
Abstract
The LIM kinase family includes two proteins: LIMK1 and LIMK2. These proteins have identical genomic structure and overall amino acid identity of 50%. Both proteins regulate actin polymerization via phosphorylation and inactivation of the actin depolymerizing factors ADF/cofilin. Although the function of endogenous LIMK1 is well established, little is known about the function of the endogenous LIMK2 protein. To understand the specific role of endogenous LIMK2 protein, we examined its expression in embryonic and adult mice using a rat monoclonal antibody, which recognizes specifically the PDZ domain of LIMK2 but not that of LIMK1. Immunoblotting and immunoprecipitation analyses of mouse tissues and human and mouse cell lines revealed widespread expression of the 75-kDa LIMK2 protein. Immunofluorescence analysis demonstrated that the cellular localization of LIMK2 is different from that of LIMK1. LIMK2 protein is found in the cytoplasm localized to punctae and is not enriched within focal adhesions like LIMK1. Immunohistochemical studies revealed that LIMK2 is widely expressed in embryonic and adult mouse tissues and that its expression pattern is similar to that of LIMK1 except in the testes. We have also demonstrated that endogenous LIMK1 and LIMK2 form heterodimers, and that LIMK2 does not always interact with the same proteins as LIMK1. (J Histochem Cytochem 54:487-501, 2006)
T
Several LIMK2 mRNA isoforms were identified and included LIMK2a, LIMK2b, and tLIMK2 (Ikebe et al. 1997,1998). Whereas LIMK2a represents the full-length transcript, LIMK2b is missing half of the first LIM domain and tLIMK2 is lacking both LIM domains and half of the PDZ domain. LIMK2a and 2b mRNAs are expressed in all tissues examined, whereas tLIMK2 expression is restricted to the testes (Ikebe et al. 1997, 1998). We have recently established the expression pattern and cellular localization of LIMK1 and found that it is widely expressed in all mouse tissues in both cytoplasm and nuclei. In cultured fibroblasts, LIMK1 is localized mainly to focal adhesions but is also found in the perinuclear region and sometimes in the nuclei. In primary cultures of embryonic day 18 (E18) rat hippocampal pyramidal neurons, LIMK1 is found in axons and growth-cone filopodia. In the soma of these neurons, LIMK1 is found in the perinuclear region identified as cis-Golgi (Foletta et al. 2004; Rosso et al. 2004). However, little is known about the expression pattern and cellular localization of the LIMK2 protein.
To date, published data suggest that the function of LIMK1 and LIMK2 is very similar because both proteins phosphorylate cofilin resulting in its inactivation, suggesting that the main difference between these two proteins may be their cell type-specific expression and a different subcellular localization.
Interestingly, there are differences between the phenotype of mice lacking the LIMK1 and LIMK2 genes. Although both mouse lines are healthy and fertile, LIMK1-deficient mice have abnormal spine morphology and synaptic regulation as well as deficient learning and memory (Meng et al. 2002). In contrast, mice devoid of the LIMK2 gene have only a mild phenotype in their testes. They have reduced-size testes, some of the tubules contain very few germ cells, and syncytial multinucleated cells are observed in their testes (Takahashi et al. 2002). Interestingly, mice lacking both LIMK1 and LIMK2 genes are also viable and fertile and exhibit a similar but slightly increased brain phenotype as the LIMK1-defecient mice (Meng et al. 2004).
However, important unanswered questions are whether the two proteins are expressed in distinct cell types and if the function of LIMK1 and LIMK2 is redundant in cells that express them both, or if the two proteins differ in their subcellular localization, which would imply some different functions for each. In the present study we have generated several specific rat anti-LIMK2 monoclonal antibodies (MAbs) that recognize specifically endogenous mouse and human LIMK2 proteins but not LIMK1. We have used these antibodies to study the expression pattern of LIMK2 in mouse tissues and human and mouse cell lines by immunohistochemistry, immunoblotting, and immunoprecipitation. We have also studied by immunofluorescence the cellular localization of LIMK2 in a variety of cell lines. These antibodies can be used to study the specific function of LIMK2 that distinguishes it from LIMK1 and answer the important question about the overlap between the functions of these two family members.
Materials and Methods
Expression Constructs and Protein Purification
Full-length mouse HA-tagged LIMK2 cDNA was obtained from Dr. Nakamura. LIMK2 cDNA corresponding to the PDZ domain (aa 145-260) was cloned by PCR into the BamH1 site of pGEX4T-1. The production of GST-PDZ protein and subsequent purification on glutathione-Sepharose column (Amersham Biosciences; Uppsala, Sweden) was performed according to standard protocols.
Tissue Culture and Transfections
Human cell line 293T, mouse cell lines NIH3T3, and monkey COS-7 were originally obtained from American Type Culture Collection (ATCC; Manassas, VA), whereas the origin of the mouse olfactory epithelial cell line, Olf 4.4.2, is described elsewhere (Bernard 1993). Cells were routinely grown in DME medium supplemented with 10% FCS. 293T cells were transfected with LIMK1 and LIMK2 constructs using the Fugene transfection reagent (Roche; Indianapolis, IN). Two days posttransfection, cells were harvested for analysis of overexpressed and endogenous LIMK proteins.
Immunization and Hybridoma Fusion
Rats were immunized by SC injections of 100 μg of purified protein as previously described (O'Reilly et al. 1998). Hybridomas were generated by fusing spleen cells from immunized rats with the SP2/0 myeloma cell line, as previously described (Galfre et al. 1977). Hybridoma-producing monoclonal antibodies (Mabs) to LIMK2 were identified and their isotype determined by a screening strategy previously described (O'Reilly et al. 1998). Briefly, 293T cells (2 × 106/10-cm dish) were transiently transfected with the vector encoding HA-tagged LIMK2 cDNA using Fugene transfection reagent (Roche). The following day the cells were fixed in 1% paraformaldehyde/PBS, permeabilized with 0.3% saponin (Sigma; St Louis, MO) and stained with hybridoma supernatants. Bound antibodies were revealed with fluorescein isothiocyanate-conjugated goat anti-rat IgG antibodies (1:100; Southern Biotechnology, Birmingham, AL) and detected by FACS scan analyzer (Becton, Dickinson Co.; Franklin Lakes, NJ). An anti-HA mouse MAb (1:1000; Covance, Richmond, CA) was used as positive control to detect the efficiency of LIMK2 transfection and to compare the profile of the antibody-producing clones. The mouse anti-rat IgG and Igκ MAbs used for isotyping were kindly provided by L. O'Reilly and A. Strasser and are described elsewhere (O'Reilly et al. 1998). LIMK2 antibody producing hybridoma clones were cloned three times and then adapted to growth in low serum medium. For production of large amounts of antibodies, hybridomas were cultured for several weeks in the miniPERM classic 12.5-kDa bioreactors (Vivascience; Hannover, Germany). Antibodies were purified on a protein G-Sepharose column (Amersham Biosciences).
