
Abstract
Reticulocalbin (RCN) is one member of the Ca2+-binding proteins in the secretory pathway and is localized in the endoplasmic reticulum. RCN may play a role in the normal behavior and life of cells, although its detailed role remains unknown. Overexpression of RCN may also play a role in tumorigenesis, tumor invasion, and drug resistance. The new antibody for human RCN is used in the distribution of RCN in normal human organs of fetuses and adults with or without inflammation. Immunohistochemical examination demonstrated a broad distribution of RCN in various organs of fetuses and adults, predominantly in the endocrine and exocrine organs. However, RCN expression was heterogeneous in each constituent cell of some organs. Among non-epithelial organs, vascular endothelial cells, testicular germ cells, neurons, and follicular dendritic cells showed strong staining. Plasma cells were the only RCN-positive cells among hematopoietic and lymphoid cells. In inflammatory conditions, RCN expression was enhanced in both epithelial and non-epithelial cells. Heterogeneous expression of RCN indicates that the amount of RCN needed for cell behavior and life may be variable, depending on each cell type and, therefore, RCN may be helpful in establishing the cell origin of neoplasms in some organs. However, further study is needed to establish the significance of RCN in tumorigenesis and in some peculiar features of neoplasms.
Ca2+
A similar Ca2+-binding protein localized in the secretory pathway was initially found in the mouse teratocarcinoma cell line and was designated as reticulocalbin (RCN) (Ozawa and Muramatsu 1993). Subsequently, a human RCN gene was cloned from the human transitional cell carcinoma cell line (Ozawa 1995). RCN also possesses multiple EF-hand motifs and localizes strictly in the ER. At present, in addition to RCN, some RCN family proteins are identified in both vertebrates and invertebrates (Honore and Vorum 2000). Unfortunately, the fundamental properties of these RCN family proteins are still unknown, but they may play an important role in the maintenance of normal cell behavior and life because deletion of a region containing the RCN gene is lethal for mice (Kent et al. 1997).
To understand the pathophysiological roles of RCN, it is inevitably required to examine the expression of RCN in normal tissues and changes under various pathological conditions. We will describe the application of a recently established monoclonal antibody against RCN, demonstrating the heterogeneous cytoplasmic expression of RCN in various cell types of each organ and discuss the implication of RCN expression for pathophysiological roles.
Materials and Methods
Cell Lines and Tissues
The human lung adenocarcinoma cell line (LAC-1), human acute myelogenous leukemia cell line (KY-821), and human ovarian cancer cell line (FOC3) were maintained in our laboratory and used in the analysis of RCN mRNA and Western blotting. Ethical approval was obtained from the Fukushima Medical University local research ethics committee, and informed written consent was obtained from patients. Normal human tissues were obtained from autopsy cases and surgically resected organs. Elements from fetuses at 8 to 12 weeks gestation were obtained from uterine contents upon spontaneous abortion from unknown etiologic factors. All tissues were fixed with 10% buffered formalin and embedded in paraffin using routine methods.
Antibodies
The monoclonal antibody to RCN (clone 6A1; IBL, Takasaki, Japan) was used as described elsewhere (Hirano et al. 2005). The monoclonal antibody to Ki-67 was purchased from Immunotec (Marseille, France).
Extraction of RNA
Total RNA was extracted using the acid-phenol-chloroform method as described elsewhere. RT-PCR was carried out using Takara Ver. 2.1 RT-PCRkit (Takara Co.; Shiga, Japan). Primers used in this study were described by Liu et al. (1997) as follows: RCN, 5′-CCAGGACGGGAAGTTAGACAAAGACAAAGATGA-3′,5′-ATTACTGCCTCCCTCAACCCACTCACC-3′, β-actin, 5′-TGACGGGGTCACCCACACTGTGCCCATCTA-3′, 5′-CTAGAAGCATTTGCGGTGGACGATGGAGGG-3′. These primers were all purchased from Bekkusu Co. (Tokyo, Japan). Cycling reactions were performed with an Astek DNA thermal cycler (Fukuoka, Japan). PCR reaction consisted of a denature time of 1 min at 94C, annealing time of 2 min at 50C, and an extension time of 2 min at 72C, with a final extension of 10 min at 72C at the end of 30 cycles. PCR products were applied to 1.2% agarose gel.
Western Blotting
Cells of each cell line were homogenized in a lysis buffer containing 0.1% SDS and protease inhibitors (Proteinase Inhibitors Set; Roche Applied Science, Tokyo, Japan). The supernatants were mixed with a sample buffer in a 2:1 ratio. Protein concentration was determined using the Bradford method, and 50 μg protein from each sample was subjected to SDS-PAGE and then blotted onto a PVFD membrane (Hybond P; Amersham Biosciences, Arlington, IL). The membrane was incubated with diluted anti-RCN antibody (1:2000 in PBS) overnight at 4C, followed by washing with PBS containing 1% NP40 twice, and subsequent washing with PBS containing 0.05% Tween 20 three times. Subsequently, the membrane was incubated with anti-mouse rabbit antibody coupled with horseradish peroxidase (Nichirei Co.; Tokyo, Japan). Peroxidase reaction products were visualized using the ECL Plus detection system (Amersham Biosciences).
