
Abstract
Animal fatty acid synthase (FAS: EC 2.3.1.85) (Poncari 1981) is a multifunctional enzyme that catalyzes the synthesis of long-chain fatty acids by using acetyl-CoA as a primer, malonyl-CoA as a two-carbon donor, and NADPH as a reductant (Bressler and Wakil 1961; Wakil 1989). FAS synthesizes mainly palmitate, as shown in a human breast cancer cell line in which FAS synthesized fatty acids composed of 80% palmitate, 10% myristate, and 10% stearate (Kuhajda et al. 1994). The enzyme is a homodimer whose molecular weight has been reported to be 480 kD (Thompson et al. 1981) or 550 kD (Kuhajda et al. 1994) and the synthase subunit to be 260 kD (Kuhajda et al. 1994) or 270 kD (Amy et al. 1989; Jayakumar et al. 1995). Most information about FAS is derived from biochemical studies, and the FAS level has been described to be low in normal human tissues due to its downregulation by dietary lipids (McCarthy and Hardie 1984; Weiss et al. 1986). It has also been reported that FAS expression is controlled by hormones (insulin, glucagon, glucocorticoids, and thyroid hormone) and nutrients (glucose and fatty acids) (Sul and Wang 1998). Antibodies, such as OA-519, against haptoglobin-related protein epitopes that recognize FAS (Kuhajda et al. 1994) have been used mainly in tumor biology research (Kuhajda et al. 1989a,b; Shurbaji et al. 1991, 1992; Redston et al. 1992; Jensen et al. 1993, 1995; Epstein et al. 1995; Hardman et al. 1995; Pizer et al. 1995; Alo et al. 1996; Gansler et al. 1997; Rashid et al. 1997), which indicates that high expression of FAS in breast, ovarian, and prostate cancer is associated with a poor prognosis (Kuhajda et al. 1989b; Shurbaji et al. 1991, 1992; Epstein et al. 1995; Hardman et al. 1995; Jensen et al. 1995; Alo et al. 1996). However, this is not the case in colorectal carcinoma (Rashid et al. 1997). FAS has also been detected in endometrial glands and stromal cells from the proliferative to the early secretory phase, and their FAS expression is closely associated with cell proliferation and expression of progesterone or estrogen receptors (Escot et al. 1990; Pizer et al. 1997). After cessation of cell proliferation in the mid- to the late secretory phase, the endometrial tissues become FAS-negative. However, the precise cellular localization and/or distribution of FAS has not been elucidated in normal human tissues, including human fetal tissues that exhibit active cell proliferation. Because human FAS cDNA has been cloned and its deduced amino acid sequence determined (Jayakumar et al. 1995; Semenkovich et al. 1995), we recently developed a new FAS antibody using a FAS peptide.
Here we describe the cellular distribution of FAS in normal human tissues in fetuses, newborns, infants, and adults.
Materials and Methods
Tissues
Normal human tissues were sampled at autopsy from three newborns, one infant, and three adults. For the central nervous system, seven adult brains (four females and three males, 29-69 years old) were employed. All cases were free of hormone (insulin and/or corticosteroid) or transfusion treatment. Fetal tissues, placenta, and decidua from 6, 20, 25, and 36 weeks of gestation were obtained from spontaneous abortions or stillbirths. In addition, curetted endometrium, surgically removed gallbladder for treatment of cholecystolithiasis, and extirpated breast for treatment of mammary carcinoma were also employed. Tissues were fixed with 10% neutral formalin and paraffin-embedded. Fetal brains were not included because they had not been processed for routine histological examination.
Antibodies
A polyclonal antibody was produced by immunizing New Zealand White rabbits by SC injections of a synthetic peptide composed of 17 amino acids corresponding to residues 1283-1299 of human brain FAS, which was coupled with keyhole limpet hemocyanin (Difco; Detroit, MI). The 17 amino-acid peptide is the sequence of the 3′ terminus of the predicted human brain FAS gene product (Jayakumar et al. 1995), which does not contain the acyl carrier protein or the thioesterase active site, which is also in the 3′ terminus (Semenkovich et al. 1995). Antisera were fractionated through protein A Avide Gel (Bio Probe International; Tustin, CA) to obtain IgG and were specifically purified by antigen-bound activated thiol Avide Gel F affinity column chromatography (Bio Probe International). Polyclonal antibodies against anterior pituitary hormones (adrenocorticotropin, growth hormone, thyroid-stimulating hormone, luteinizing hormone, and prolactin) were purchased from DAKO (Kyoto, Japan) and polyclonal anti-apoprotein antibody from IBL (Fujioka; Gunma, Japan). Mouse monoclonal antibody against Ki-67 antigen (MIB-1) was from Immunotech SA (Marseille, France).