Immunoblotting and Immunoprecipitation
Various mouse organs/tissues were harvested from C57Bl/6 mice, aged 12 weeks, and were frozen on dry ice before homogenizing in 10 vol lysis buffer per wet weight at 4C. Cell lysates were prepared from various cell lines or mouse tissues using cold lysis buffer [50 mM Tris, pH7.5, 150 mM NaCl, 0.1% Triton X-100, 10 mM dithiothreitol (DTT), 10 mM NaF, 1 mM Na3VO4, 1 mM sodium pyrophosphate, and one complete protease inhibitor tablet per 50 ml of buffer (Roche)]. The concentration of protein was determined using Bio-Rad protein assay reagent (cat. #500-0006; Bio-Rad Laboratories, Hercules, CA) and 100 μg of total protein from various tissues and cell lines were analyzed by immunoblotting or immunoprecipitation as previously described (Farlie et al. 1995). Immunoblotting was performed using 200 ng/ml of rat anti-LIMK2 MAb followed by 1:1000 dilution of horseradish peroxidase-conjugated anti-rat IgG (Chemicon International; Temecula, CA) and developed with ECL (Amersham), Supersignal (cat. #34,080; Pierce Biotechnology, Paddington QLD, Australia), or Supersignal extended duration substrate (cat. #34,075, Pierce Biotechnology) as the substrate using X-ray film to detect the chemiluminescent signal. As a control for protein loading, the membranes were stripped and re-probed with a mouse anti-tubulin MAb (1:1000 dilution, clone DM1A; Sigma) or HRP-conjugated polyclonal actin antibody (1:5000; Santa Cruz Biotechnology, Santa Cruz, CA). Coimmunoprecipitation analyses of cell lysates were performed essentially as previously described (Bernard et al. 1994) but using the lysis buffer for the wash buffer. One μg of anti-paxillin, -vinculin, -LIMK1, -LIMK2, or its isotype control (rat IgG2κ; Southern Biotechnology), MAbs were used for the immunoprecipitations. Immunoblotting was performed using the following dilutions: anti-vinculin mouse MAb (1:1000, clone VIN-11-5; Sigma); anti-paxillin mouse MAb (1:1000, clone 349; BD Transduction Labs); and 20 ng/ml anti-LIMK1 rat MAb (clone 1B11) and the anti-LIMK2 MAbs.
Immunofluorescence
Cells were grown to ∼50% confluence on coverslips pre-treated with fibronectin II and then fixed in 1% paraformaldehyde/PBS. Following blocking in 5% normal goat serum/PBS and 0.1% Tween-20, expression of LIMK1 was detected in the same blocking solution plus 10 μg/ml of rat anti-LIMK2 MAb (clone 10E5-19) following incubation for 1 hr at room temperature or overnight at-4C. A FITC-labeled goat anti-rat IgG antibody (1:100; Southern Biotechnology) was then used to reveal LIMK1 staining. Staining for vinculin and paxillin was performed by incubation with anti-vinculin mouse MAb (1:100 dilution) and anti-paxillin mouse MAb (1:100 dilution), respectively. Staining was revealed by incubation with Alexa 594 anti-mouse IgG (1:800 dilution; Molecular Probes). To stain for F-actin, Alexa 594 phalloidin (1:100; Molecular Probes) was added to the secondary antibody mix. Images were acquired using confocal microscopy (Leica Microsystems, Germany) or a Zeiss AxioVision 4.0 microscope (Zeiss, Germany). Visualization of cis-Golgi was carried out by staining with anti-GM130 MAb (1:100; BD Transduction Laboratories). GM130 is thought to be a structural element of the Golgi apparatus and is associated with the cis-compartment.
Controls for immunofluorescence staining included use of the IgG2 isotype control antibody (1:100), preincubation of the antibody with the immunizing peptide (100 μg peptide per 5 μg of antibody), use of secondary antibody alone, and for double-label immunofluorescence studies, use of each antibody separately to ensure that no fluorescence bleed-through from any one fluor occurred in the second channel. All of these methods demonstrated the specificity of the LIMK2 antibody, but only a few examples of the control staining are shown in the figures.
Immunohistochemistry
Paraffin-embedded, formalin-fixed tissues were sectioned and deparaffinized and gradually rehydrated. Antigens were retrieved by boiling the slides in 10 mM citrate buffer, pH, 6-10 min. Nonspecific staining was reduced by blocking with 10% FCS in PBS. Sections were incubated overnight at 4C with rat anti-LIMK2 MAb (10 μg/ml) or with an isotype-matched control antibody at the same concentration followed by washings with PBS/0.2% gelatin and addition of biotinylated goat anti-rat IgG (Vector Laboratories; Burlingame, CA). After incubation with 0.3% hydrogen peroxide in 10% methanol, the slides were washed and incubated with ABC complex (Vector) for 30 min followed by addition of diaminobenzidene (DAKO), counterstained with hematoxylin, and dehydrated in graded concentrations of alcohol and histolene before mounting in DPX (BDH; Poole, UK).
Results
Characterization of the Anti-LIMK2 MAbs
Rats were immunized with bacterially expressed recombinant mouse LIMK2 protein that contains the PDZ domain (amino acids 145-260) followed by fusion of the spleen cells with the myeloma cell line SP2/0. The hybridoma clones secreting the MAb against LIMK2 were identified by flow cytometric analysis of 293T cells overexpressing HA-tagged LIMK2 (see Materials and Methods). Hybridoma clones secreting MAb specific for LIMK2 yielded a profile similar to the one produced when anti-HA antibodies were used to detect the overexpressed HA-LIMK2 (Figure 1A, top left panel), which served as a positive control. The staining profile of the transfected 293T cells incubated with secondary antibody alone and non-transfected cells incubated with the anti-HA and secondary antibodies served as negative controls (Figure 1B). From an initial screen of 1000 hybridoma supernatants, 100 anti-LIMK2 MAb secreting hybridoma clones gave a profile similar to that shown in Figure 1A (positive control) and were selected for further analysis (Figure 1A). From these original clones, after further analysis by immunoblotting of non-transfected and HA-LIMK1-transfected 293T cells, we selected four clones as they specifically recognized endogenous mouse and human LIMK2. These clones, 10E5-19, 1G1-2, 1C6-1, and 1E5-7, were isotyped as IgG2a-specific MAbs (data not shown). These clones were grown and the supernatant isolated and purified using protein-G affinity columns for further use and analysis.
Specificity of the Anti-LIMK2 Antibodies
To determine whether the anti-LIMK2 antibodies are specific for LIMK2 and do not recognize LIMK1, 293T cells were transfected with HA-tagged LIMK1 or HA-tagged LIMK2, and the cell lysates were subjected to immunoblotting. The filters were probed with the anti-LIMK2 MAbs. HA-tagged LIMK2, but not HA-LIMK1, was detected with the four anti-LIMK2 antibodies (Figure 2A, top panel). To confirm that both proteins were equally expressed, the filters were stripped and reprobed with anti-HA antibodies (Figure 2A, bottom panel). As the 10E5-19 and 1C6-9-1 MAbs gave similar profiles in Western blots (Figure 2B), these antibodies were interchangeably used for all experiments.
