
Abstract
W
Materials and Methods
Antibody Production
Because MAGP-2 has recently been shown to contain sequences similar to regions of MAGP-1, a new anti-MAGP-2 antibody was prepared using a synthetic peptide (made by Chiron Mimotopes; Melbourne, Australia), SETFTEDPNLVNDPATDET(C), which corresponds to a unique conserved amino acid sequence encoded by cDNA for bovine MAGP-2. The peptide was coupled to diphtheria toxoid and the conjugate was used to raise an antibody in rabbits. The antibody was affinity-purified using MAGP-2-Sepharose columns. These techniques have been described previously (Gibson et al. 1997). Monoclonal antibodies (MAbs) 11B and A5, raised to MAGP-1 and fibrillin-1, respectively, and polyclonal antibody to tropoelastin have been described previously (Henderson et al. 1996; Gibson et al. 1995,1997).
Assessment of Antibody Specificity
The affinity-purified antibody was assessed by ELISA and immunoblotting for crossreactivities with other matrix proteins (Gibson and Cleary 1985). Purified bovine proteins and proteoglycans to be tested were MAGP-2, MAGP-1, fibrillin-1, MP78/70, tropoelastin, collagen Types 1, III, V, and VI (native and pepsin-treated), decorin, and biglycan. The sources and methods of preparation of these proteins have been described elsewhere (Gibson et al. 1997). The specificity of the affinity-purified anti-MAGP-2 peptide antibody was also confirmed by immunoblotting against extracts of nuchal ligament from a 210-day-old fetal calf. Extracts were generated by sequential treatment of tissue homogenate with saline, 1 M NaCl, 6 M guanidinium chloride (GuHCl), saline plus dithiothreitol (DTT), and 6 M Gu-HCl plus DTT as previously described (Gibson et al. 1989).
Immunofluorescence Microscopy
Tissue blocks were taken from adult cattle or 200- to 210-day-old fetal calves within 1 hr of maternal death and snapfrozen in OCT compound. Sections (4 μm thick) were cut using a cryostat and incubated with primary antibody followed by an appropriate secondary antibody coupled to fluorescein as described previously (Gibson and Cleary 1987). For optimal activity, the anti-fibrillin-1 MAb A5 required pretreatment of the tissue sections with 6 M GuHCl/DTT. In these instances the tissue was prefixed with paraformaldehyde (Gibson et al. 1989). Antibodies to MAGP-1, MAGP-2, and tropoelastin were applied to unfixed tissues. Affinitypurified rabbit antibodies were applied at a concentration of 20 μg/ml and the MAbs used were ascites fluid diluted 1:100. Control sections were incubated with IgG (20 μg/ml) purified from preimmune rabbit serum and diluted ascites fluid containing an MAb to the parasite Giardia lamblia (Henderson et al. 1996). Ear cartilage sections were pretreated in a humidity chamber for 90 min at 37C with testicular hyaluronidase (EC 3.2.1.35; Boehringer Mannheim, Sydney, Australia) at a concentration of 0.5 U/μl in 150 mM NaCl, 20 mM sodium acetate, pH 6.0, and washed extensively with PBS. Sections were examined and photographed with automatic exposure using a Nikon microphot FX-A microscope as described previously (Gibson et al. 1997). Control sections were photographed using manual exposure times matching those of the appropriate antibodies.
Immunoelectron Microscopy
Tissues were dissected from a 220-day-old fetal calf within 10 min of maternal death and cut into blocks of approximately 1 mm3. The blocks were washed in five changes of PBS for 1 hr in total, treated with PBS containing 1% ovalbumin for 20 min, and then incubated for 18 hr with affinity-purified anti-MAGP-2 peptide antibody (10 μg/ml in PBS/ovalbumin). Control blocks were incubated with IgG (10 μg/ml) from preimmune rabbit serum in place of the primary antibody. After washing in PBS/ovalbumin (six times for 5 min), the blocks were incubated for 1 hr with protein A coupled to 20-nm gold particles (Sigma; St Louis, MO). After further washing, the blocks were fixed with 1.25% glutaraldehyde and 4% paraformaldehyde, then treated with 1% osmium tetroxide and embedded in Spurr's resin as described previously (Kumaratilake et al. 1991). Because the gold particles do not penetrate far into the tissue, thin sections were cut with the surface of the blocks orientated at an angle of 10-20° to the knife to obtain a part of the surface of the block in each section. The sections were mounted on nickel grids, stained with uranyl acetate and lead citrate, and examined with a JEOL 100 transmission electron microscope.
Northern Blotting
Tissue samples were dissected from fetal and adult bovines of defined ages (110-day-old fetuses to 2-year-old adult) within 1 hr of maternal death and immediately frozen in liquid nitrogen. The frozen tissues were powdered and total RNA was isolated by guanidine isothiocyanate extraction followed by CsCl density gradient centrifugation. RNA samples (10 μg) were electrophoresed on 1% agarose and the quality of RNA and equal loading of lanes were confirmed by staining with ethidium bromide. The RNA was then transferred to nylon membranes and hybridized with [32P]-dCTP-labeled cDNAs for bovine MAGP-1 (clone cM32), bovine MAGP-2 (clone cI5), or human tropoelastin (pcHEL 2) using previously described methods (Gibson et al. 1991, 1995,1996). Relative mRNA levels were measured by densitometric scanning of bands visualized on XAR-5 film (Kodak; Rochester, NY) after exposure to the membranes at −70C with Biomax MS intensification screens (Kodak). To check for non-uniform RNA loading and transfer, the membranes were stripped of radiolabeled probes by boiling in 0.1% SDS plus 2 mM EDTA for 2 min and rehybridized with [32P]-dCTP-labeled cDNA for the “housekeeping” protein GAPDH (American Type Culture Collection; Rockville, MD), followed by densitometric scanning as above. Scanning values were normalized to GAPDH mRNA when necessary. When different tissues were compared, steady-state mRNA levels for each protein were expressed relative to the value from nuchal ligament tissue (assigned 100 arbitrary units). When nuchal ligament was compared at different stages of development, steady-state mRNA levels were expressed relative to those found in tissue from 2-year-old adult bovines (assigned 1 arbitrary unit). RNA was pooled from at least two animals at each developmental stage.