Western Blotting
Human gastric carcinoma cell line TKGW from IBL was employed because immunostaining of FAS by our antibody was positive in this cell line. A gastric cancer cell line, MKN-1, maintained in our laboratory and negative for FAS immunostaining, was also used. Both were homogenized in lysis buffer containing CHAPS and protease inhibitors (pefabloc, leupeptin, pepstatin, and aprotinin) and centrifuged at 15,000 × g for 30 min. The supernatant was mixed with sample buffer (18.5 ml (1 mol)/liter Tris-HCI, pH 7.8, 15 ml 2-mercaptoethanol, 30 ml glycerol, and distilled water, to 100 ml) in a ratio of 2:1. Protein concentration was determined by the Lowry method, and 10 μg protein was subjected to SDS-PAGE and then blotted onto a nitrocellulose membrane. The membrane was then incubated with 2 μg/ml anti-FAS antibody IgG or FAS antibody absorbed by 5 μg antigen, followed by anti-rabbit goat IgG coupled to horseradish peroxidase (IBL). The peroxidase reaction product was visualized with Konica Immunostain HRP1S-50B (Konica; Tokyo, Japan).
Immunohistochemistry
Immunohistochemical procedures were described previously (Hiraki et al. 1997). In short, 3-±-thick sections were treated with methanol-hydrogen peroxide for 20 min to block endogenous peroxidase activity and then with 5% skimmed milk (Yukijirushi; Sapporo, Japan) in PBS for 30 min to avoid nonspecific adsorption of IgG. The concentration of the primary FAS antibody was 2 μg IgG/ml. The antibodies against anterior pituitary hormones were prediluted and the MIB-1 was diluted to 1:100. For MIB-1, the sections were microwaved for 15 min to retrieve the antigen. All slides were incubated with the antibodies overnight at 4C, after which they were reacted with biotin-labeled anti-rabbit IgG (Nichirei kit; Tokyo, Japan) for the polyclonal antibodies or anti-mouse Igs (Nichirei kit) for the mouse monoclonal antibody, and then with streptavidin-conjugated peroxidase (Nichirei kit) for 20 min at room temperature. Peroxidase reaction product was visualized with the Graham-Karnovsky solution (Graham and Karnovsky 1966) containing 65 mg of NaN3/dl to block endogenous peroxidase activity. Nuclei were counterstained with 1% methyl green.
In breast cancer tissues known to be FAS-positive (Thompson et al. 1981; Kuhajda et al. 1989b; Chalbos et al. 1990; Joyeaux et al. 1990; Shurbaji et al. 1991; Jensen et al. 1995; Alo et al. 1996; Milgraum et al. 1997), the specificity of FAS immunostaining was confirmed by omission of the primary antibody and use of preimmune rabbit IgG (2 μg/ml) or primary antibody absorbed with 5 μg antigen for the first step of the immunostaining procedure.
Results
Specificity of the FAS Antibody and Validity of Immunostaining
The antibody purified by affinity chromatography reacted with a 270-kD protein in the extracts of liver and TKGW cells but did not react with the extract of MKN-1 by Western blotting, as shown in Figure 1A. The 270-kD protein has the same molecular weight as the monomer of human brain FAS (Jayakumar et al. 1995), which is believed to be formed through disruption of disulfide bonds in the dimer (Stoops and Wakil 1982) by 2-mercaptoethanol in the sample buffer. Absorbed antibody or normal rabbit IgG did not reveal any bands in the TKGW cell extract by Western blotting (Figure 1A).
As shown below, the conditions of immunostaining with the FAS antibody were established by immunohistochemistry with human breast cancer tissue, which is known to be FAS-positive (Thompson et al. 1981; Kuhajda et al. 1989b; Chalbos et al. 1990; Joyeaux et al. 1990; Shurbaji et al. 1991; Jensen et al. 1995; Alo et al. 1996; Milgraum et al. 1997). The antibody clearly labeled breast carcinoma cells (Figure 1B), but the normal rabbit IgG (not shown) and antibody absorbed with the homologous antigen were negative (Figure 1C). FAS was distributed homogeneously in the cytoplasm and had a granular appearance in the intensely stained cells.