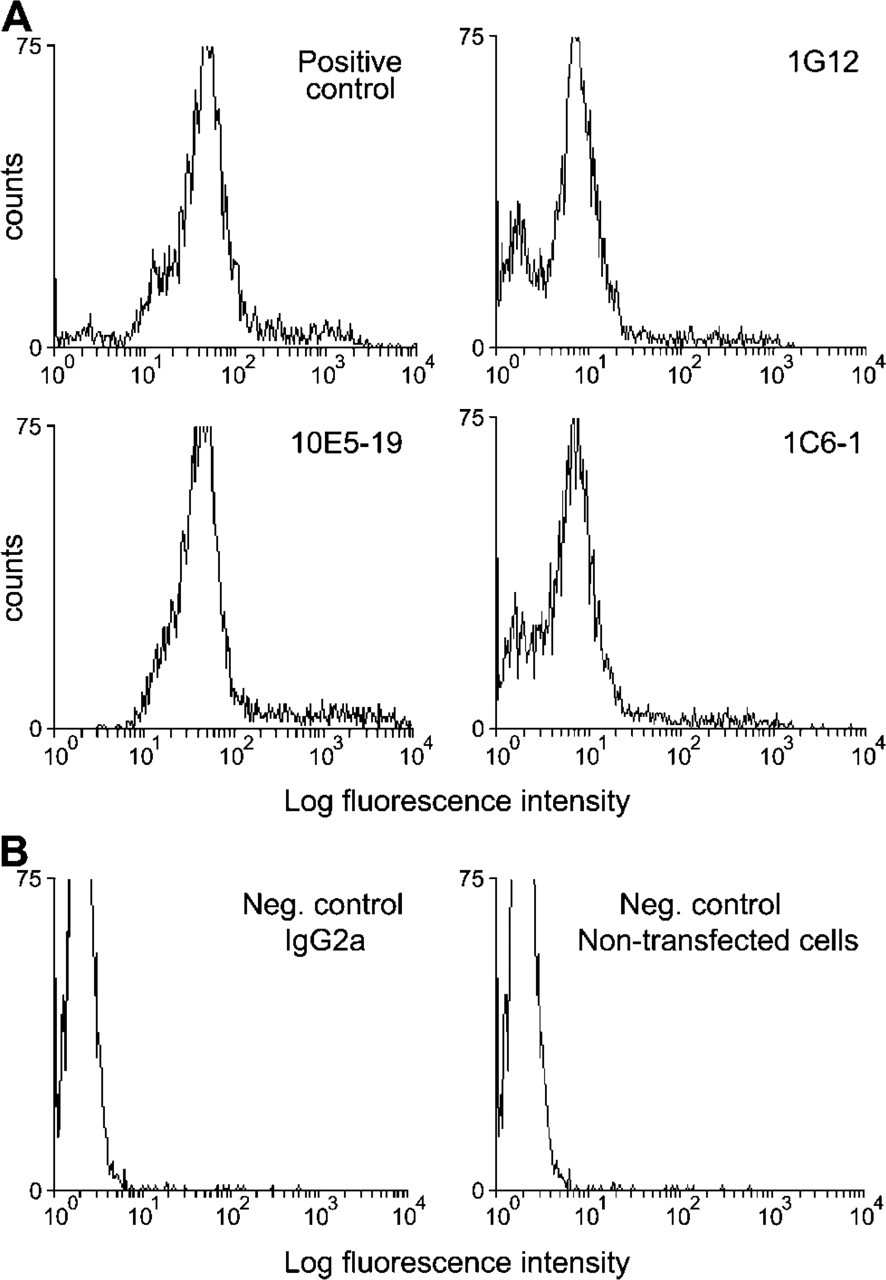
Determination of the specificity of the LIMK2 monoclonal antibodies (MAbs). Transiently transfected HA-LIMK2 293T cells were analyzed by FACS analysis and served as positive control. The level of fluorescence per 3000 viable cells for each sample was measured following fixation, permeabilization, and staining with an anti-HA antibody. (
Because there is little variation between the amino acid sequences of the PDZ domain among species, we performed immunoblot analysis of LIMK2 expression in mouse, monkey, and human cell lines. Western blot analysis of cell line extracts probed with the anti-LIMK2 MAb indicated that the antibody recognizes human (293T), monkey (COS), and mouse (3T3) LIMK2 (Figure 2C) in all cell lines examined with the exception of HeLa cells where LIMK2 expression was detected only upon prolonged exposure of the immunoblot (Figure 2C and data not shown).
Expression of LIMK2 in Mouse Tissues
Initially we used immunoblot analysis to study the expression pattern of LIMK2 protein in mouse tissues. However, because of irreproducible results we also studied LIMK2 expression by immunoprecipitation. We found that LIMK2, like LIMK1, is widely expressed in all of the tissues tested. High levels of LIMK2 were evident in uterus, thymus, and spleen whereas lower levels were observed in brain, heart, liver, stomach, skeletal muscle, and testes (Figure 2F). Very low levels were found in kidney, whereas very high levels of LIMK2 were present in intestines where expression could be easily detected by Western blotting (Figure 2G, left panels). A unique band of ∼75 kDa corresponding to LIMK2 protein was observed in all cell lines and mouse tissues. This band disappeared when the antibody was preincubated with the immunizing peptide (Figure 2G, right panels). Only the anti-LIMK2 MAb immunoprecipitated LIMK2 from mouse brain lysate, whereas the isotype control IgG2a failed to do so (Figure 2G, middle panels). A comparison between LIMK1 and LIMK2 proteins in COS-7 cell extract indicated that LIMK1 and LIMK2 have the same mobility on 10% SDS-PAGE whereas on a 4-20% gel, LIMK2 migrates a bit slower than LIMK1 (Figures 2D and 2E, respectively).
Cellular Localization of Endogenous LIMK2
We examined the subcellular localization of LIMK2 in various cells lines by immunofluorescence and observed its expression mainly in the cytoplasm. In NIH3T3, HeLa, and Olf cells, LIMK2 localized to panctae (Figures 3B and 3C, left panel; Figure 4A, right panel; Figures 4C and 4D, middle panel; and Figures 5A and 5B, left panel), a distinct localization from that of LIMK1, which localizes to actin filaments and to focal adhesions (Figures 3A and 3C, right panel; Figures 4A and 4B, middle panel; and Foletta et al. 2004). To confirm that LIMK2 is not expressed at the focal adhesions, costaining of LIMK2 and paxillin was performed and revealed that these proteins do not colocalize (Figure 5A). Specificity of the anti-LIMK2 antibody was confirmed by preincubation with the immunizing peptide that removed all specific staining in addition to using secondary antibodies alone (Figure 3, bottom panels). The pattern of LIMK2 staining in HeLa cells that expresses very low levels of LIMK2 is similar to that of 3T3 and Olf cells although the staining is much weaker, further confirming the specificity of the antibody (Figure 3C, left panel).
Additional evidence indicating that LIMK2 does not colocalize to focal adhesions comes from coimmuno-precipitation experiments with anti-LIMK2 and anti-LIMK1 MAbs. These experiments demonstrate that endogenous LIMK2 does not associate with either paxillin or vinculin, whereas LIMK1 associates with paxillin but not with vinculin (Figure 5D).