Results
Antibody Specificity
The affinity-purified anti-MAGP-2 peptide antibody reacted strongly by ELISA and immunoblotting with MAGP-2 (25 kD). No crossreaction was detected with MAGP-1, fibrillin-1, MP78/70 (βig-H3), or tropoelastin (Figures 1Aand 1B). Decorin, biglycan, and collagen Types I, III, V, and VI were also unreactive with the anti-peptide antibody (data not shown). The anti-MAGP-2 peptide antibody was also tested for crossreactivity with unknown tissue antigens by immunoblotting against a series of sequential extracts from fetal nuchal ligament (Figure 1C). The antibody showed no crossreactivity with other proteins solubilized from the tissue. Anti-MAGP-2 peptide antibody stained only bands corresponding to MAGP-2 and its putative degradation/processing product. Only trace amounts of the protein were detected in the 1 M NaCl and 6 M GuHCl extracts (Figure 1C, Lanes 7 and 8). Interestingly, most of the MAGP-2 was extracted only after treating the tissue with a reducing agent (Figure 1C, Lanes 9 and 10), indicating that the protein was disulfide-bonded to insoluble microfibrils. Preincubation of the affinity-purified antibody with MAGP-2 or MAGP-2 peptide completely blocked its activity as measured by ELISA, immunoblotting, and immunofluorescence. The evidence confirmed the specificity of the anti-MAGP-2 peptide antibody that was used for the immunohistochemical investigations described below.
Immunofluorescence
The comparative distributions of MAGP-2 and MAGP-1 were determined in a range of elastic and nonelastic tissues from a 210-day-old fetal calf and in some adult bovine tissues. Control sections were incubated with preimmune rabbit IgG or with anti-Giardia MAb instead of primary antibody. When photographed for the same exposure time as sections treated with the primary antibody, the controls showed no fluorescence on any of the tissues (not shown). All tissues were obtained from a 210-day-old calf unless otherwise stated.
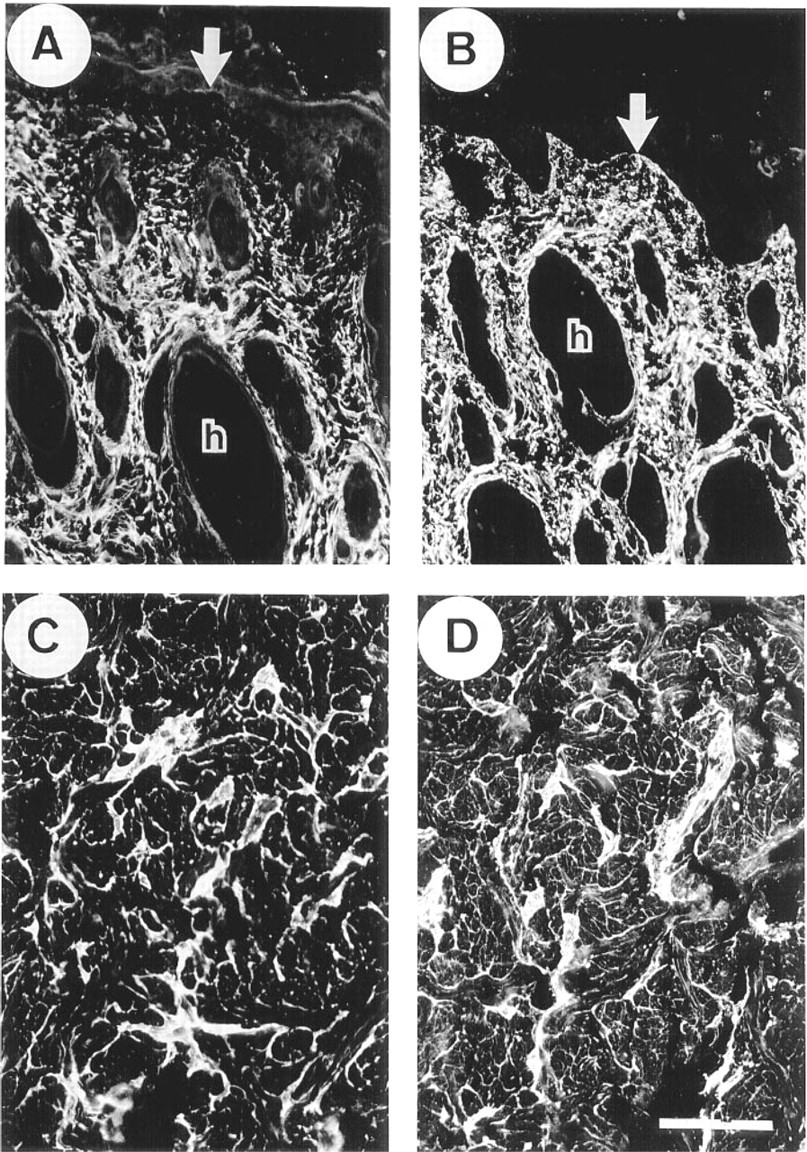
Elastic Tissues. In nuchal ligament, the anti-MAGP-2 antibody localized specifically to elastic fibers (Figure 2A). The staining pattern was indistinguishable from those of antibodies to MAGP-1 (Figure 2B), fibrillin-1 (not shown), and tropoelastin (not shown). In lung, MAGP-2 was immunolocalized strongly on elastic fibers surrounding bronchioles (Figure 2C), with a staining pattern that was similar to those of MAGP-1 (Figure 2D) and fibrillin-1 (not shown). Anti-MAGP-2 immunostaining was also evident on the elastic fibers of the surrounding parenchymal tissue, but this was relatively much weaker than that detected with anti-bodies to MAGP-1 or fibrillin-1. In addition, MAGP-2 was identified only in the internal elastic lamina and adventitia of major blood vessels, in contrast to MAGP-1 and fibrillin-1, which were also localized in the muscular medial layer of this tissue. The anti-MAGP-2 antibody also strongly stained elastic tissue in the interlobular fibrous septa (Figure 2E), which were relatively less prominently stained by the anti-MAGP-1 antibody (Figure 2F).