FAS-positive Cells in Adults
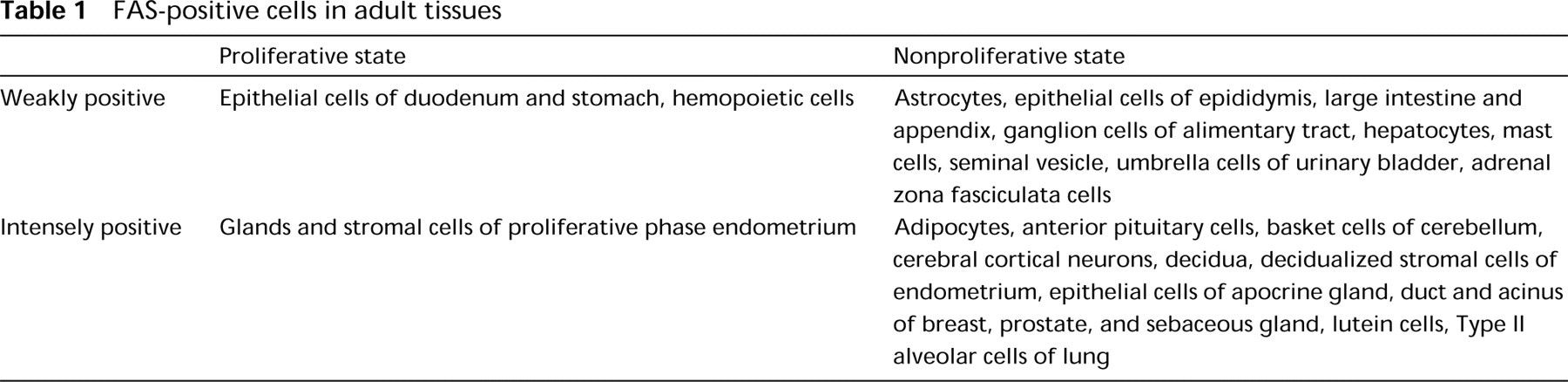
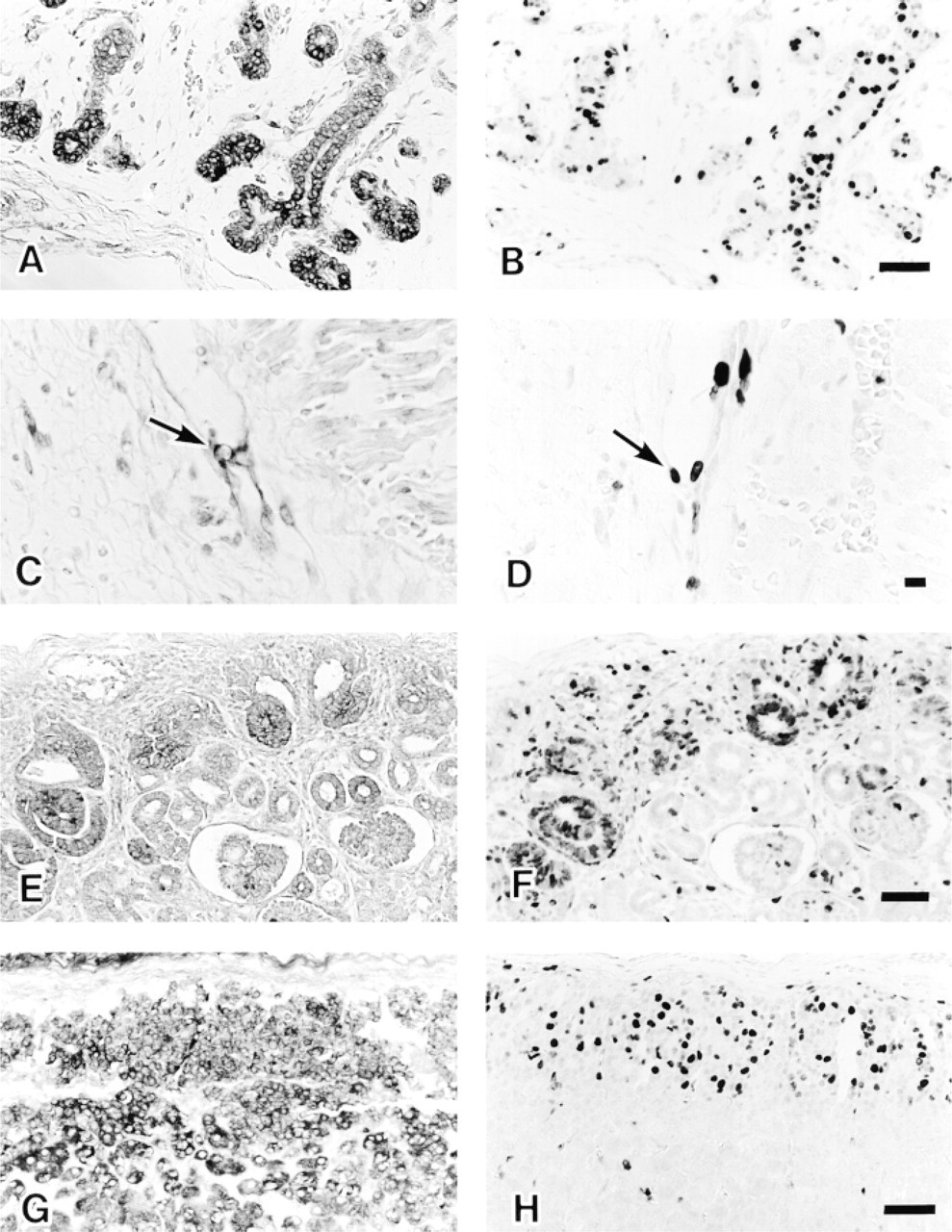
The results with human adult tissues are summarized in Table 1. Strongly FAS-positive cells were mainly nonproliferating, and limited amounts of FAS were detected in areas of active cell turnover, which consisted of cells in the proliferative zone (Ki-67-labeled) of gastroduodenal mucosa, hematopoietic cells in bone marrow, proliferating gland cells, and stromal cells of endometrium. Nonproliferating FAS-positive cells in the CNS were cerebral cortical neurons of the frontooccipital lobes, mainly in the third to fifth layers (Figure 2A), some neurons in the basal ganglia, hippocampus, paraventriculus, pons and medulla oblongata, basket cells of the cerebellum (Figure 2B), and a few astrocytes. Axons of basket cells that terminate on or attach to Purkinje cell bodies were also positive. Hippocampal basket cells were not labeled. There were no gender differences in the frequency of FAS-positive neurons in cerebral cortices. Cerebellar cortical basket cells showed similar positive staining in the seven brains examined, whereas cerebral cortical neurons exhibited mild fluctuation in the frequency of FAS staining from brain to brain. FAS-positive cells were also found in the digestive system [absorptive cells of the large intestine (Figure 2C) and the appendix vermiformis and hepatocytes in Zone 1 to Zone 2 (Figure 2D)], some Type II alveolar cells of lung (Figure 3A) also positive for surfactant apoprotein, in the urogenital system [a subset of epithelial cells of the epididymis and seminal vesicle, main prostate glandular cells (Figure 3B), mucosal and submucosal glands of the prostate, decidualized stromal cells of the endometrium, lutein cells of the ovary, and umbrella cells of the bladder], anterior pituitary cells (Figure 3C) (partly identical with GH and/or ACTH cells), subcutaneous and intra-abdominal fat cells, apocrine gland and sebaceous gland of skin (Figure 3D), tissue mast cells, adrenal zona fasciculata cells, and epithelial cells of ducts and acini of the breast.