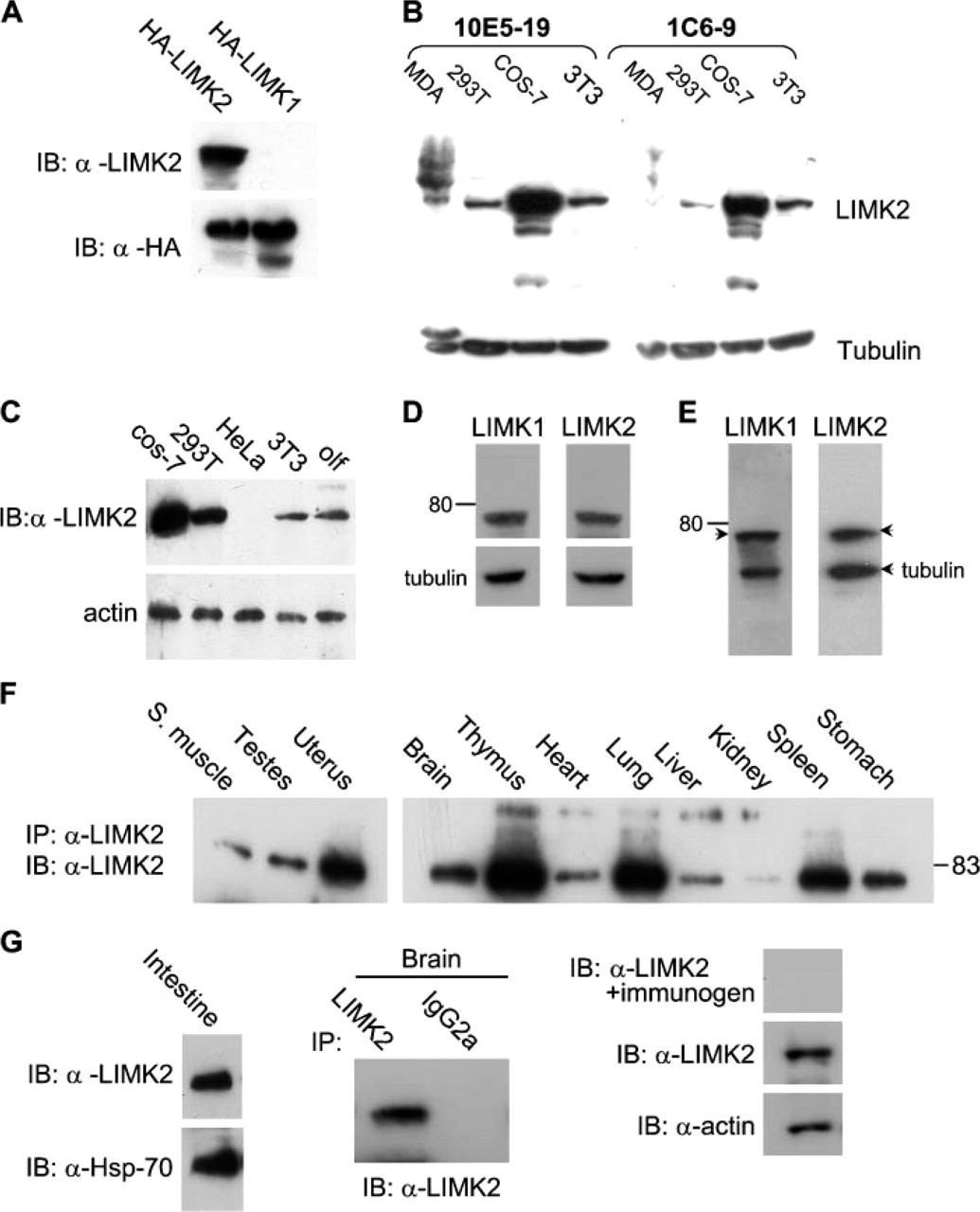
Expression analysis of over-expressed and endogenous LIMK2 proteins. (
LIMK2 Localizes in Part to the Golgi Apparatus
The punctae staining of LIMK2 protein suggests that it may associate with vesicles such as the Golgi apparatus. To further compare the localization of LIMK2 and LIMK1, NIH3T3 cells were costained for GM130, a protein known to be associated with the cis-compartment of the Golgi apparatus, together with anti-LIMK1 or-LIMK2 MAbs. Immunofluorescence staining showed that only some of the LIMK2 protein is associated with GM130 as seen in the merged images (Figures 4C and 4D) and thus with the Golgi apparatus particularly in the perinuclear region (Figure 5D). Although the subcellular localization of LIMK1 is distinct from that of LIMK2, it is also colocalized with GM130 in some neuronal cells as we, and others, have previously demonstrated (Foletta et al. 2004; Rosso et al. 2004).
Overexpressed LIMK2 Is Also Localized to Panctae
Previous studies on the cellular localization of over-expressed LIMK2 have demonstrated that it is very similar to that of the overexpressed LIMK1 protein. Furthermore, like the overexpression of LIMK1, LIMK2 overexpression was also reported to induce the formation of actin stress fibers and focal adhesions (Sumi et al. 1999,2001). In these experiments the level of the over-expressed LIMK2 protein was very high, suggesting that its high levels might have influenced the localization of the overexpressed protein. To address this possibility we transfected Olf cells with low amounts of HA-LIMK2 cDNA construct (2 μg) and studied its cellular localization by staining the transfected cells with anti-HA MAb.