Immunohistochemistry
Detailed immunohistochemical procedures have been described elsewhere (Kusakabe et al. 2000). In brief, 4-μm-thick sections were deparaffinized, rehydrated in graded ethanols, treated with 3% hydrogen peroxide in methanol for 30 min to block endogenous peroxidase, and then treated with 5% skim milk (Yukijirushi Co.; Sapporo, Japan) in PBS for 60 min to avoid nonspecific absorption of immunoglobulin. Concentration of the primary RCN antibody was 0.05 μg/ml, and diluted anti Ki-67 (1:100 with PBS) was used. Sections were incubated in an electronic oven for 15 min in 0.01 M citrate buffer, pH 6.0, to retrieve the antigen. All slides were incubated overnight with the antibody at 4C, followed by incubation with biotin-labeled anti-mouse IgG (Nichirei Co.) for 60 min, and then streptavidin-conjugated peroxidase (Histofine Kit; Nichirei Co.) for 60 min at room temperature. Localization of RCN was visualized with 5 mg diaminobenzidine (Dojin; Kyoto, Japan) of NaN3/dl in 0.05% H2O2. Nuclei were counterstained with 1% hematoxylin.
Specificity of the primary antibody was confirmed by omission of the primary antibody and by use of preimmune mouse immunoglobulin for the first step of the immunostaining procedure. In addition, because full-length RCN DNA has been cloned (Hirano et al. 2005), recombinant RCN was purified and used for absorption of the primary antibody. Sections of lung cancer and breast cancer were used as positive controls (Liu et al. 1997; Hirano et al. 2005). In addition, because the FOC3 ovarian cancer cell line showed expression of RCN mRNA, a section obtained from serous papillary ovarian carcinoma was also used as a control.
Results
Specificity of Anti-RCN Antibodies and Validity of Immunoreaction
Results of RT-PCR demonstrated a 573-bp band in LAC-1 and FOC3 cell lines and a fainter similar band in the KY-821 cell line (Figure 1).
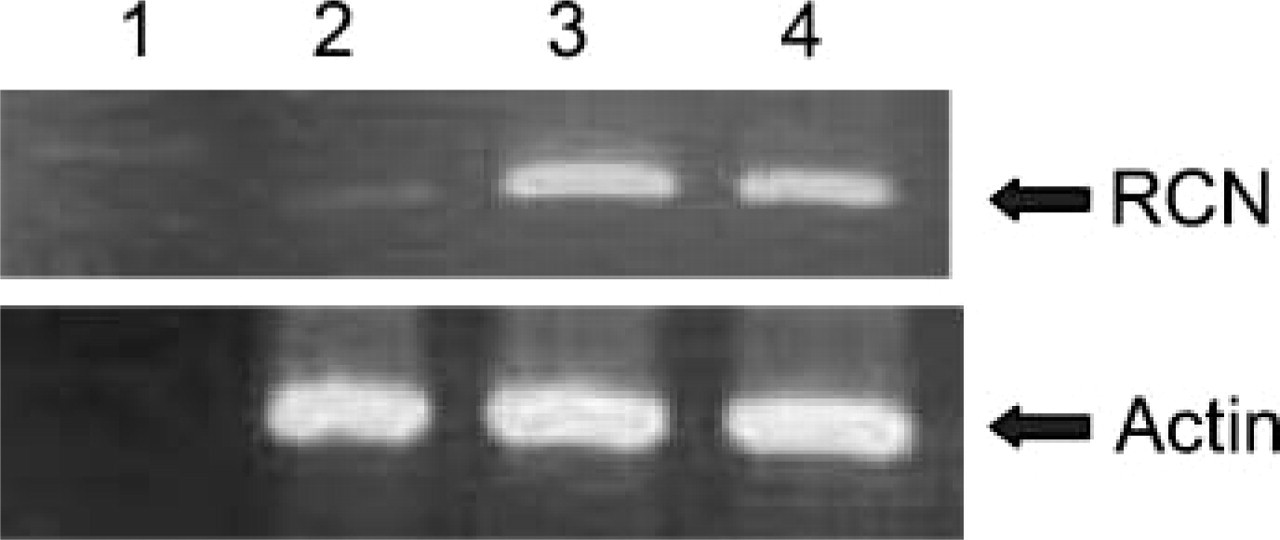
PCR products. Lane 1, marker; Lane 2, KY-821; Lane 3, LAC-1; Lane 4, FOC3. LAK-1 and FOC3 demonstrate strong expression of reticulocalbin (RCN) mRNA, and KY-821 does weakly.