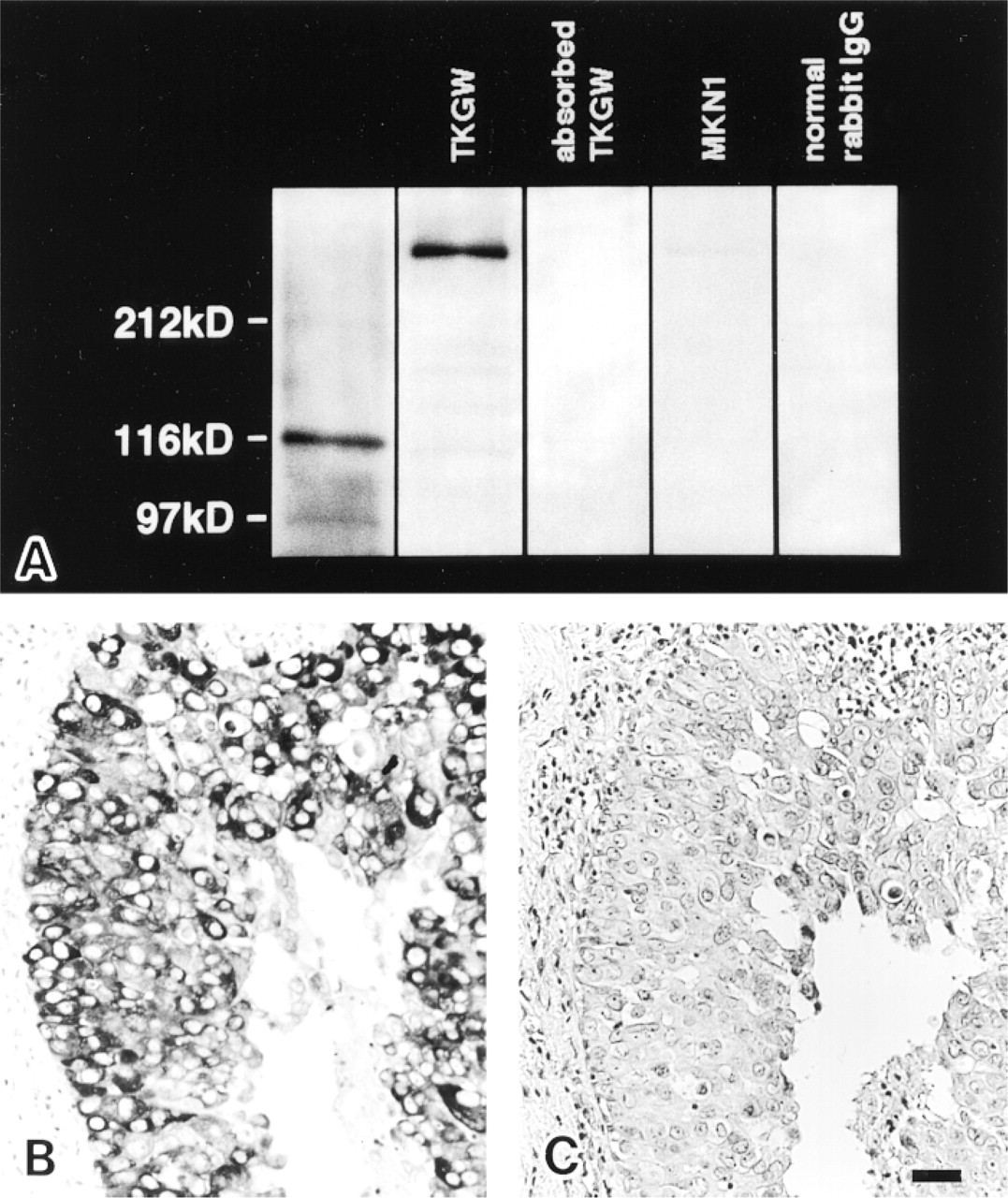
Western blotting (
FAS-positive Cells in Fetuses
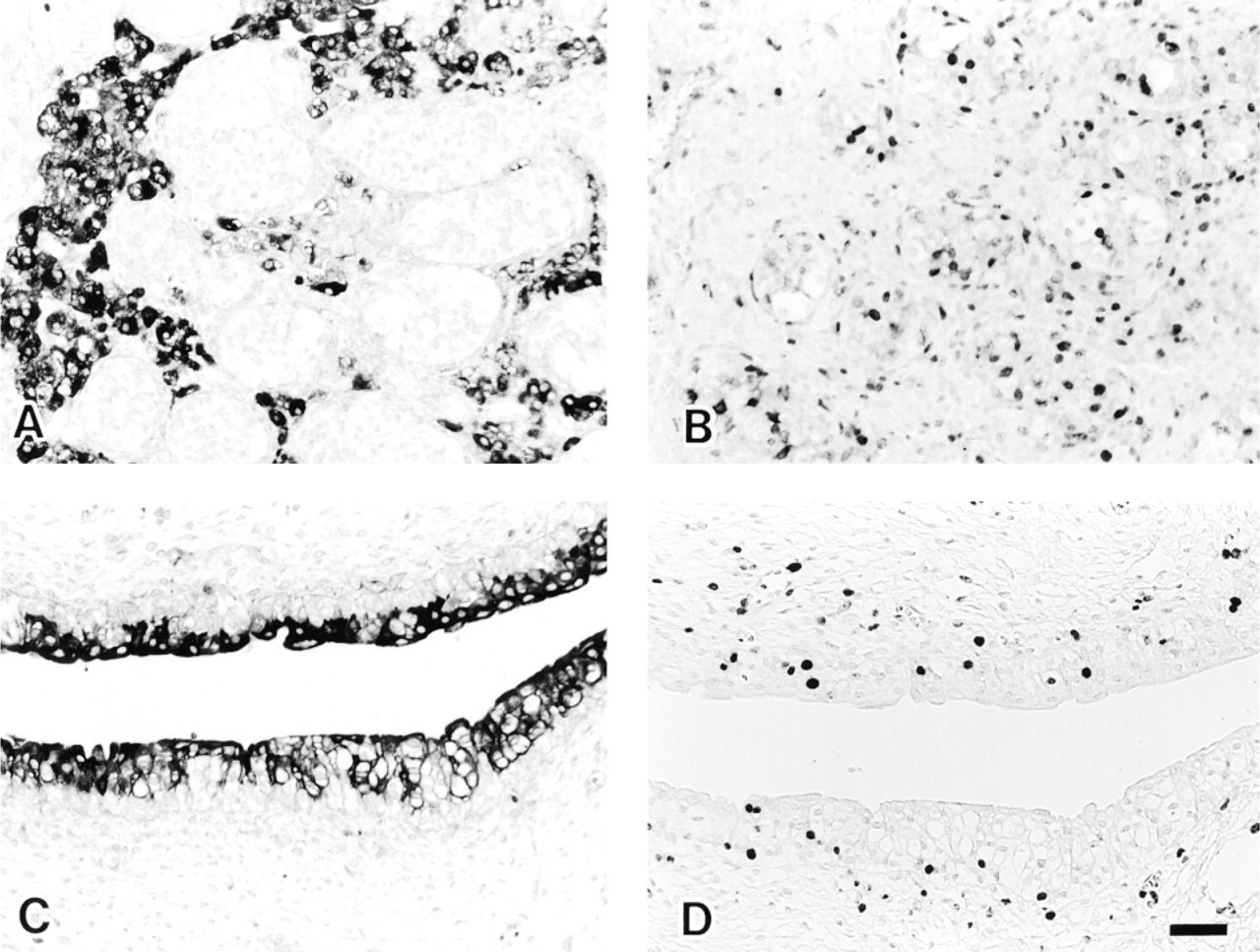
In the 6-week-old fetus, hepatocytes and hemopoietic cells, including erythroblasts, were positive. Because each visceral organ was easily identified, two 20-week-old fetuses were examined precisely except for their brains. The results are summarized in Table 2. The FAS-positive cells were predominantly those labeled with Ki-67 in areas of cell proliferation, although the Ki-67-labeled cells exceeded the number of FAS-positive cells. Positive cells in the digestive system were basal layer cells of squamous epithelium of tongue and esophagus, nonmucinous salivary gland cells (Figures 4A and 4B), glandular cells of gastrointestinal mucosa, pancreatic acinar cells, and hepatocytes. In the cardiopulmonary system, positive cells were found in the endocardium and tracheobronchial glands, bronchopulmonary epithelium, chondrocytes of tracheobronchial cartilage, and vascular endothelial cells (Figures 4C and 4D). Positive cells of the urogenital system were found in the S-shaped proximal tubules of kidney (Figures 4E and 4F) and in the prostate gland. In the endocrine system, anterior pituitary cells, thyroid follicular cells, and subcapsular adrenocortical cells were positive (Figures 4G and 4H). In addition, thymocytes and nodal lymphocytes in the periphery of the tissue were weakly positive. Schwann cells and mesenchymal cells of peripheral nerves in sympathetic ganglia and the Auerbach plexus of the gastrointestinal tract, a