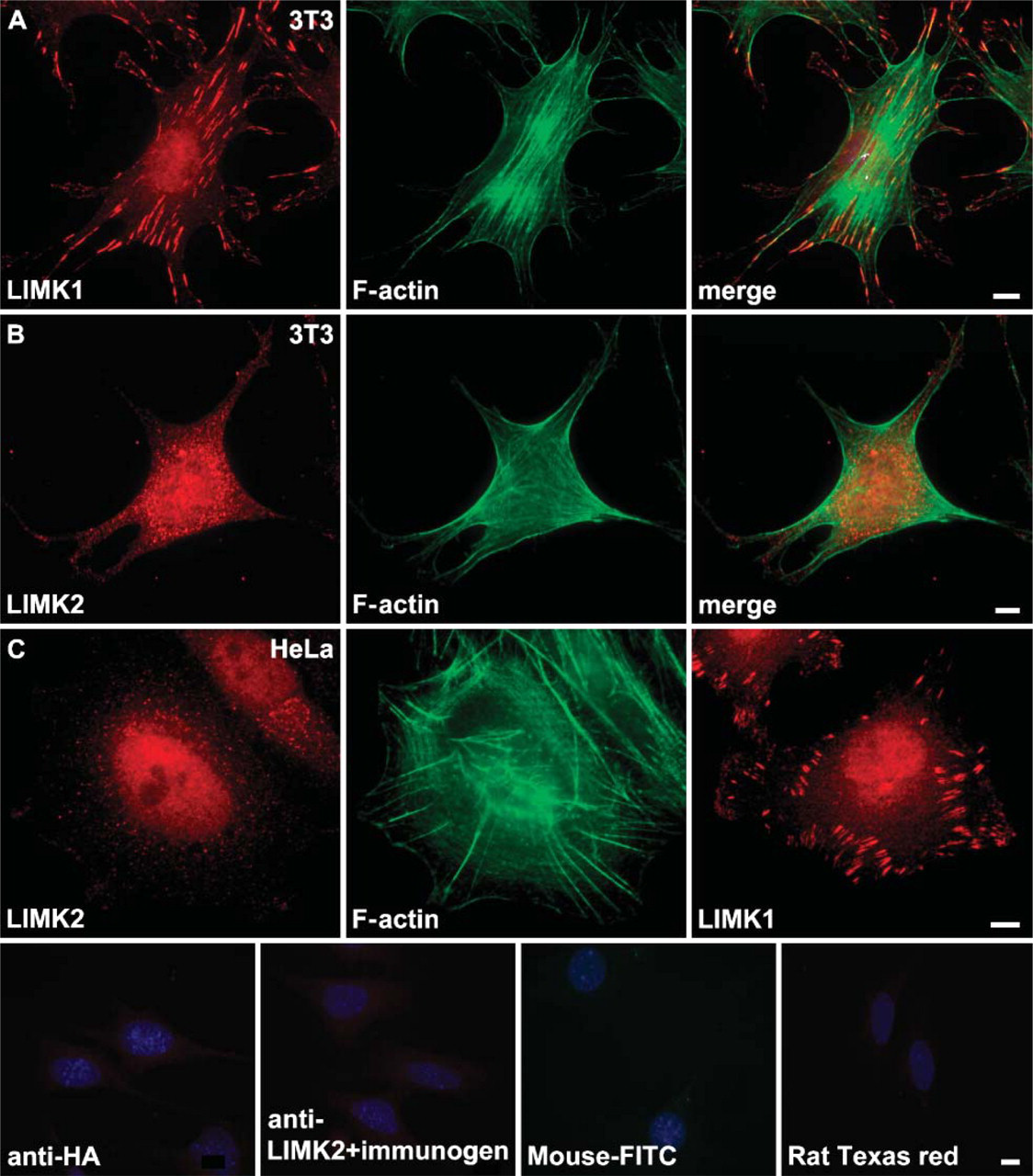
Cellular localization of endogenous LIMK2 is distinct from that of LIMK1. (
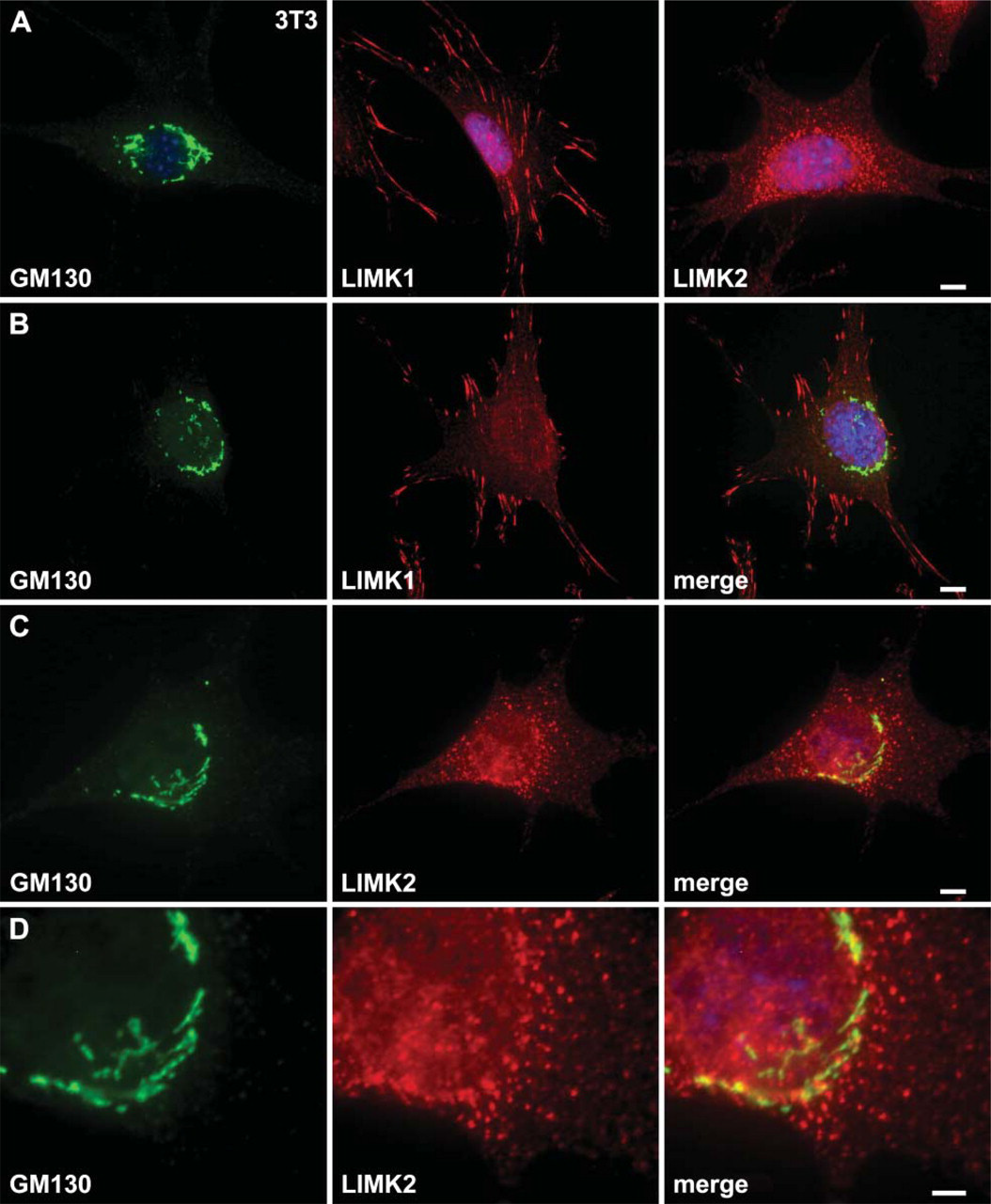
LIMK2 localizes in part to the Golgi apparatus in NIH3T3 cells. (
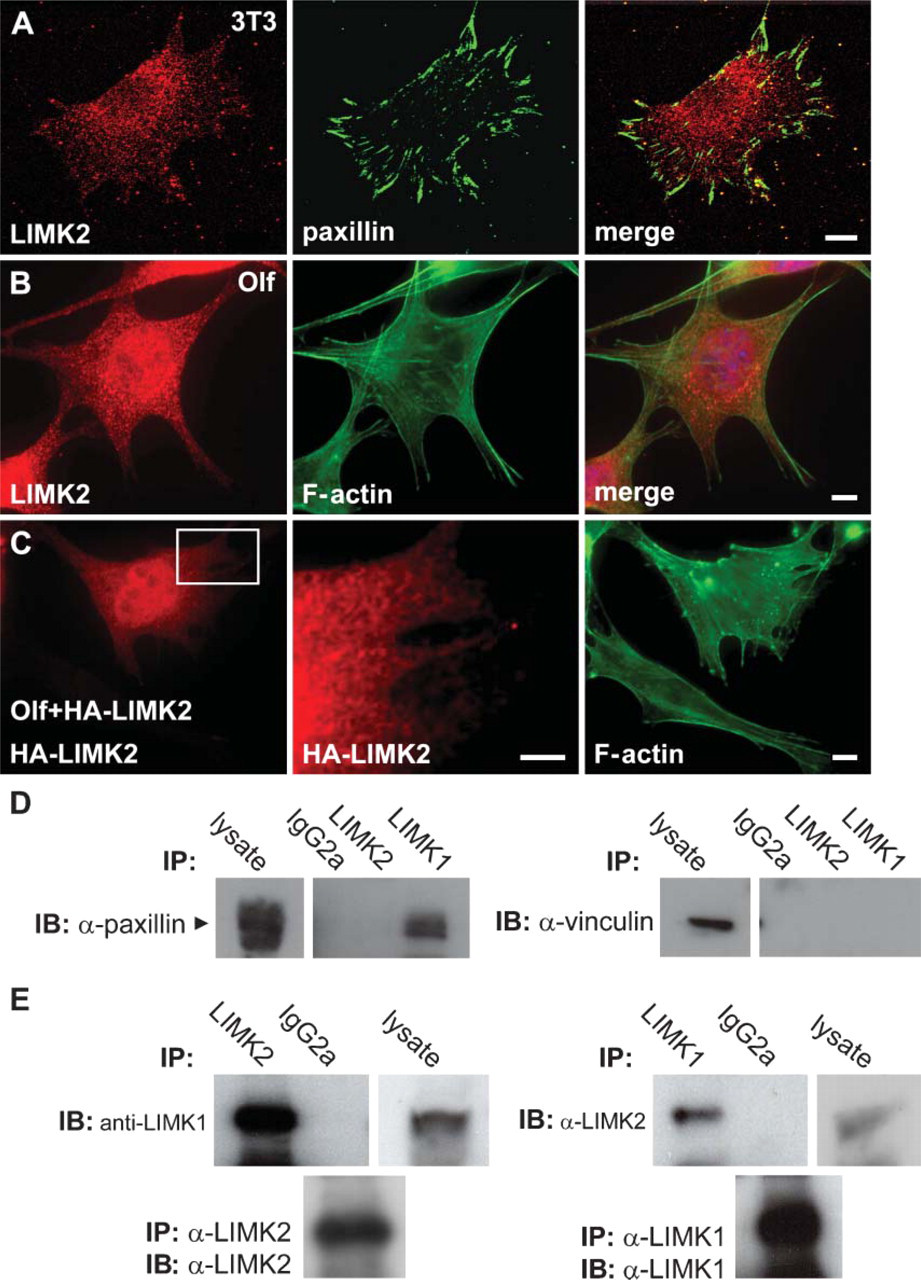
LIMK2 does not colocalize or form heterodimers with the focal adhesion protein paxillin but forms heterodimers with LIMK1. (
The cellular localization of the overexpressed LIMK2 protein was very similar to that of the endogenous LIMK2 protein, further indicating that LIMK2 is localized to panctae and not to F-actin and focal adhesion (Figure 5C).
Endogenous LIMK1 and LIMK2 Proteins Form Heterodimers
We have previously demonstrated that overexpressed LIMK1 and LIMK2 proteins can form heterodimers (data not shown). To test the possibility that such an interaction can also take place between the endogenous proteins, we immunoprecipitated LIMK2 from COS cell lysates with anti-LIMK2 MAb and after immunoblotting probed the filter with anti-LIMK2 MAb. LIMK2 was immunoprecipitated by the antibody together with LIMK1 (Figure 5E, left panels). We have also performed the reciprocal experiment where LIMK1 was immunoprecipitated and the filter was probed with anti-LIMK2 and also demonstrated interaction between LIMK1 and LIMK2, confirming that endogenous LIMK1 and LIMK2 can associate in vitro like the overexpressed proteins (Figure 5E).
