year-old stallion. Although cytoplasmic immunoreactivity of Sertoli cells was observed in all spermatogenic stages (arrowheads), the pattern was stage-related. Immunoreactivity in stages III and VI is more intense in the basal than in the adluminal compartment of the tubule, in contrast to stages IV and VII. Immunoreactivity is uniformly distributed in stages I, II, V and VIII. No apparent immunostaining was seen in controls (data not shown). Bars = 5 μm.
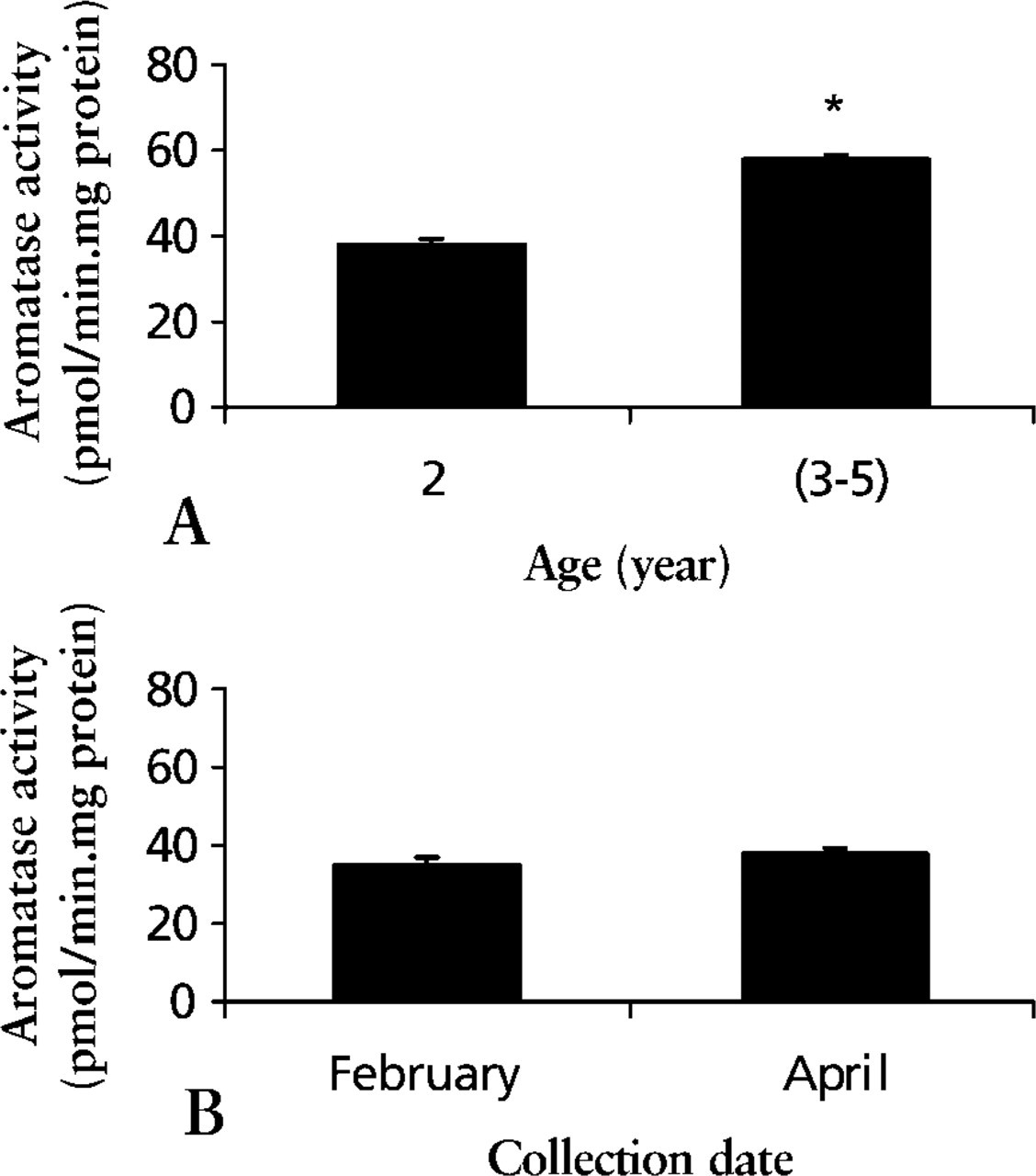
Testicular microsomal aromatase activity. (
Moreover, our study indicates that there are two types of Leydig cell populations in horse testes from 2-year-old postpubertal stallions that react differently with anti-equine aromatase. This is in agreement with results reported by Eisenhauer et al. (1994). Heterogeneity of Leydig cell populations, i.e., light and dark cell populations, was previously reported in stallion (Almahbobi et al. 1988) and in rat (Browne et al. 1990), indicating that there is a common feature in the mammalian testis that is likely related to different steroidogenic capacities of Leydig cells, as suggested by Maillard et al. (1994).