few neuroblasts in the adrenal medulla, fibroblasts (especially in the perivertebral region), striated muscle cells of tongue, and hematopoietic cells, including erythroblasts and megakaryocytes, showed cytoplasmic FAS, but the frequency of positive cells was low. However, cardiomyocytes and smooth muscle cells of visceral organs were hardly labeled despite being positive with Ki-67. Some Ki-67-negative cells were FAS-positive in the fetus, but this was limited and consisted of adipocytes, ganglion cells of sympathetic ganglia (proliferating Schwann cells were also positive) and the Auerbach plexus, Leidig cells of testis (proliferating seminiferous tubules were negative) (Figures 5A and 5B), sebaceous gland, urinary tract uroepithelial cells (Ki-67-labeled cells were negative) (Figures 5C and 5D), and the upper layer of adrenocortical cells (Figures 4G). Tissue mast cells and osteoblasts were also positive. Placenta from the sixth week of gestation or later showed FAS-positive cytotrophoblasts. Syncytiotrophoblasts and other components of the placenta were negative. Tissues of later gestational ages (25 and 36 weeks), newborns at 1 month, infants at 6 months, 10 months, and infants at 3 years had much more limited distributions of FAS-positive cells, almost similar to those of adult tissues. For example, a 1-month-old infant was FAS-negative in proximal tubules of kidney, subcapsular adrenocortical cells, Schwann cells of sympathetic ganglion, and mesenchymal cells (fibroblasts, chondrocytes, endothelial cells), which had ceased to proliferate.