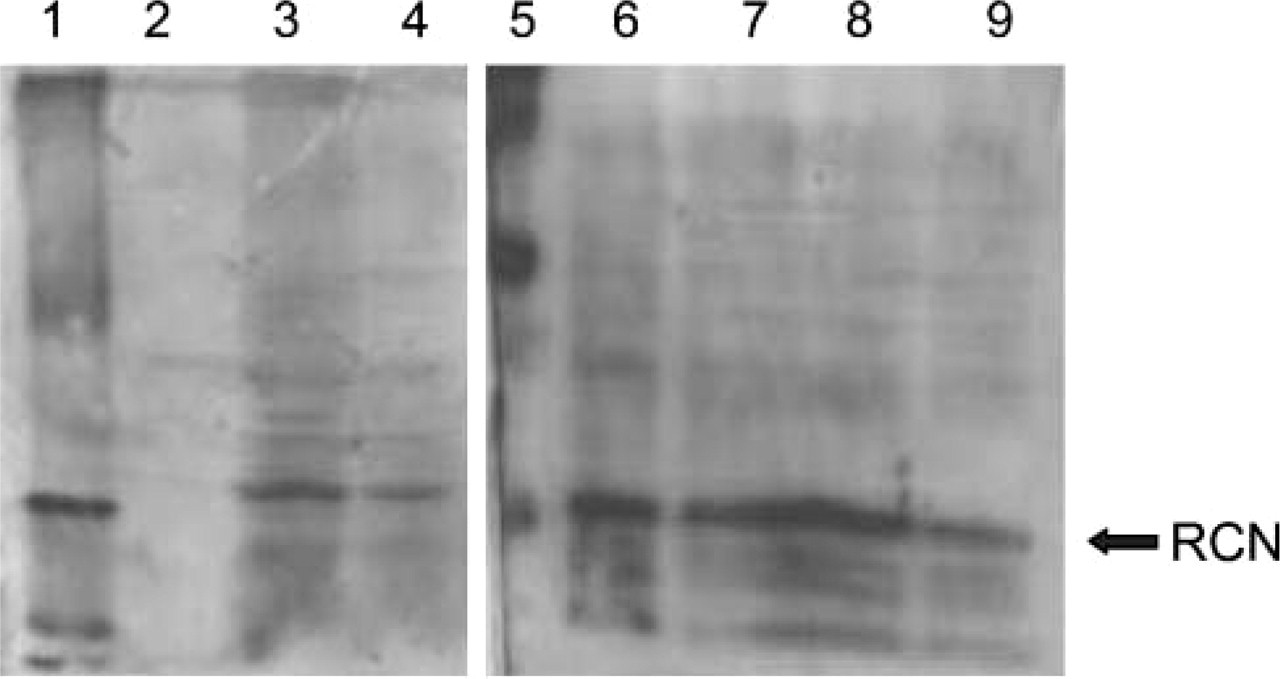
Lane 1, marker; Lane 2, KY-821; Lane 3, LAC-1; Lane 4; FOC3; Lane 5, marker; Lane 6, colon; Lane 7, liver; Lane 8, testis; Lane 9, pancreas. Corresponding to PCR results, LAC-1 and FOC3 apparently express RCN, but KY-821 does not.
RCN antibody reacted with a 44-kDa protein in the extracts of both LAC-1 and FOC3 cell lines but did not in the KY-821 line when using Western blotting (Figure 2). RCN antibody also reacted with the same protein in the extracts taken from normal colons, livers, testes, and pancreata. The absorbed antibody and normal mouse IgG failed to show any bands using Western blotting.
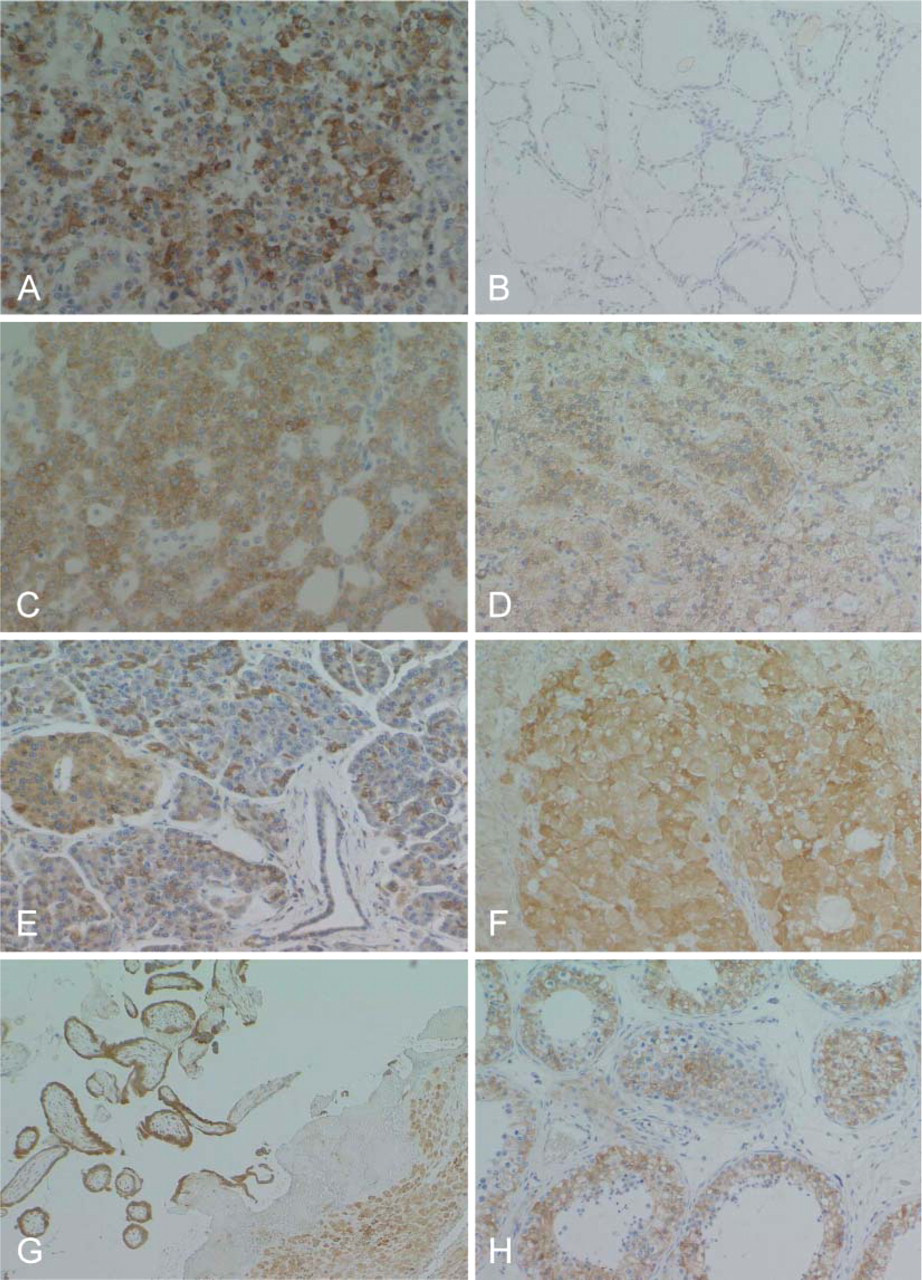
The optimal condition of immunostaining with anti-RCN antibody was established using immunohistochemistry with human lung, breast, and ovarian cancers (Figures 3A-3C). The same tissues were negative when the normal mouse IgG and an absorbed antibody with recombinant RCN were used. RCN was distributed homogeneously in the cytoplasm of each cell.