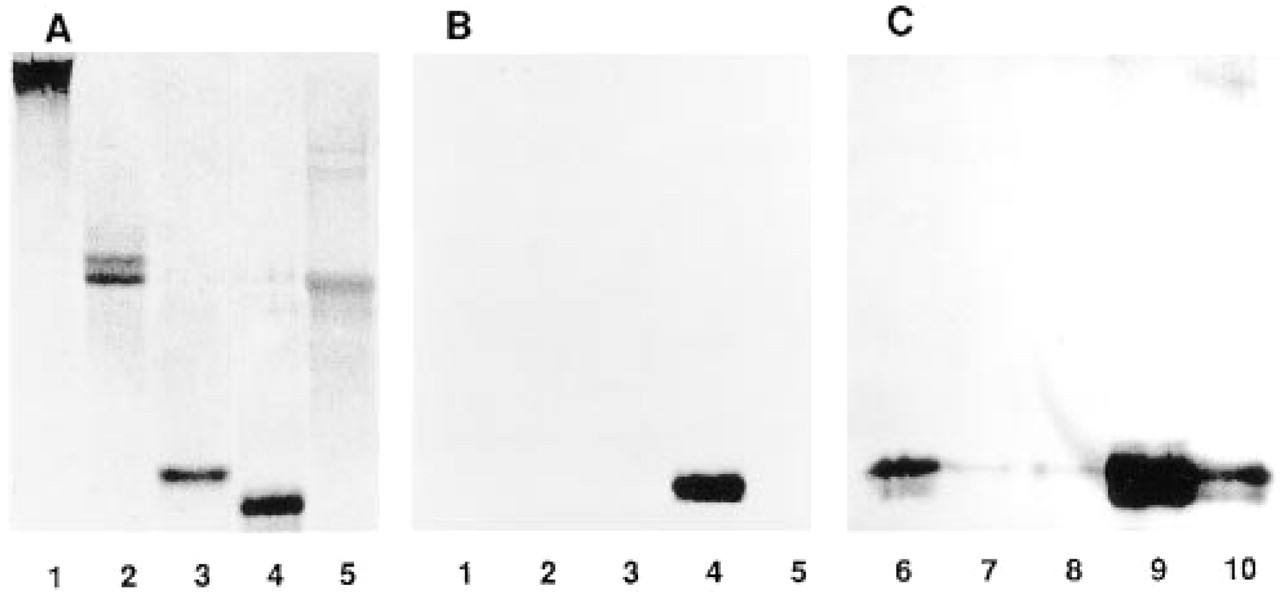
Immunoblot analysis of anti-MAGP-2 peptide antibody. Purified ligament proteins were analyzed by SDS-PAGE on 10% gels
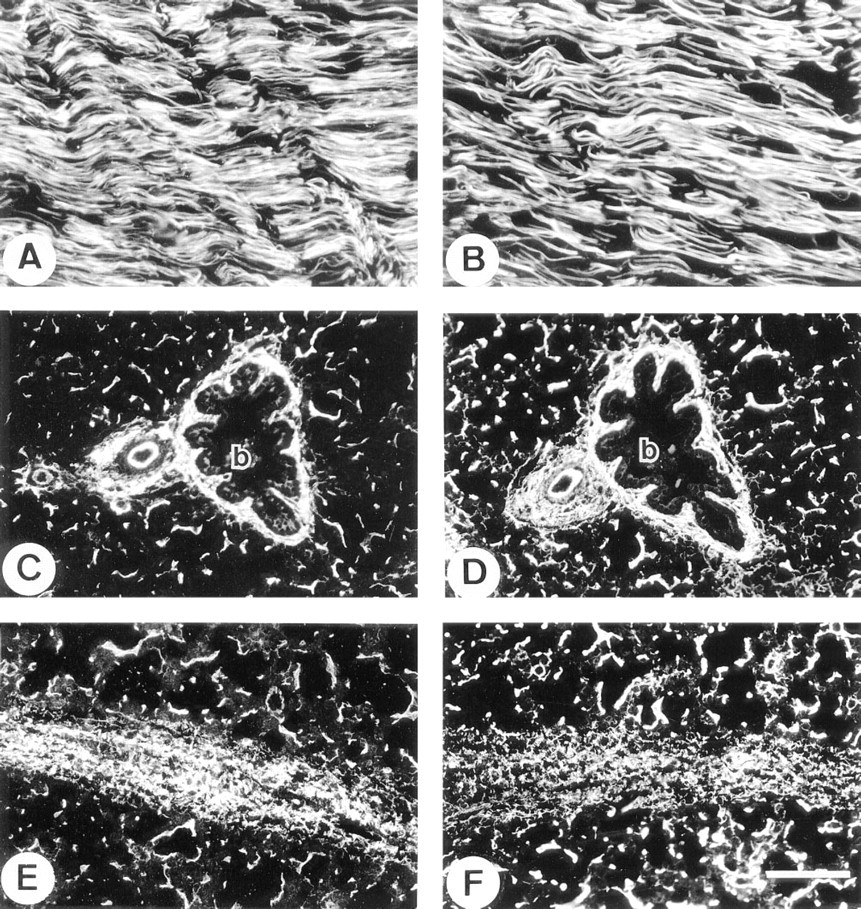
Immunofluorescence localization in fetal nuchal ligament
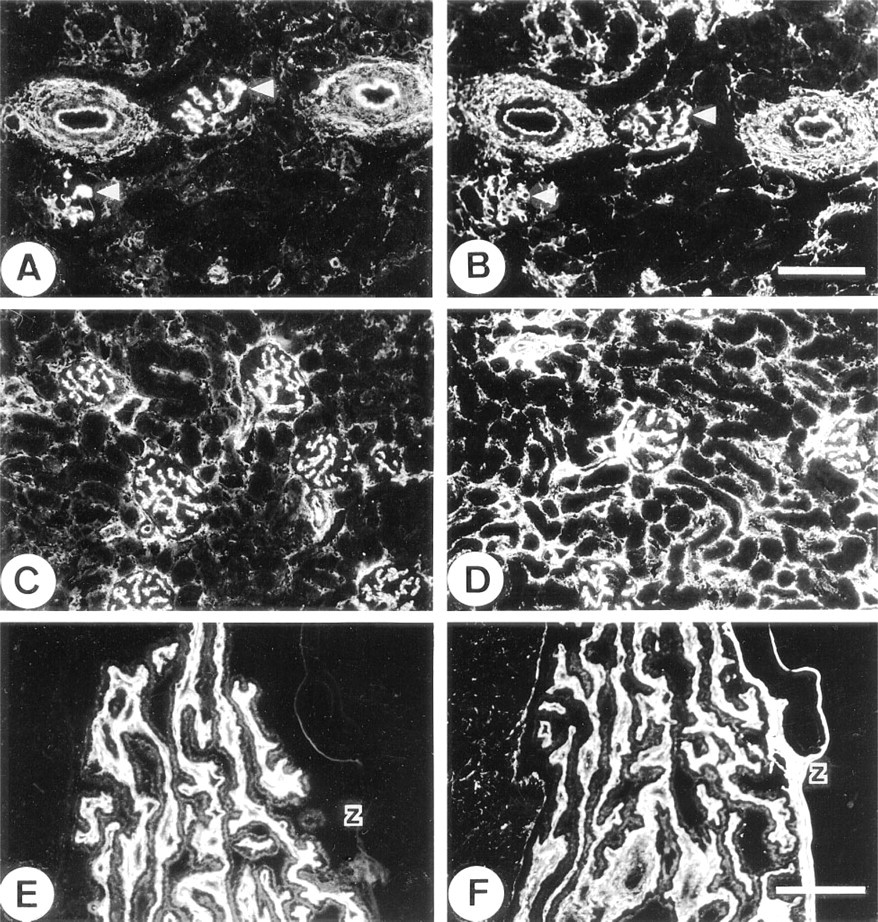
In thoracic aorta, the immunostaining for MAGP-2 was markedly different from that of MAGP-1 and fibrillin-1. The anti-MAGP-2 antibody strongly stained elastic tissue of the adventitia but showed no labeling of adjacent medial tissue, which is also rich in elastic fibers (Figure 3A). Some weak staining was evident in a few layers of elastic fibers close to the intimal region of the vessel (Figure 3B). Otherwise, the media was relatively free of immunoreactive MAGP-2. In contrast, anti-MAGP-1 (Figures 3C and 3D) and antifibrillin-1 antibodies (Figures 3E and 3F) strongly stained elastic fibers throughout the vessel wall.