FAS-positive cells in adult tissues
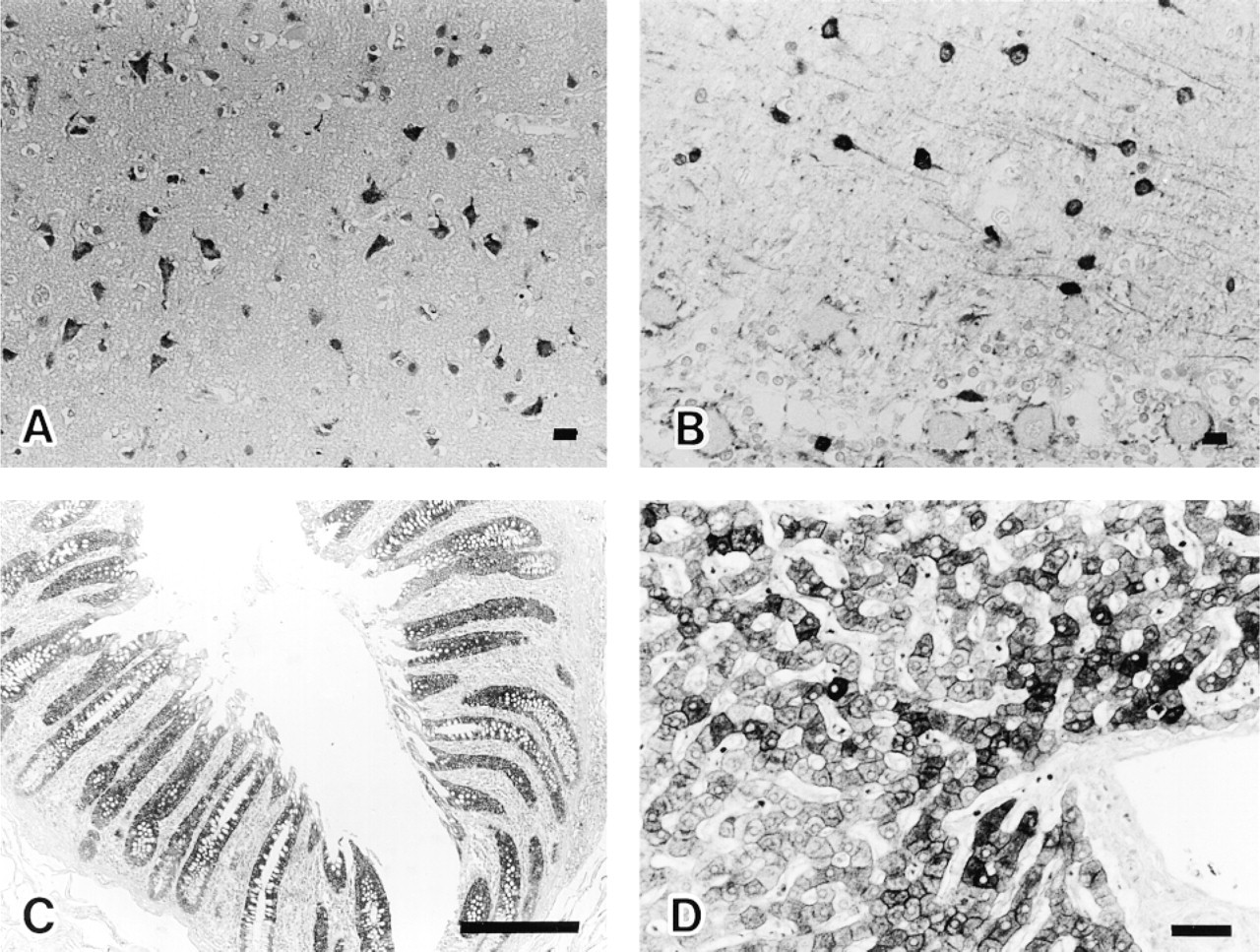
FAS-labeled cells in adult tissues. Cortical neurons mainly in the third to fifth layer show FAS-positivity (frontal lobe). Polygonal larger neurons and a few neuron processes are labeled (
Discussion
Long-chain fatty acids, such as palmitate, are essential constituents of membrane lipids and are also important substrates for cellular energy metabolism. FAS synthesizes mainly palmitate, as stated above. Expression of FAS mRNA in nonfetal human tissues was high in brain, liver, lung, intro-abdominal adipose tissue (Shurbaji et al. 1992; Jayakumar et al. 1995), and in duct cells and acinar cells of breast (Chalbos et al. 1990). However, the precise expression of FAS protein at the cellular level has not been clarified in normal human tissues, because the FAS antibody used thus far failed to label fat cells in cryostat sections (Joyeaux et al. 1990) and the antibody OA-519 also failed to react with normal or hyperplastic prostate (Shurbaji et al. 1992) and normal breast (Jensen et al. 1995). As shown in this study, our new antibody raised against human brain FAS peptide recognizes FAS protein in human cells and can be used with formalin-fixed, paraffin-embedded tissues. The reason(s) for difference between the antibody described and ours is not clear, but the antibody reported may have a low affinity for the antigen(s) or may recognize a different antigen epitope(s) in the FAS molecule. With immunohistochemistry, we showed that FAS is present in various cells of tissues from the human fetus, newborn/infant, and adult. There are two types of FAS expression, one of which is closely associated with cell proliferation and constitutive expression in resting cells. In this study, the proliferative activity was evaluated with anti-Ki-67. The suitability of monoclonal anti-Ki-67 antibody for detecting proliferative activity was established previously because Ki-67 is known to be expressed in nuclei of cells cycling from G1- to M-phase of the cell cycle (Gerdes et al. 1984a,b). The expression of FAS associated with cell proliferation was clearly observed in human fetal tissues at 20 weeks of gestation. FAS was detected not only in developing epithelial cells but also in growing mesenchymal cells. FAS-containing cells were not always Ki-67-positive, but the frequency of negative cells was consistently lower than that of positive ones. Because