Distribution of RCN in Fetuses
RCN was distributed extensively in various organs. In the central nervous system, ependymal cells, neuroblasts, and a minority of glial cells were positive (Figure 4A). In visceral organs, liver cells (Figure 4B), epithelial cells from alimentary tracts and pancreata, alveolar epithelial cells, epithelial cells from the kidney (Figure 4C), adrenal cortical cells (Figure 4D), chondrocytes (Figure 4E) and osteoblasts (Figure 4F), vascular endothelial cells (Figure 4G), skeletal muscle cells (Figure 4H), and germ cells in the testes were strongly positive for RCN. RCN-positive stromal cells were scattered throughout the stroma in each organ. Skin and hematopoietic cells were negative for RCN (Figure 4B).
Distribution of RCN in Adults
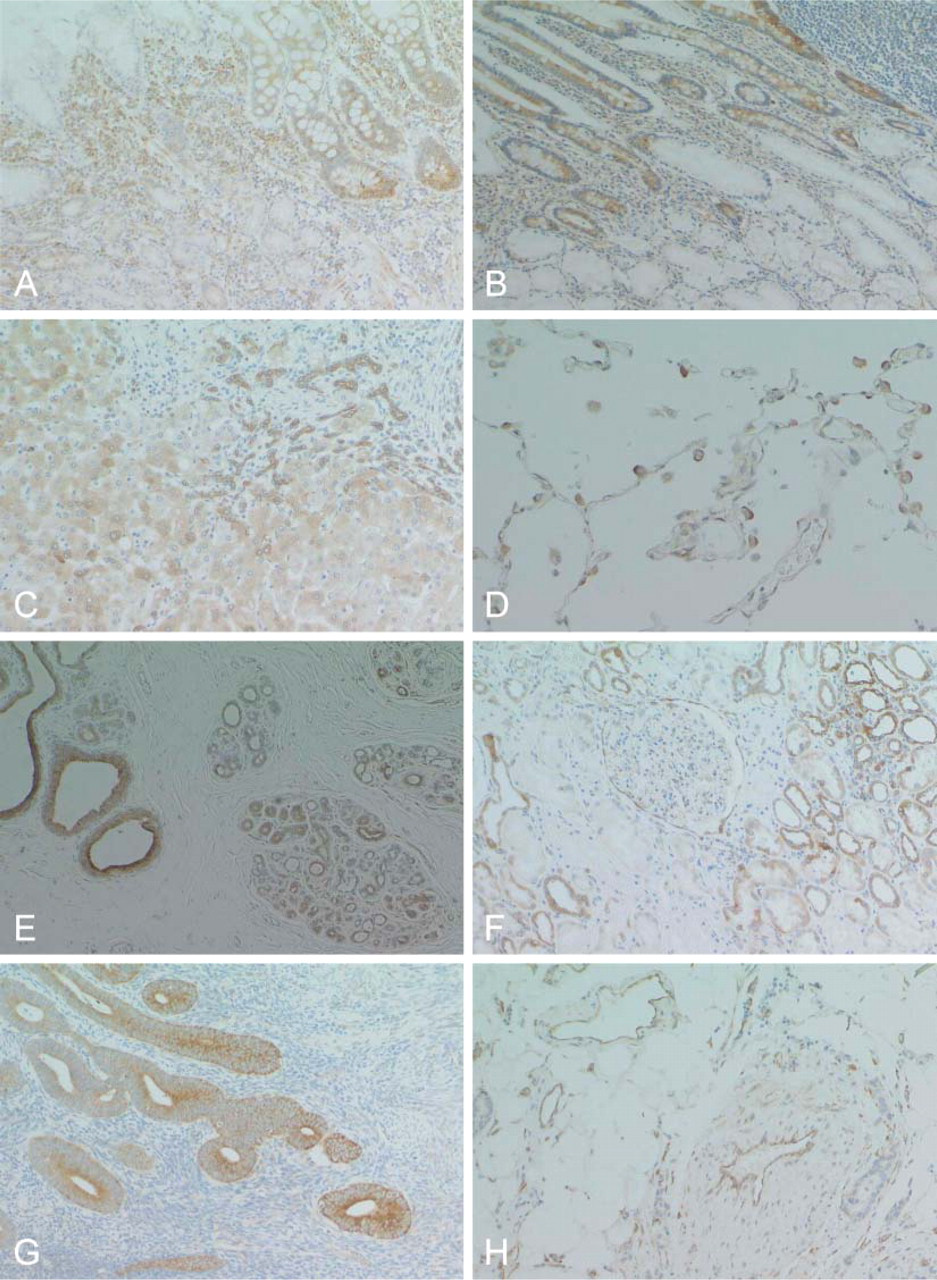
RCN was also expressed in various organs in adults, but its expression was heterogeneous in the cells of each organ (Table 1). RCN demonstrated variable positive staining in epithelial cells in a wide range of endocrine and exocrine organs, with the exception of several organs (Figures 5A-5E, Figures 6A-6G). Follicular epithelial cells from the thyroid gland showed no, or a very weak, positive stain (Figure 5B). Epithelial cells of mucinous glands from the salivary glands, fundic, pyloric and duodenal glands (Figures 6A and 6B), and pancreatic acinar cells (Figure 5E) were largely negative. Epithelial cells of gastric foveolar epithelium in the zone of