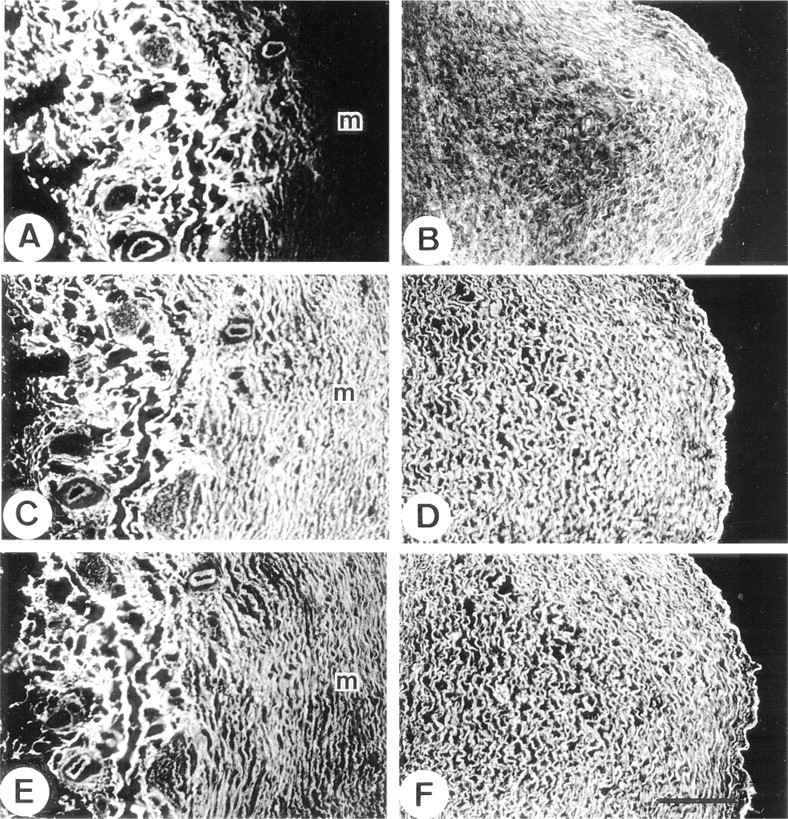
No MAGP-2 was detected in elastic ear cartilage, although the glycoprotein was strongly immunolocalized in the elastic fibers of the perichondrium and the surrounding deep dermal tissue (Figure 4A). In contrast, antibodies to MAGP-1 and fibrillin-1 strongly stained the outer margins of the cartilage matrix and the pericellular environment around chondrocytes, in addition to labeling of the perichondrium (Figures 4B and 4C, respectively). Interestingly, the major elastic fibers of the cartilage matrix did not stain for any of the above microfibril-associated proteins even when the tissue had been pretreated with hyaluronidase to enhance antibody penetration. The anti-elastin antibody strongly localized to these fibers, indicating that antibodies were able to penetrate the pretreated tissue (Figure 4D).