In the present study, the immunoreactivity of aromatase in Leydig cells was obviously very much stronger than that in Sertoli cells, indicating that Leydig cells are still the main sites of estrogen synthesis in stallion testes. This is a general phenomenon observed in most mammalian species (for review see Carreau et al. 2001) and in humans (Brodie et al. 2001). Moreover, aromatase in Sertoli cells may change with the season, as in the black bear (Tsubota et al. 1997). It increases from January to June and disappears in October. The black bear is known to be a true seasonal breeder. The onset of Sertoli cell aromatase in January is related to the initiation of testicular recrudescence. Therefore, the presence of aromatase and estrogen may play a role in re-initiation of spermatogenesis. However, our study failed to detect any important variation of aromatase immunostaining in Sertoli cells or around germ cells of 2-year stallion testes collected in winter (February) and spring (April to June). Horses do not hibernate and their testicular activity does not totally cease. It is likely that in stallion a Sertolian expression of aromatase is required during spermatogenesis, as previously suggested in other animals (Carreau et al. 2001; O'Donnell et al. 2001). Indeed, male mice lacking a functional CYP19 gene exhibit an increase in apoptotic germ cells, an arrest during spermiogenesis, leading to a decrease in round and elongated spermatids and finally to a reduction in fertility (Robertson et al. 1999). Moreover, long-term estrogen administration (70 consecutive days) restored spermatogenesis in the hypogonadal mouse (Ebling et al. 2000).
In the stallion the seminiferous epithelium cycle consists of eight stages with a helical pattern distribution in the tubule (Swierstra et al. 1974; Johnson et al. 1990). This allows observation of more than one spermatogenic stage in a transverse section. Our results indicate a differential distribution of aromatase in Sertoli cells and in the cytoplasm around germ cells related to the spermatogenic stages in seminiferous tubule in stallions. The variation always appears to be related to the spermatogonia and spermatid positions in the tubule, i.e., the basal and adluminal compartments of the Sertoli cells containing germ cells undergoing differentiation and meiosis (spermatogonia and spermatocytes) and cell differentiation (spermatids), respectively. Stage-dependent changes in Sertoli cells for steroidogenic enzyme activities and estrogen receptor (ER) distributions have been reported in the dogfish by Callard et al. (1985). They observed that microsomal aromatase activity was highest in regions of testicular lobule spermatocytes containing germ cells undergoing meiosis or early spermiogenesis, whereas ER were more concentrated in the regions containing spermatogonia. More recently, Betka and Callard (1998) showed that E2 could regulate spermatogenic progression by initiation of the DNA synthesis in testicular zones containing spermatogonia and premeiotic spermatocytes that could favor their differentiation. In the same way, E2 enhances the stimulatory effects of FSH on initiation of spermatogenesis in the rat (Kula et al. 2001).
In this work, we show microsomal aromatase activity in stallion testes increases with age. It is significantly higher in 3–5-year-old than in 2-year-old stallions. This is in agreement with results previously reported by our group concerning enzymatic activity, proteins, and aromatase mRNA in the horse testis (Lemazurier et al. 2001). In addition, aromatase activity evolves with the age-related changes of testicular morphology and steroid production, which is different from humans and rodents. Indeed, the peculiarities in horse are not only the increase in Leydig cell number from 2 to 20 years, but also in Sertoli cell number for up to 4 years, as previously reported by Johnson and Tatum (1989). Furthermore, whereas gonadotropin secretion increases around 17–20 months, testosterone concentration rises slowly from 17 to 20 months and then more rapidly to a steady state at 23 months (Naden et al. 1990).
In conclusion, it is likely that local aromatase production in Sertoli cells, which seems age- and stage-related in the stallion, would be necessary for spermatogenesis, particularly for spermatogonia undergoing proliferation and meiosis, and for maturation of spermatozoa or spermiogenesis.
Footnotes
Acknowledgements
Supported by Fonds Feder and CRAB (Comité de Recherche pour les Agrobioindustries, Calvados), and Conseil Général du Calvados. HS had a studentship from SFERE (Société Fran¸aise d'Exportation des Ressources Educatives).
We wish to thank Dr J. Fresnel for excellent assistance with microphotography.