proliferation showed moderate positive staining, although most were negative (Figure 5A). In the kidney, glomeruli and epithelial cells of the proximal tubules were negative (Figure 6F). In the female genital system, ova were negative, and sex cord cells and ovarian stromal cells showed weak positive staining. Corpus luteum was strongly positive (Figure 5F). Endometrial glands were strongly positive (Figure 6G), but cervical glands were negative. Trophoblasts and decidual tissue were also strongly positive (Figure 5G). In the male genital system, germ cells in various differentiation stages were strongly positive, and Leydig cells were weakly positive (Figure 5H). Squamous cells of the skin and esophagus contained no RCN-positive cells. Vascular endothelial cells and medial smooth muscle cells from both arteries and veins showed apparent positive staining (Figure 6H). Sinus endothelial cells of the spleen showed strong positive staining (Figure 7A). In the soft tissues and stroma of each organ, a small number of fibroblasts and adipocytes were positive (Figure 6H). In addition, activated fibroblasts in an inflammatory condition (Figure 7B) and around cancer tissue strongly expressed RCN. However, endometrial stromal cells were entirely negative (Figure 6G). Skeletal muscle was also strongly positive, and smooth muscles in each organ showed weak cytoplasmic staining in a limited or dot-like manner (Figure 7C). In the skeletal system, osteoblasts and chondrocytes