Immunofluorescence localization in fetal thoracic aorta.
Immunofluorescence localization in fetal ear cartilage.
In skin, the anti-MAGP-2 antibody stained the elastic fibers surrounding hair follicles but showed little staining of the dermal microfibrillar bundles close to the dermal/epidermal junction (Figure 5A). In contrast, the anti-MAGP-1 antibody strongly stained the microfibrils close to the epidermis as well as the elastic fibers surrounding the hair follicles (Figure 5B). The elastic fibers of the deep dermis were the structures in the skin that most strongly labeled with anti-MAGP-2 antibody (Figure 5C), although these structures also stained brightly with antibodies to MAGP-1 (Figure 5D), fibrillin-1 (not shown), and elastin (not shown).
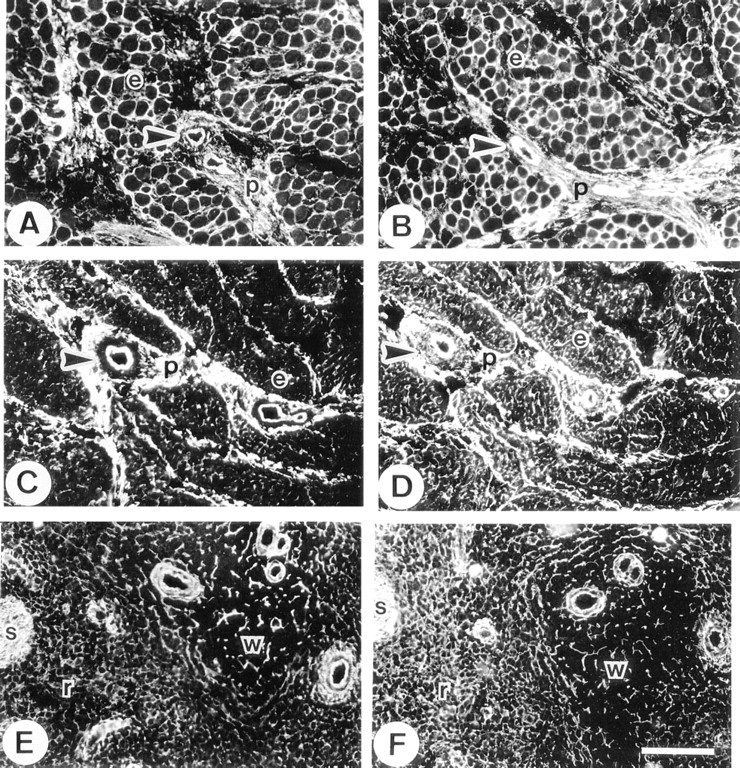
Other Tissues. In skeletal muscle, MAGP-2 was identified throughout the endomysium and perimysium and in the walls of blood vessels (Figure 6A). This distribution was indistinguishable from those of MAGP-1 (Figure 6B) and fibrillin-1 (not shown). In myocardium, MAGP-2 showed intense immunolocalization in the perimysium and intermittent localization throughout the endomysium. In the perimysium, the internal elastic lamina and adventitial layers of arterioles were also strongly stained (Figure 6C). The anti-MAGP-1 antibody also showed strong perimysial staining but appeared to localize more extensively in the endomysium than MAGP-2. In arterioles of the perimysium, MAGP-1 was detected throughout the vessel wall, including the medial layer (Figure 6D). In spleen, MAGP-2 was identified as a reticular network throughout the red and white pulp and in association with elastic fibers of the fibrous septa. The walls of small arterioles were also prominently immunostained (Figure 6E).
Immunofluorescence localization in fetal skin.
This distribution pattern was indistinguishable from that of MAGP-1, although anti-MAGP-1 antibody appeared to localize more strongly to the reticular fibers of the red pulp relative to the other immunostaining structures. (Figure 6F).