Immunohistochemical Analysis of LIMK2 Expression in Mouse Tissues
The expression pattern of LIMK2 in mouse tissues was further examined by immunohistochemical staining. As the antibody did not recognize LIMK2 initially in paraffin-embedded tissues, an antigen retrieval technique was used to expose the LIMK2 epitope (see Materials and Methods). For negative controls, tissues were stained with an isotype-matched rat IgG2a MAb and antibodies preabsorbed with the immunogen.
Our immunoblot analysis revealed LIMK2 expression in all embryonic and adult mouse tissues examined, albeit at different levels according to tissue type. Immunohistochemistry staining confirmed these findings. Like LIMK1, LIMK2 staining was found in all tissues examined, and it was evident mainly in the cytoplasm and in some tissues also in the nucleus (Figures 6–8; Table 1). No nonspecific staining was observed in all tissues examined (Figures 6B, 6D, 6H, and 6K and Figures 7C′-7H′).
The Developing and Adult Nervous System
To study tissue-specific expression during development, immunohistochemical analysis of whole embryos was performed. In the developing central nervous system (CNS), LIMK2, like LIMK1, is expressed in postmitotic neurons of E14 mouse embryos (Figure 6A). In the olfactory bulb and spinal cord, abundant immunoreactivity of LIMK2 is observed. Lower level of LIMK2 immunoreactivity is observed in the developing mid-brain, cerebellum, hindbrain, and hypothalamus.
In the adult CNS, high levels of LIMK2 protein are found in the neurons of the hippocampus and the cerebral cortex as well as in Purkinje cells of the cerebellum (Figures 6C and 6E-G). In addition, high levels of LIMK2 are also found in the choroid plexus and the ependymal layer of the mature brain (Figure 6I). No staining was found in glial cells. Although LIMK2 expression in cell lines was found mainly in the cytoplasm, neurons express this protein in both the cytoplasm and nuclei.
Retina
LIMK2 staining was evident in all the layers of the retina including the photoreceptors, outer nuclear layer, inner nuclear layer, inner plexiform layer, and ganglion cell layer. Highest expression is seen in the in the horizontal bipolar and amacrine cells of the inner nuclear layer and the ganglion cells of the ganglion cell layer (Figure 6J).
Respiratory Tract
LIMK2 expression in the lung is very high and is found in all cells including epithelial cells lining the lumen of the respiratory bronchioles and the alveoli of the lung. Immunostaining was also found in the non-ciliated Clara cells and the terminally differentiated ciliated cells as well as in the epithelial cells lining the arterioles (Figure 7A).
Heart
High levels of LIMK2 are found in the myocardium of the heart (Figure 7B).
Digestive System