were strongly and weakly positive, respectively, but osteoclasts and osteocytes were negative. Synovial cells were also weakly positive but strongly positive when inflammation involved the joint. Immature or mature hematopoietic cells and lymphocytes, including those of a transformed type, were largely negative (Figures 7D and 7F) except for plasma cells that showed diffuse staining in the cytoplasm (Figure 7E). Interestingly, follicular dendritic cells and sinus reticulum cells in the lymph node were strongly and weakly positive, respectively (Figure 7F). In the brain and spinal cord, neurons in each site except for Purkinje cells (Figures 7G and 7H) and activated astrocytes in various conditions also showed strong staining (Figure 7I). Choroidal plexus cells, granular cells, and Bergman astrocytes in the cerebellum (Figure 7H) demonstrated moderate to weak staining. However, oligodendrocytes were largely negative. In contrast, ganglion cells in the sympathetic nerve, Auerbach and Meisner plexus, and adrenal medulla were negative, whereas Schwann cells showed weak staining.
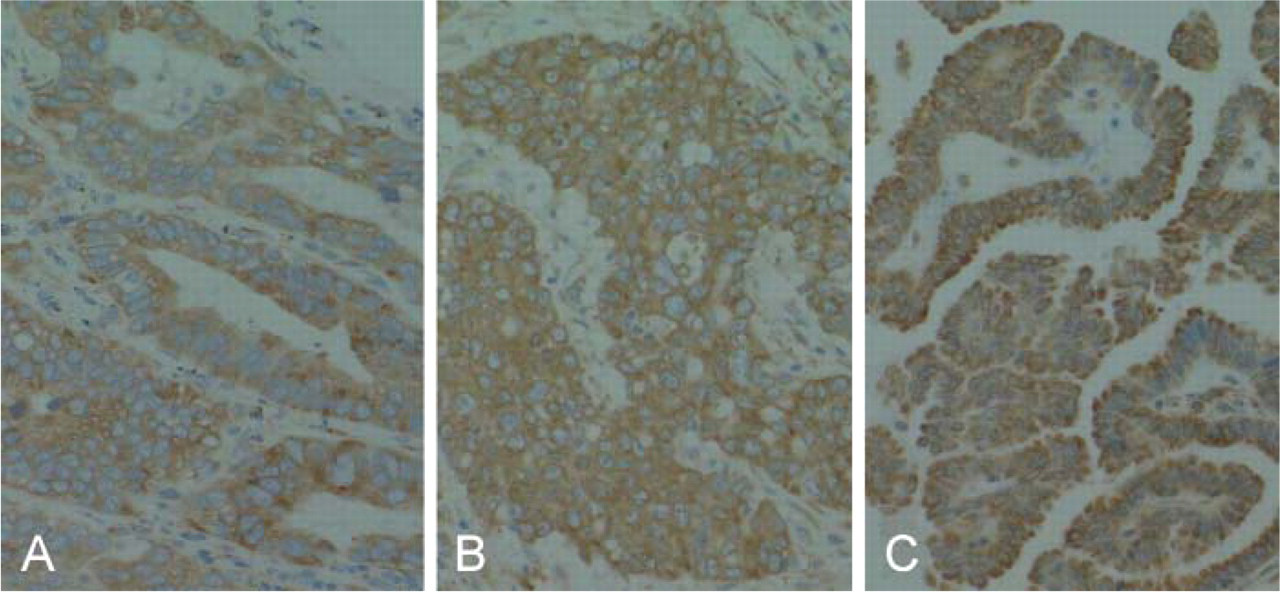
Adenocarcinoma of the lung (
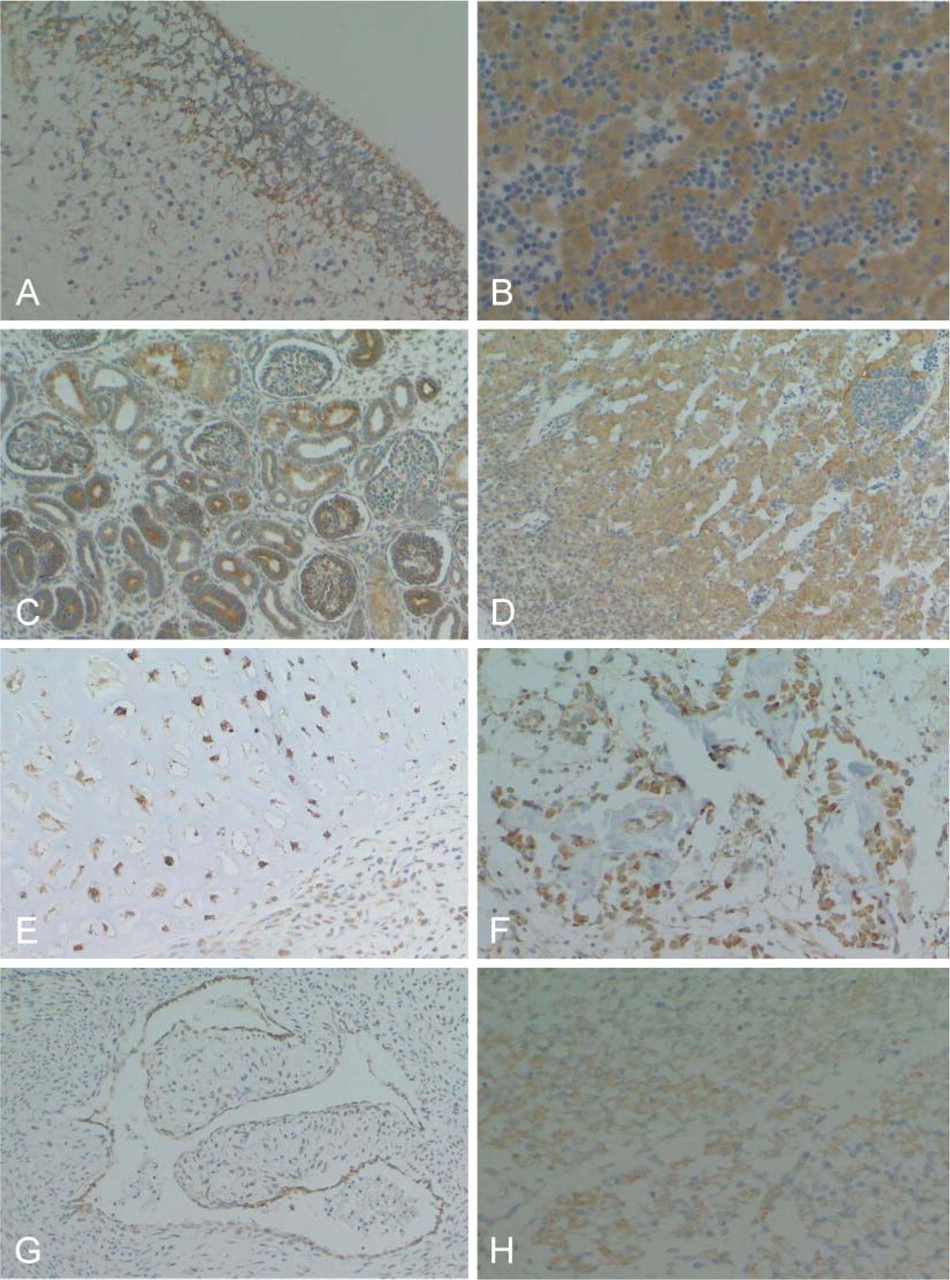
RCN is expressed in various fetal organs. Ependymal cells, neuroblasts, and glial cells in brain (
Reticulocalbin expression of various organs in adult


Scattered.
Epithelial cells of neck zone in foveola (+).
(−), negative; (+), weakly positive; (++), strongly positive.