the half-life of FAS mRNA is 4 hr (Xu and Rooney 1997) and Ki-67 expression lasts from G1- to the M-phase of the cell cycle, this finding appears reasonable. However, cardiomyocytes, seminiferous tubules, and esophagogastrointestinal tract smooth muscle cells lacked FAS expression despite their apparent Ki-67 labeling. In the FAS-negative proliferating cells, another type of FAS that does not react with the antibody we used might be present, since there are minor differences in the amino acid sequences of brain FAS and FAS in the hepatocellular carcinoma cell line HepG2 (Jayakumar et al. 1995). Alternatively, fatty acid binding protein, which removes fatty acids from the circulation and is rich in fetal and adult heart (Watanabe et al. 1993), might function in the heart in place of FAS. Some resting cells of the 20-week fetus were also FAS-positive. These included adipocytes, ganglion cells of peripheral nerves or sympathetic ganglia, Leidig cells of testis, sebaceous gland, uroepithelium, and fetal adrenocortical cells. Cells of this type might utilize FAS for active lipid metabolism and, among these fat cells, sebaceous glands and umbrella cells of urinary bladder remained FAS-positive in adult tissues. Although a few replicating cells in stomach and duodenum, as well as bone marrow cells, were FAS-positive in the adult, positive cells were found mostly among resting cells that have active lipid metabolism and/or hormone sensitivity. Sebaceous gland cells, fat cells, hepatocytes, and Type II alveolar cells (surfactant or dipalmitate lecithin producers) are cells with active lipid metabolism. Apocrine gland (sex hormone-sensitive), prostate, endometrium, mammary glands, seminal vesicle (testosterone-sensitive), and anterior pituitary cells are hormone-sensitive cells. Adrenocortical cells show both characteristics. Several cells that express FAS do not fall into these two categories: ganglion cells of cerebral cortices, glia, basket cells of cerebellum, mast cells, decidua, and cytotrophoblasts. Basket cells of cerebellar cortex are well-known inhibitory neurons, and similar inhibitory neurons, also called basket cells, exist in the hippocampus but these lacked FAS, indicating that FAS expression is not a characteristic of all inhibitory neurons. The physiological role of FAS in basket cells and some neurons of the cortex or sympathetic ganglia might be elucidated by neurophysiological research.