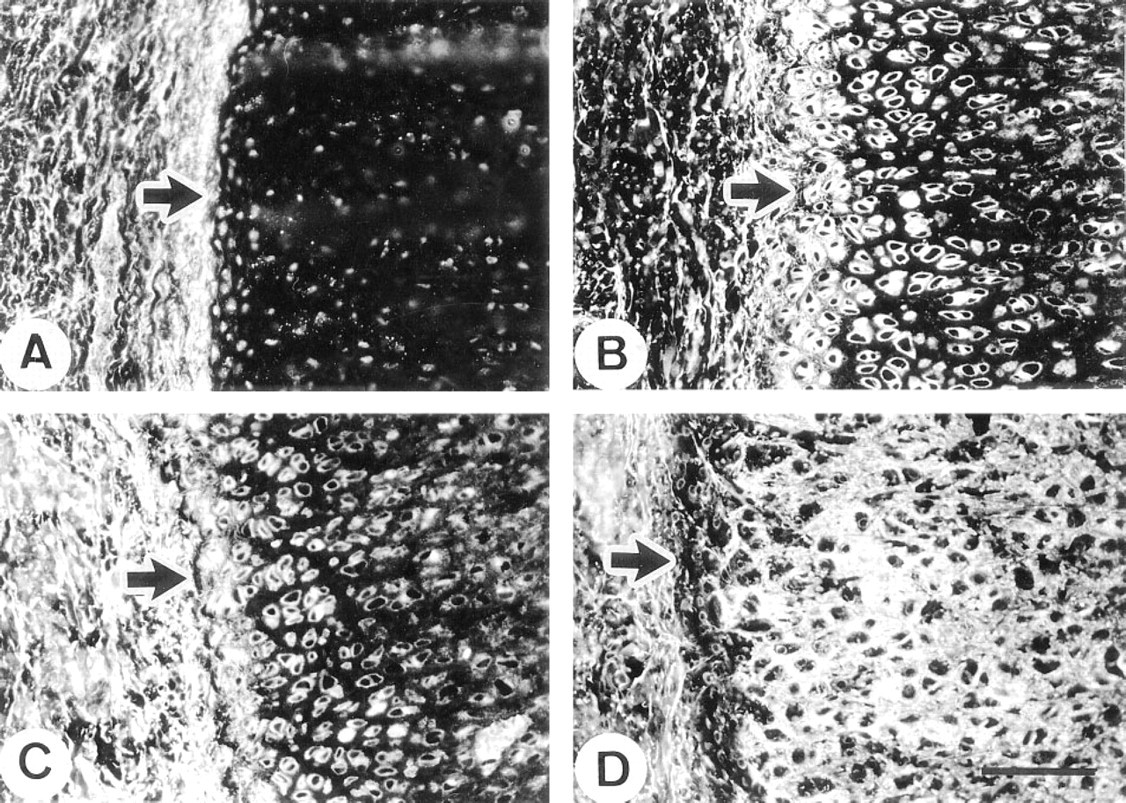
In fetal kidney, the anti-MAGP-2 antibody was very strongly localized to the glomerular mesangium. The antibody also stained the walls of arterioles, but little staining was evident in peritubular connective tissue (Figure 7A). In contrast, the anti-MAGP-1 antibody stained the mesangium, the blood vessels, and peritubular regions with equally strong intensity (Figure 7B). In adult kidney, the anti-MAGP-2 antibody again localized strongly to the mesangial tissue and, to a lesser extent, to blood vessels. Some weak intermittent staining around tubules was also detected (Figure 7C). Again, in contrast, the anti-MAGP-1 antibody localized strongly to the mesangium, blood vessels, and as a continuous peritubular network (Figure 7D). Fibrillin- 1 immunostaining patterns were similar to those of MAGP-1 in kidney (not shown). In the adult eye, anti-MAGP-2 peptide antibody did not stain the suspensory ligament of the lens (ciliary zonule), although strong staining of the elastic fibers of the adjacent ciliary body was evident (Figure 7E). In contrast, MAGP-1 was strongly immunolocalized in association with both the zonule and the elastic fibers of the ciliary body (Figure 7F).
Immunofluorescence localization in fetal muscles and spleen.
Immunoelectron Microscopy
Immunoelectron microscopy was used to confirm that the anti-MAGP-2 peptide antibody was specifically localizing to fibrillin-containing microfibrils in tissues that showed positive staining with immunofluorescence. Unfortunately, the reactive epitopes appeared to be sensitive to the fixation and embedding processes so that the postembedding labeling method could not be used. A pre-embedding labeling method was developed that enabled antibody binding to be detected by protein A-gold particles. Because the gold particles were too large to penetrate far into the tissue, labeling was confined to structures at the surface of tissue blocks. Nevertheless, very specific localization of the antibody to fibrillin-containing microfibrils was obtained in a range of tissues. Examples are shown in Figure 8. In fetal nuchal ligament, the anti-MAGP-2 antibody localized only to the elastin-associated microfibrils and not to the “amorphous” elastin core of developing elastic fibers. The antibody showed no localization to collagen fibrils or their associated fine filaments (Figure 8A). Control blocks incubated with preimmune rabbit IgG instead of the affinity-purified anti-MAGP-2 antibody, showed no immunogold labeling of the microfibrils (Figure 8B). Similarly, in the deep dermal layer of fetal skin the antibody localized specifically to bundles of fibrillin-containing microfibrils. No labeling of other matrix structures was observed (Figure 8C). Specific immunogold labeling of fibrillin-containing microfibrils was also confirmed in the mesangium of fetal kidney (Figure 8D), the perimysium of skeletal muscle (Figure 8E), and the adventitial layer of developing thoracic aorta (Figure 8F). Although the pre-embedding labeling method was suboptimal for preservation of cellular structures, it was evident (as shown in Figures 8D and 8E) that the fibrillincontaining microfibrils immunostaining for MAGP-2 are often in close association with cell surfaces. Control sections of skin, aorta, kidney, and muscle, incubated with IgG from preimmune rabbit serum instead of primary antibody showed no labeling with immunogold particles (not shown).
Immunofluorescence localization in kidney and ocular zonule.
Steady-state mRNA Measurements
The relative steady-state mRNA levels of MAGP-2 in a range of tissues from a 210-day-old fetal bovine were estimated by northern blotting of total RNA (Figure 9A). The tissue exhibiting the highest level of MAGP-2 mRNA expression, by far, was nuchal ligament. Significant expression was also detected in lung, heart, skeletal muscle, skin, and Achilles tendon. Expression was also detected, at lower levels, in aorta and cartilage (including perichondrium) and at trace levels in kidney, spleen, and liver. The above pattern of tissue expression was distinct from that of MAGP-1 (Figure 9B). Highest levels of MAGP-1 mRNA expression were found in lung and nuchal ligament, followed by kidney, skin, aorta, and tendon. Low levels of mRNA were also identified in skeletal muscle, heart, spleen, and cartilage. A trace of MAGP-1 mRNA was identified in liver. Relative to expression in other tissues, MAGP-2 appeared to have higher mRNA levels in nuchal ligament, heart, and skeletal muscle, and lower levels in aorta and kidney, than MAGP-1.