In the intestine, strong LIMK2 expression is detected in the crypts of Lieberkühn located at the base of the villi, the smooth muscle cells surrounding the small intestine and the goblet cells (Figure 7C). The chief and parietal cells of the stomach express high levels of LIMK2, whereas moderate levels are evident in the mucous neck cells (Figure 7D). In the liver, staining is observed in all cells including parenchymal cells that make up 80% of its cell population and the endothelial cells that line the sinusoids (Figure 7E).
Hematopoietic Tissues
Consistent with the immunoblot data, strong LIMK2 immunostaining was detected in the thymus, including cortical thymocytes that contain the more immature T-cells as well as the more mature T-cells present in the medulla. In the medulla, the epithelial reticular cells also stain for LIMK2 (Figure 7F). Similarly, as the immunoblot indicated, LIMK2 immunostaining in the spleen was strong in both the white and red pulp (Figure 7G).
Skin
LIMK2 expression is high in most of the cells in the skin including the epidermis, dermis, hair follicles, bulb cells, and sebaceous glands (Figure 7H).
Kidney
Although Western blot indicated low LIMK2 expression in the kidney, immunostaining revealed moderate expression in most of the cells. LIMK2 expression was found in all tubular structures including proximal and distal convoluted tubules as well as in the endothelial cells lining the arterioles and the venules. No staining was found in the glomeruli (Figure 7I).
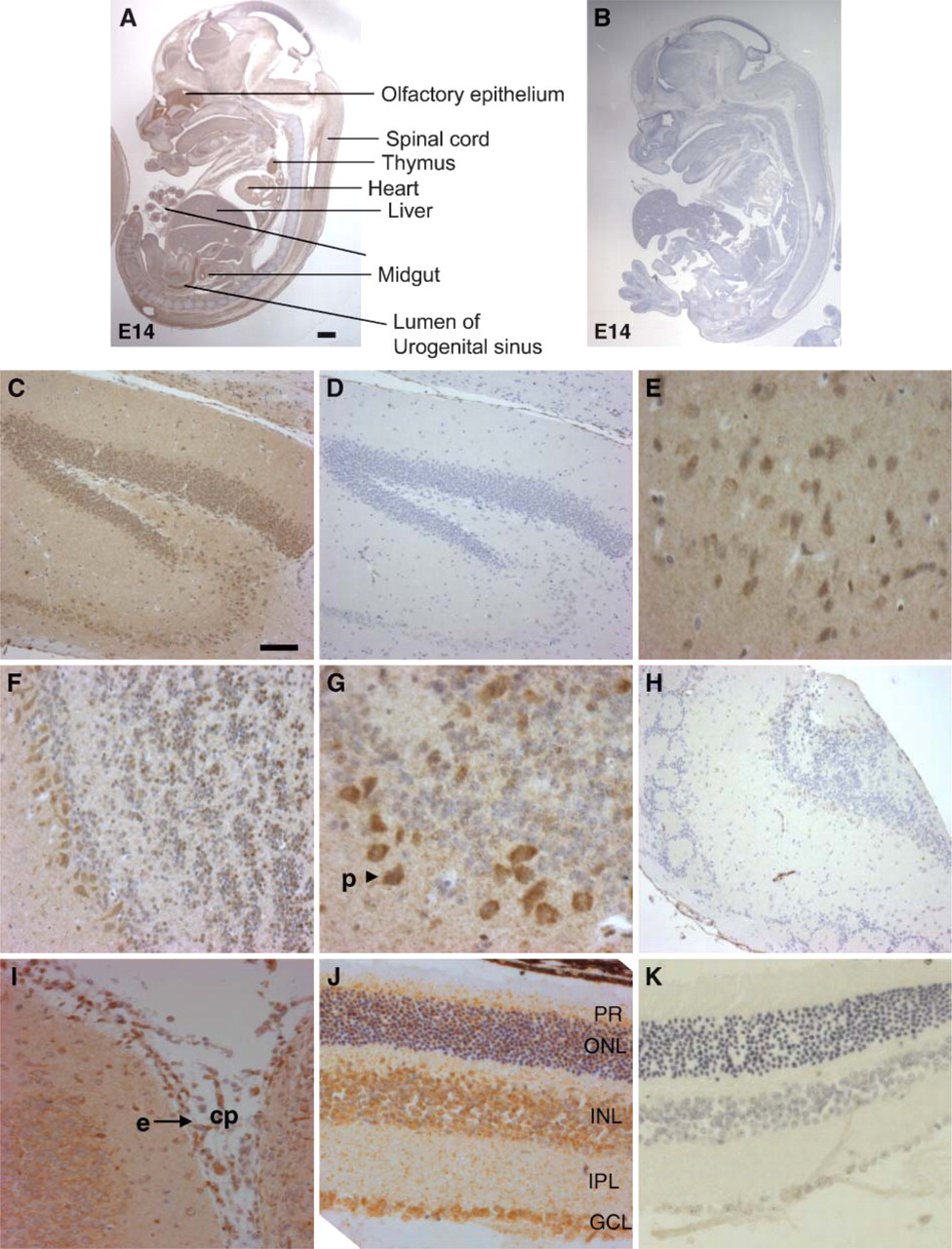
Photomicrograph of LIMK2 immunohistochemical staining of E14 mouse embryo, adult mouse brain, and retina. (
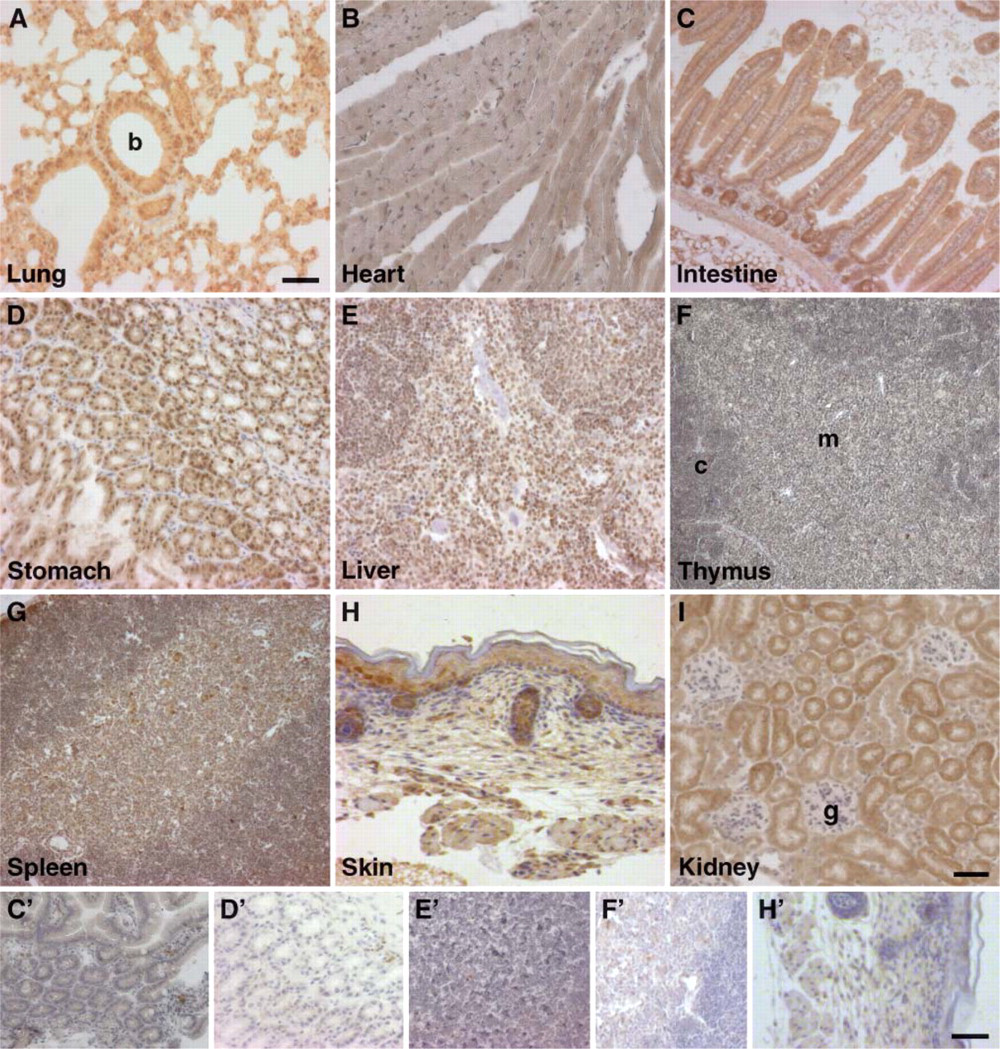
Immunohistochemical staining of LIMK2 in sections from a variety of mouse tissues as indicated. Tissues at the bottom panels were stained with isotype control antibody and correspond to LIMK2-stained tissues marked with uppercase letters. b, bronchiole; c, cortex; m, medulla; g, glomerulus. Magnification:
Reproductive Organs
In the uterus, a very high level of LIMK2 is found in the endometrial cells lining the uterine glands and the lumen of the uterus. In contrast, very low levels were observed in the myometrium (Figure 8A). In the ovaries, a high level of LIMK1 was found in the corpus luteum and folliculus, whereas moderate staining was observed in the epithelial layer (Figure 8B).
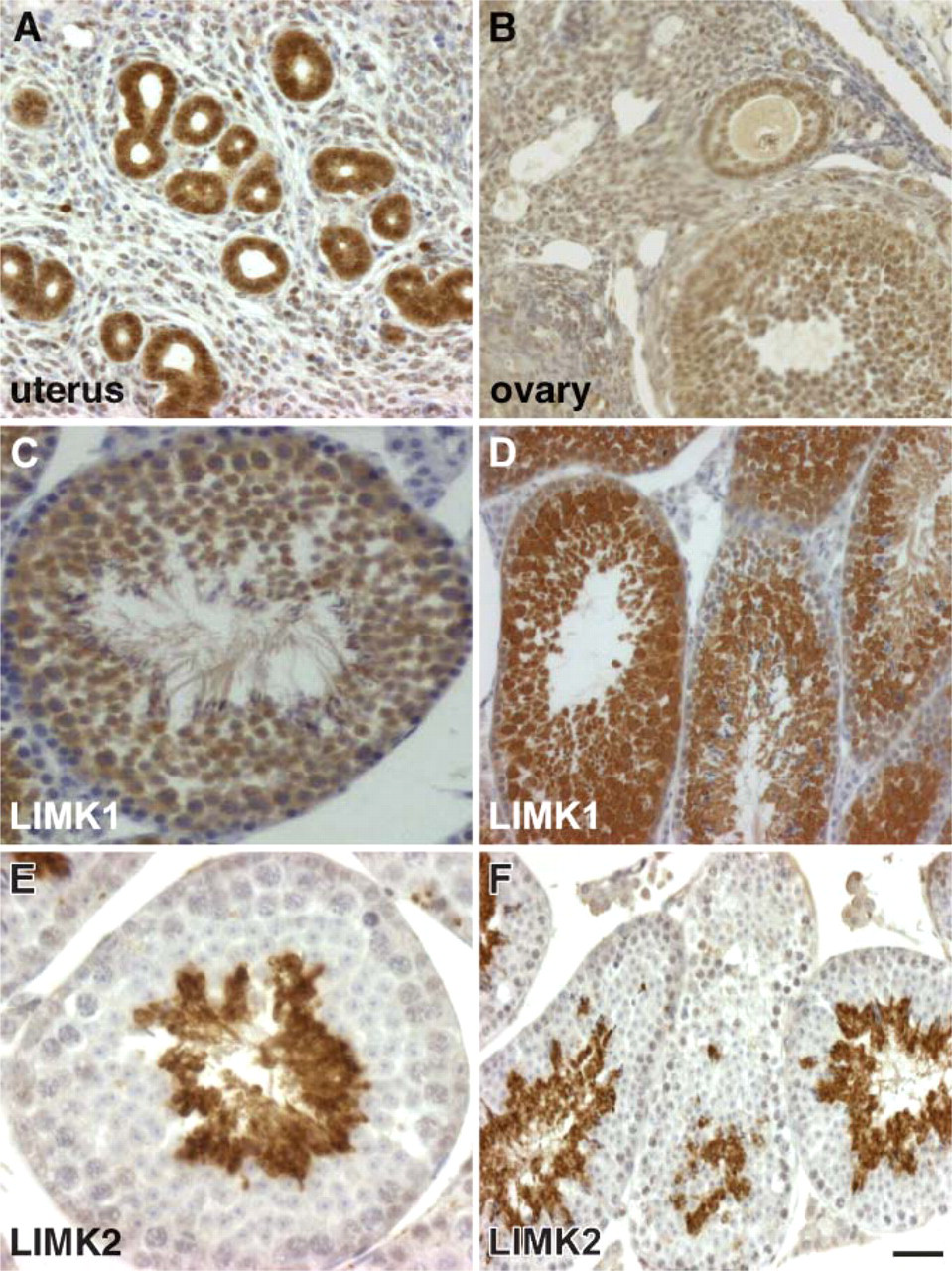
LIMK2 expression in the reproductive organs and comparison of LIMK1 and LIMK2 staining in testes. (
The most striking difference between the expression pattern of LIMK2 and LIMK1 is found in the testes where LIMK2 expression is complementary to that of LIMK1 (Figures 8C-8F). The very high LIMK2 expression is evident exclusively in the elongated spermatids (Figures 8E and 8F). In contrast, LIMK1 expression is found in the spermatocytes, Sertoli cells, and early spermatids in both the nuclei and cytoplasm, with lower levels observed in the mature spermatids and spermatogonia (Figures 8C and 8D).