Cells of various endocrine organs with the exception of the thyroid gland show strong staining of RCN. (
Similarly, epithelial cells in various organs stain positive to various degrees. (
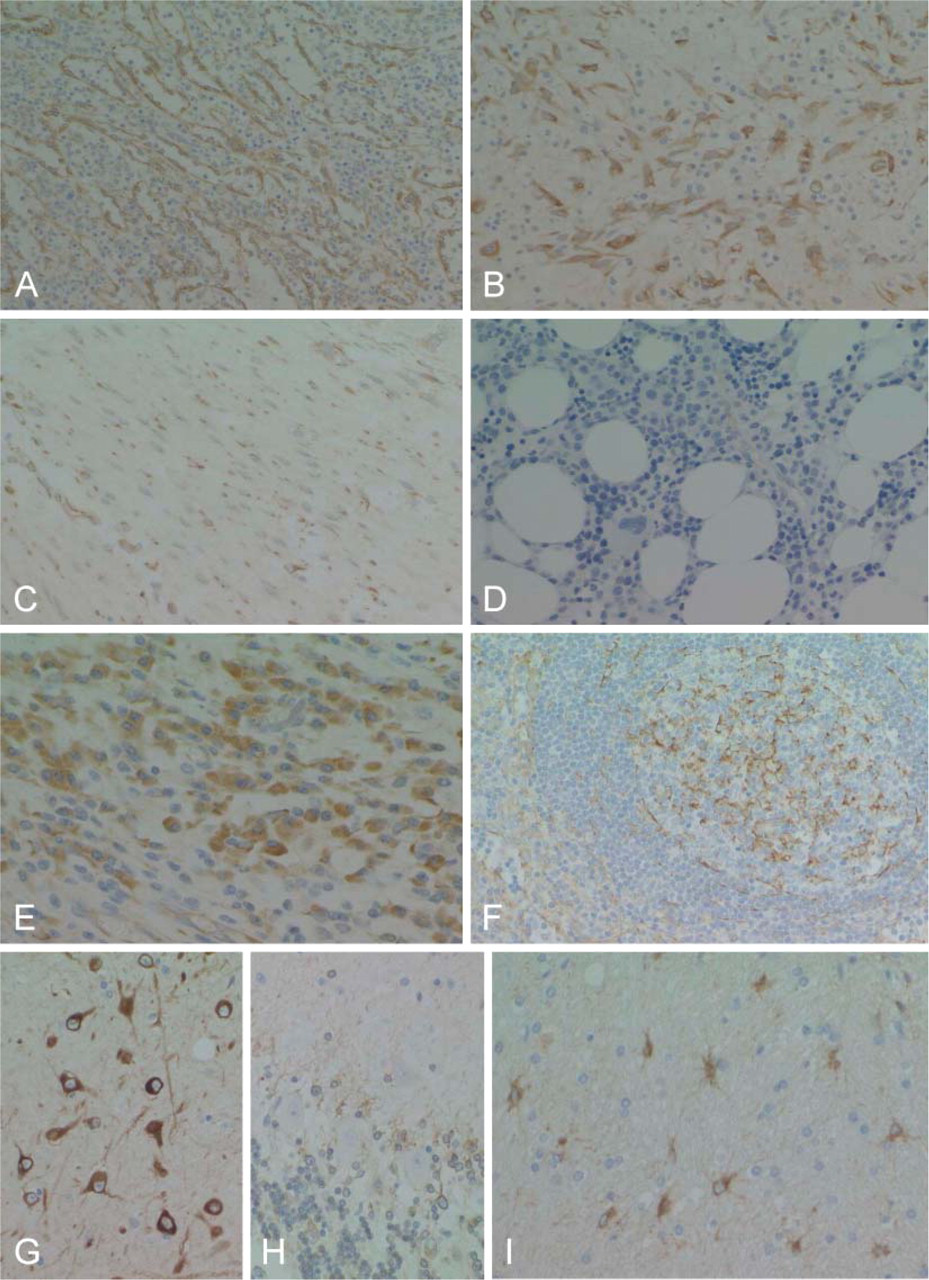
Sinus cells of the spleen also show strong staining (
Activated fibroblasts demonstrated diffuse and strong staining of RCN in inflammatory lesions or fibrosis around cancer in any parts, and RCN expression was enhanced in vascular endothelial cells under inflammatory conditions. Furthermore, proliferating small bile ducts in the portal area of the cirrhotic liver strongly expressed RCN (Figure 6C). Epithelial cells of the bronchus and cervical glands of the uterus also showed strong staining when inflammation occurred in these regions, although uterine cervical glands were negative under normal conditions. Metaplastic gastric epithelial cells showed strong staining (Figure 6A).
Discussion
RCN is a Ca2+-binding protein containing EF-hand motifs and is localized in the ER. Mammalian RCN was initially discovered in mice and detected in different types of cultured cells (Ozawa and Muramatsu 1993). Subsequent research in humans has demonstrated the ubiquitous expression of RCN mRNA in various organs (Ozawa 1995). Although the exact functions of RCN remain unknown, it may play important roles in the maintenance of normal cell behavior and cell life because homozygous deletion of the RCN gene is lethal for mice (Kent et al. 1997). However, the amount of RCN may be at low levels because most RCN family proteins were detected at low levels in various organs (Honore and Vorum 2000). Overexpression of RCN has been reported in human hepatocellular carcinoma cell lines (Yu et al. 2000), suggesting a close correlation between overexpression of RCN and malignant transformation. In addition, ERC55, one of the RCN family proteins, was reported to play an important role in the oncogenesis of human cervical carcinomas infected by the papilloma virus (Chen et al. 1995). Furthermore, Liu et al. (1997) reported that the mammary cancer cell line with an aggressive nature expressed RCN, but non-invasive lines did not. Hirano et al. (2005) reported decreased RCN expression in cisplatin-resistant human lung cancer. In this context, the study of RCN expression in each cell in normal organs is important to understand pathophysiological roles under various pathological conditions and its implications in oncogenesis of RCN.
In addition to RCN, the RCN family proteins include ERC55 (Weis et al. 1994), crocalbin (Hseu et al. 1999), Cab45 (Vorum et al. 1998), and calumenin (Koivu et al. 1997). RCN and ERC55 are strictly localized to the ER, Cab45 is localized to the Golgi complex, and calumenin is distributed throughout the whole secretory pathway. These proteins are all ubiquitously expressed in various human organs, as in the same manner of RCN. However, because heterogeneous expression of these proteins has been demonstrated by Northern blotting (Weis et al. 1994; Ozawa 1995; Koivu et al. 1997), it is possible that each of the RCN family proteins may be expressed at different levels among the various organs or cells. Unfortunately, because there is no report of the detailed study on the RCN family proteins in cell levels, the expression and amount of the RCN family proteins in each constituent cell of various organs remain obscure. We believe that the new monoclonal antibody for RCN used in this study recognizes RCN specifically because of complete absorption with recombinant RCN, different amino acid residues of immunogen from other RCN family proteins, and the positive expression in the cytoplasm of each cell but not in the Golgi complex.