To determine if expression of MAGP-2 changed during the development of an elastic tissue, the relative steady-state levels of its mRNA in nuchal ligament were measured in tissue obtained from fetuses ranging in age from 110 days to term (270 days). The values obtained from densitometric scanning were plotted relative to the value obtained from the ligament of an adult (2-year-old) bovine. The MAGP-2 mRNA expression profile was compared to those for MAGP-1 mRNA and elastin mRNA (Figure 10). From the graph, expression of MAGP-2 mRNA appeared to be at its highest between 160 days and 210 days of fetal development, when it was at least sixfold higher than in the adult tissue. This period corresponds, in the nuchal ligament, to the early stages of elastinogenesis, which peaks around birth. Interestingly, the expression profile for MAGP-1 did not correlate with those of MAGP-2 and elastin. Expression levels of MAGP-1 appeared to have peaked much earlier in development of the tissue, although steady-state MAGP-1 mRNA levels remained relatively high throughout development and in mature ligament. It should be noted that the arbitrary mRNA expression values cannot be used to measure comparative expression levels of different mRNAs.
Discussion
Fibrillin-containing microfibrils appear to perform a range of functions within the extracellular matrix, depending on the tissue and its stage of development. These functions include providing a scaffold for the deposition and alignment of the elastin precursor, tropoelastin, during elastic fiber assembly, acting as anchoring filaments for elastic fibers and other structures of the extracellular matrix, and providing a substrate for cellular interaction with the surrounding matrix (Cleary and Gibson 1983,1996; Sakamoto et al. 1996; Finnis and Gibson 1997). Only recently have clues appeared as to how the structure of the microfibrils may vary to accommodate these diverse functions. Earlier, the evidence suggested that fibrillin-containing microfibrils ubiquitously consisted of fibrillin-1 in combination with MAGP-1 and other proteins. However the cloning and characterization of fibrillin-2 indicated that these microfibrils are heterogeneous in structure (Zhang et al. 1994,1995). The two fibrillins have markedly different expression patterns during development, and it has been suggested that fibrillin-1 may provide force-bearing structural support to tissues, whereas fibrillin-2 is predominantly associated with the early stages of elastic fiber assembly (Zhang et al. 1995). However, the same authors state that the proposed function for each fibrillin should be considered a distinguishing attribute rather than an exclusive feature.
Further evidence for structural diversity of fibrillin-containing microfibrils has come from the cloning of MAGP-2, which established the molecule as a distinct microfibril-associated protein. MAGP-2 was found to have close sequence similarity to MAGP-1 in a central region of 60 amino acids, where the spacing of seven cysteine residues was identical in both proteins. Elsewhere, the MAGP-2 molecule is rich in serine and threonine residues and lacks the proline-, glutamine-, and tyrosine-rich sequences characteristic of the aminoterminal domain of MAGP-1 (Gibson et al. 1996). MAGP-1, unlike MAGP-2, binds in vitro to the elastin precursor tropoelastin and to Type VI collagen (Brown-Augsburger et al. 1994,1996; Finnis and Gibson 1997). It is evident that MAGP-1 plays an important role in the interaction of the fibrillin-containing microfibrils with elastin during elastic fiber formation (Brown-Augsburger et al. 1996). MAGP-1 also appears to mediate an interaction between fibrillin-containing microfibrils and Type VI collagen microfibrils, which may be important for anchoring elastic fibers to the structural elements of the surrounding matrix. Both of these molecular interactions appear to involve a region close to the amino terminal region of the MAGP-1 molecule (Finnis and Gibson 1997). The corresponding region of MAGP-2 contains an RGD sequence which specifically binds integrin αVβ3 (manuscript in preparation), indicating that MAGP-2 may play a role in cell-microfibril interactions. Therefore, these studies suggest that MAGP-1 and MAGP-2 are structurally divergent proteins sharing one functional motif rather than being functionally closely related molecules (Finnis and Gibson 1997). This idea is supported by recent data showing that the genes for MAGP-1 and MAGP-2 are very different in structure, with homology being confined to two conserved exons (manuscript in preparation). These exons encode a cysteine-rich motif that may be involved in the covalent binding of each protein to the microfibril.