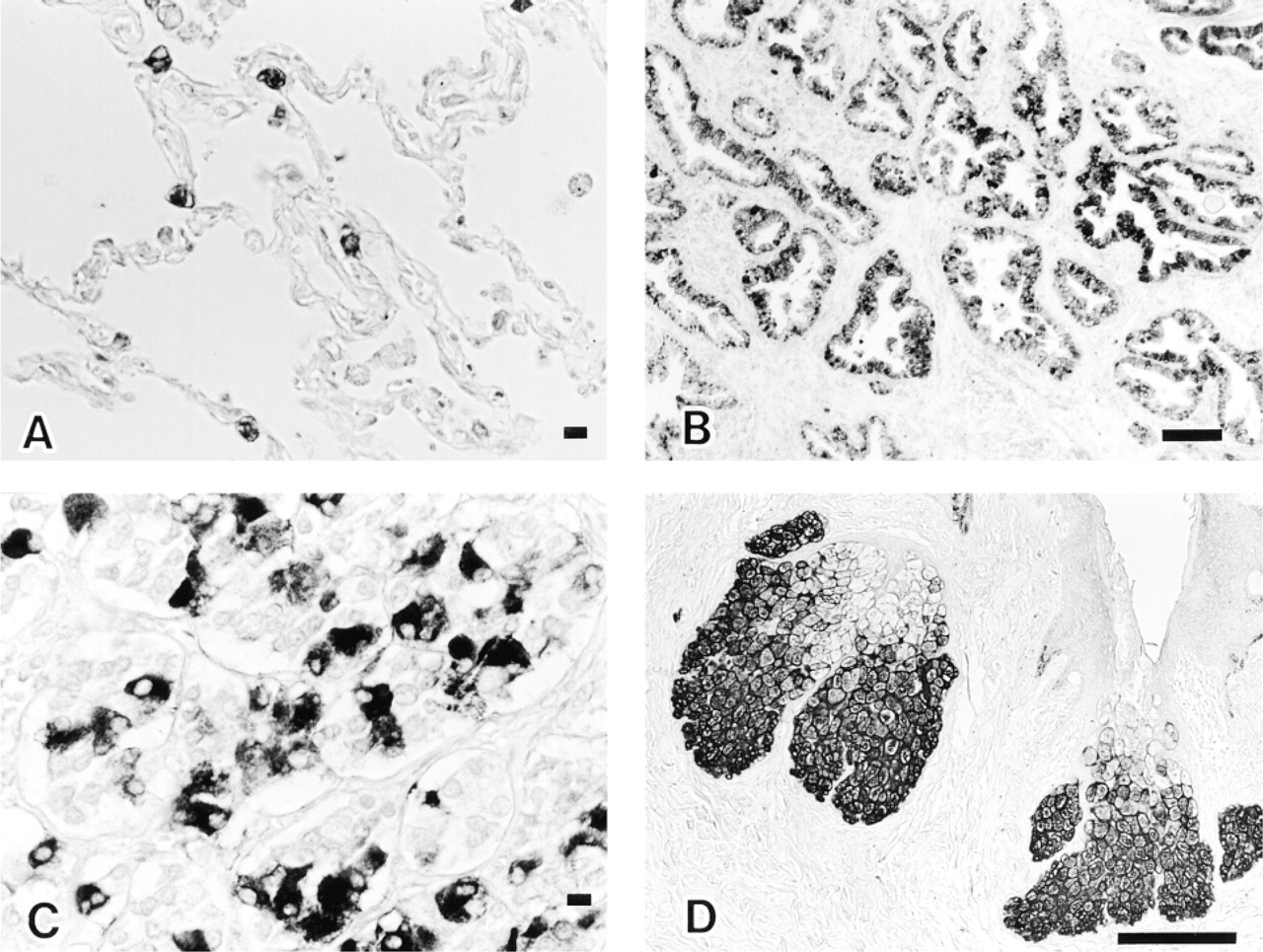
Some Type II alveolar cells of lung (
FAS-positive cells in fetus at 20 weeks of gestation

aAlso positive in nonproliferative state.
FAS-positive cells (
FAS (
It was reported that decidualized stroma cells in the late secretory phase and decidua of pregnancy were intensely positive for FAS not associated with cell proliferation (Pizer et al. 1997). In the present study, these phenomena were confirmed and cytotrophoblasts of chorionic villi also contained FAS, but its biological significance in pregnancy requires investigation. Hormone dependency of FAS expression has been described in human cycling endometrium (Pizer et al. 1997), as stated earlier, and hormonal stimulation induces upregulation of FAS expression in hormone-sensitive cells such as normal breast (Joyeaux et al. 1990), mastopathy, breast carcinoma (Chalbos et al. 1990), lactating breast (Smith and Ryan 1979), and a prostate cancer cell line (Swinnen et al. 1997). Insulin administration has been reported to rapidly induce FAS mRNA in diabetic mouse liver (Paulauskis and Sul 1989), and dexamethasone causes induction of FAS in cultured fetal rat lung (Pope et al. 1988). In these cases, the relationship between the induction of FAS and cell growth is not clear, whereas growth factors such as fibroblast growth factor-1 induce cell growth and upregulate FAS expression in fibroblasts (Hsu et al. 1993). The precise biological significance of FAS expression in normal and cancer tissues is not as clear, but FAS might be expressed and synthesized in coordination with increased demand for fatty acid metabolism and/or membrane synthesis in response to regulated or unregulated cell proliferation, and intense expression of the FAS in surface uroepithelium of both fetus and adult may play a role in excluding water through the hydrophobic nature of fatty acids.
In conclusion, under normal conditions FAS is highly expressed not only in proliferating cells and those with high lipid metabolism but also in hormone-sensitive cells and those with permeability barriers.
Footnotes
Acknowledgements
Acknowledgments
Supported in part by a Grant-in-Aid for Scientific Research (no. 10670168) from the Ministry of Education, Science, Sports and Culture of Japan.
We thank H. Hiraki for his outstanding help with the immunohistochemical studies.