Because RCN plays a role in the mechanism of normal cell behavior, it is reasonable that RCN mRNA be detected in various human organs. However, slight heterogeneity of RCN mRNA was demonstrated, i.e., there were lower levels in the brain and kidney when compared to other organs examined (Ozawa 1995). This finding prompts us to consider that expression of RCN may be different in cells from various organs. Because RCN is a specific protein localized in secretory pathways, strictly in ER, it is likely that cells with synthesis and/or secretion functions for various substances have more amounts of RCN compared with cells that are comparatively at rest. Indeed, cells from various endocrine organs are strongly positive for RCN, except for thyroid gland cells that showed no or extremely weak positive staining. Epithelial cells in a wide range of various organs also expressed RCN to various degrees, with the exception of squamous cells. These findings indicate that RCN plays an important role in synthesis and/or secretion functions in various types of epithelial cells. In addition, because epithelial cells with no or weak RCN expression under normal conditions, such as uterine cervical glands, showed enhanced expression of RCN under inflammatory conditions, RCN levels needed for cell behavior vary under normal conditions. Unexpectedly, epithelial cells from some types of organs, including those from the pancreatic acinus and from the fundic, pyloric, and duodenal glands, showed no or weak staining of RCN despite their active synthesis and secretion functions. This finding leads us to consider another possibility that other RCN family protein may function predominantly in some types of cells.
A positive immunoreaction for RCN in some types of non-epithelial cells provides us some new aspects on cell behavior. Vascular endothelial cells can produce some substances through various types of stimulation. However, strong expression of RCN indicates the active synthesis and secretion functions of vascular endothelial cells, even under normal conditions. The exact role of RCN in vascular endothelial cells remains obscure at the present time, but RCN may be useful as a marker for vascular endothelial cells such as CD34 or CD31. Although fibroblast and adipocyte are considered as relatively resting cells, a small number of fibroblasts and adipocytes are positive for RCN, probably reflecting their function of synthesis and secretion of collagen and fat to maintain normal architectures. In addition, diffuse and strong positive immunoreaction for RCN of activated fibroblasts, vascular endothelial cells in the inflammatory lesions, and fibrosis around cancer may reflect their active work in repairing destroyed tissues.
Strong staining of RCN in neurons in any brain and spinal cord sites is of particular interest because these lack endocrine and exocrine functions. However, because neurons can produce various substances including neurotransmitters, strong expression of RCN represents active synthesis functions. Nonetheless, medullary cells from the adrenal gland were entirely negative for RCN despite their evident synthesis and secretion of adrenalin and noradrenalin. Purkinje cells in the cerebellum and ganglion cells in sympathetic nerves were also negative for RCN. At this time, the significance of heterogeneous RCN expression in neurons from different sites remains obscure. On the other hand, the different expression of RCN in glial cells is available for a differential diagnosis of brain tumors.
We have initially speculated that hematopoietic cells may strongly express RCN because of their high proliferating activity. Unexpectedly, most hematopoietic and lymphoid cells, including immature hematopoietic cells of each cell line and transformed lymphocytes, are entirely negative for RCN. Indeed, KY-821 expressed a lower level of RCN mRNA, but Western blotting failed to show RCN bands, indicating an extremely small amount of RCN that could not be detected using these methods. Among hematopoietic cells, only plasma cells were positive for RCN, representing its synthesis and secretion functions for immunoglobulins. In the lymph nodes, follicular dendritic cells are strongly positive for RCN. Follicular dendritic cells are antigen-presenting cells for B cells and have a synthesis or secretion function for some substances such as cytokines (Park et al. 1999,2004), suggesting the role of RCN in secretion function. Despite high proliferating activity of lymphocytes in germinal centers, they are entirely negative for RCN. These findings indicate that RCN may be useful as a new marker for plasma and dendritic cells and for their neoplasms.
We have also speculated that cells with high proliferating activity also strongly express RCN because of active synthesis of various substances in such cells. Indeed, foveolar epithelial cells from the neck zone of the stomach, as well as testicular germ cells, are positive for RCN. However, immature hematopoietic cells and follicular transformed lymphocytes are completely negative for RCN, although they showed a high Ki-67 index. In addition, the Ki-67 index was very low in most cells that strongly expressed RCN. Thus, RCN expression is closely associated with synthesis and/or secretion functions but not with proliferating activities in each cell.
In conclusion, heterogeneity of RCN expression in constituent cells in some organs indicates that RCN expression may be related to functions of each cell and available for establishing cell origin and differential diagnoses of some types of tumors, although an extensive distribution of RCN has been demonstrated in various normal organs. Weis et al. (1994) proposed that the members of RCN family proteins might regulate Ca2+-dependent activities in the ER by modulating the function of other proteins. Indeed, it is reported that ERC55 interacts with the vitamin D receptor (Imai et al. 1997) and the transforming oncoprotein, E6, from papillomavirus (Chen et al. 1995). Research on the interaction between RCN and other proteins including other RCN family proteins is important to understand the role in malignant transformation. Furthermore, our preliminary examination demonstrates no correlation between RCN expression and invasiveness of breast cancers with or without stromal invasion (unpublished data), whereas the mammary cancer cell line with an aggressive nature expressed RCN but non-invasive lines did not (Liu et al. 1997). Further study is needed to establish a correlation between RCN expression and more special features of tumor cells, such as invasiveness or drug resistance.