Discussion
LIMK1 and LIMK2 were discovered more than a decade ago, but the precise expression pattern and cellular localization of LIMK1 has only recently been elucidated, whereas that of LIMK2 remained unknown.
As LIMK1 and LIMK2 share 50% amino acid identity, and the overexpressed proteins have very similar functions as they both phosphorylate the actin-binding proteins ADF and cofilin, it is therefore of great importance to determine their expression pattern in tissues to delineate the contribution of each protein to the regulation of actin dynamics. Because both proteins have similar activity in vitro, their different cellular functions may lie in their expression pattern or their cellular localization.
We have previously studied the expression pattern and cellular localization of LIMK1 using our rat anti-LIMK1 MAb and demonstrated that it is expressed in all mouse tissues examined (Foletta et al. 2004). Immunofluorescence studies demonstrated that LIMK1 is found mostly at focal adhesions in the cytoplasm and occasionally in the nucleus (Foletta et al. 2004). The generation of rat anti-LIMK2 MAbs has permitted an extensive examination of LIMK2 expression using multiple techniques. These MAbs were raised against the PDZ domain of mouse LIMK2 and recognized specifically both mouse and human LIMK2 but not LIMK1. The LIMK2 protein has an electrophoretic mobility corresponding to an apparent mass of ∼75 kDa, which is slightly higher than that of LIMK1 protein when separated on a 4-20% but not on a 10% SDS-PAGE. We believe that this is an anomaly of the electrophoretic behavior of the protein because the deduced number of amino acids is 638 in LIMK2 and 647 in LIMK1.
Localization and levels of LIMK2 expression in adult mouse tissues

No detectable expression;
, low;
, moderate;
, high expression.
Previous studies of the tissue distribution of LIMK2 relied entirely on the expression of mRNA by Northern blot (Koshimizu et al. 1997), RT-PCR (Ikebe et al. 1997), and in situ hybridization (Mori et al. 1997). The Northern blot and the PCR experiments identified the expression of LIMK2a and LIMK2b in all tissues examined (brain, thymus, lung, heart, stomach, spleen, kidney, liver, and small and large intestine). Similarly, the in situ hybridization experiments indicated a widespread expression throughout the developing rat embryos (Mori et al. 1997). Interestingly, comparison of the expression of LIMK1 and LIMK2 by in situ hybridization demonstrated that these two genes are coexpressed during rat development in most tissues except in the epithelium lining the digestive system where only LIMK2 expression was evident (Mori et al. 1997).
In agreement with previous studies we have found LIMK2 expression in all tissues examined. This tissue distribution is very similar to that of LIMK1 (Foletta et al. 2004). The only clear difference between the localization of these two proteins is in the testes where LIMK2 expression is complementary to that of LIMK1. Whereas LIMK1 protein is evident in most cell types in the testis including spermatogonia, Sertoli cells, and primary and secondary spermatocytes excluding Leydig cells or spermatids, LIMK2 is expressed exclusively in the elongated spermatids. It is possible that LIMK1 is required for the early stages of spermatogenesis. Alternatively, LIMK1 may play a specific role in Sertoli cells that provide structural and metabolic support to developing germ cells. These cells are also phagocytic and eliminate apoptotic germ cells during spermatogenesis (Lui et al. 2003). Indeed, it was reported that the RhoB/ROCK/LIMK1 pathway is required for the turnover of actin-based adherence junctions between Sertoli and germ cells, allowing the progressive movement of differentiating cells during spermatogenesis (Lui et al. 2003). On the other hand, LIMK2 may be required during spermatogenesis, as found in the studies of the LIMK2 knockout mice (Takahashi et al. 2002). Furthermore, like LIMK1, LIMK2 expression is also observed in the nuclei of a large number of cells including neurons and epithelial cells of the lung, stomach, kidney, uterus, and ovary. As it is now established that LIMK1 plays a role in the remodeling of tubulin (Gorovoy et al. 2005) and that it is involved in the regulation of Rho/Rock-mediated cyclin D1 expression (Roovers et al. 2003), it is not surprising to find that LIMK2 is also found in the nuclei of rapidly dividing epithelial cells. However, the role of LIMK2 in the nuclei of terminally differentiated neurons and in tubulin remodeling remains to be established.
One of the aims of this study was to differentiate between the roles of LIMK1 and LIMK2 by identifying differences in the expression patterns of these two proteins. Indeed, immunofluorescence analysis of cultured cells revealed that LIMK2 is found in the cytoplasm, but unlike LIMK1, which is found mostly at focal adhesions and around the nucleus of non-neuronal cells, LIMK2 is found throughout the cytoplasm in a punctae form. Furthermore, some LIMK2 is colocalized to the cis-Golgi apparatus and not to focal adhesions. Previous studies on the cellular localization of overexpressed LIMK2 demonstrated that it is found at focal adhesion, similar to the localization of overexpressed LIMK1 (Sumi et al. 1999). In this study we transfected cells with low amounts of LIMK2 cDNA and found that the overexpressed protein localized to panctae in a similar pattern to that of the endogenous protein, suggesting that the localization of the overexpressed LIMK2 in focal adhesions is the result of its overexpression. Finally, the difference in the cellular localization of the two LIMK proteins may indicate that they have different cellular functions and that they may be involved in different signal transduction pathways.
Mice devoid of LIMK2 gene have no overt phenotype except a mild impairment in spermatogenesis. In spite of this defect these mice are fertile (Takahashi et al. 2002). Moreover, mice devoid of both LIMK1 and LIMK2 genes are also fertile and have a slightly enhanced defect in brain function than the LIMK1 knockout mice. Considering the important role of LIMK1 in the regulation of actin dynamics, these findings may suggest that other proteins such as TESK 1 and 2, which can also phosphorylate cofilin, could compensate for LIMK function. Another possibility is that the LIMK knockout mice still express low levels of these proteins not detectable by immunoblotting. Interestingly, deletion in the LIMK1 and LIMK2 genes in these mice eliminates only the second LIM domain and half of the PDZ domains, raising the possibility that a protein with one LIM domain, half a PDZ domain, and a complete kinase domain is expressed in these mice.
Recently it was demonstrated that LIMK2 might be regulated differently than LIMK1. Although TGF-β can mediate actin cytoskeleton reorganization via LIMK2 and cofilin phosphorylation, LIMK1 is not involved in this process (Vardouli et al. 2005). Furthermore, platelet-derived growth factor and IL-1β can upregulate LIMK2 and cofilin, but not LIMK1, in canine cultured pulmonary artery smooth muscle cells (Bongalon et al. 2004). These recent findings suggest that the regulation of the activity and/or level of these two proteins are independently regulated and raise the possibility that their different cellular localization plays a role in their response to different stimuli.
Footnotes
Acknowledgments
This work was supported by the National Health and Medical Research Council (Australia).
We thank the Walter and Eliza Hall Institute monoclonal facility for performing the fusion, cloning, and purification of the monoclonal antibodies, Steven Mihajlovic for histology, Dr. Nakamura for the HA-LIMK2 cDNA construct, and Dr. Jim Bamburg for critical reading of the manuscript.