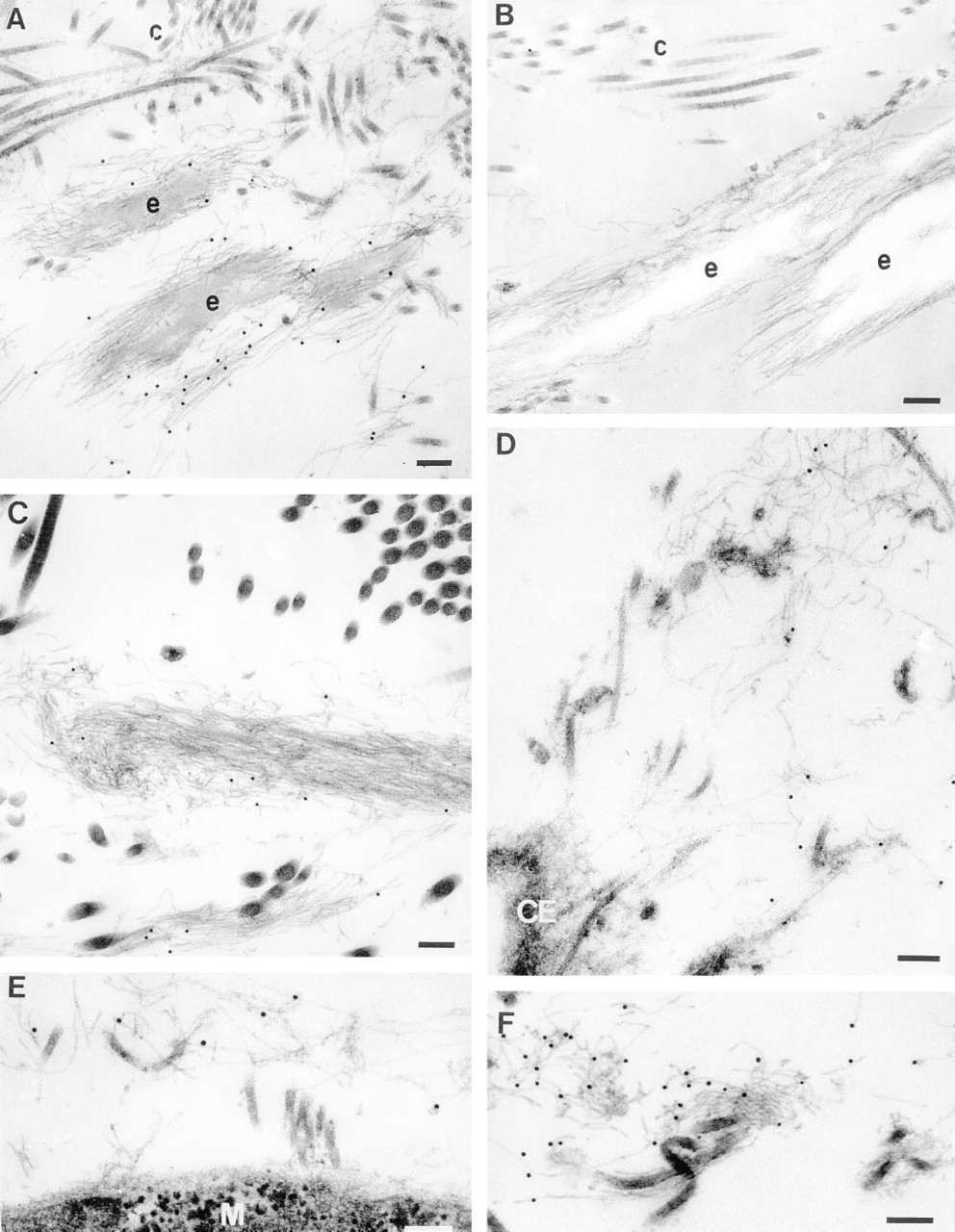
Ultrastructural localization of MAGP-2 in tissues of a 220-day-old fetal calf.
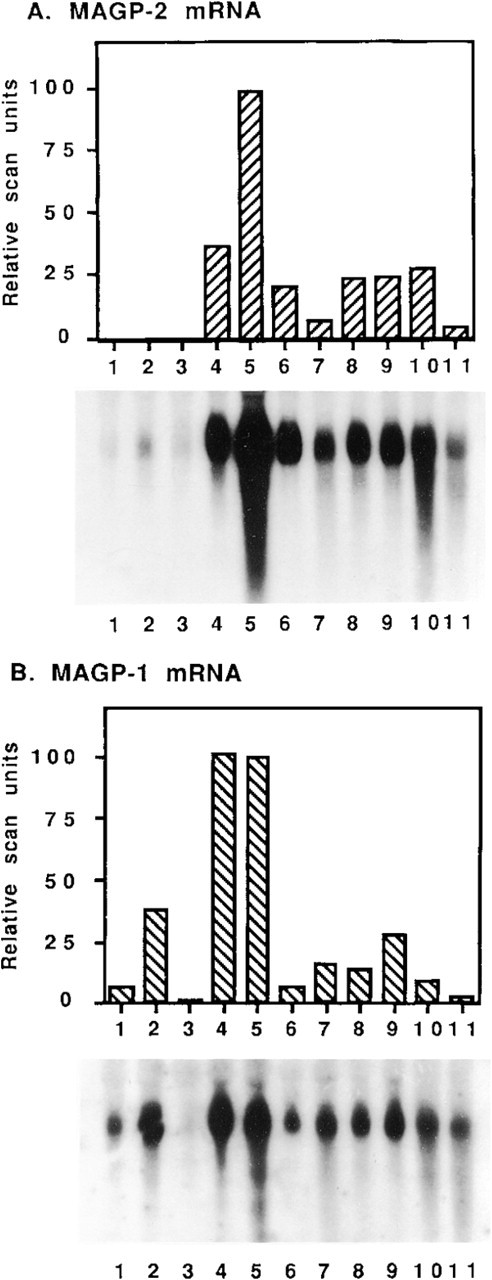
Measurement of relative steady-state mRNA levels for MAGP-2 and MAGP-1 in tissues from a 210-day-old fetal bovine. Total RNA (10 μg) from each tissue was analyzed by northern blotting using 32P-labeled cDNA probes for
The present study provides further evidence for functional diversity of the MAGPs. A range of tissues from a 210-day-old fetal calf and some tissues from an adult bovine were immunostained with an anti-MAGP-2 peptide antibody and the patterns were compared to those for MAGP-1, fibrillin-1, and elastin. The tissues were also treated with a previously described polyclonal antibody raised against the whole MAGP-2 molecule (Gibson et al. 1996) which gave staining patterns identical to those of the anti-MAGP-2 peptide antibody, confirming its specificity (not shown). The immunolocalization patterns of fibrillin-1 in many of the tissues are not shown but have recently been described by Gibson et al. (1997). The fibrillin-1 staining patterns appear to be indistinguishable from those for MAGP-1 shown here. In general, the anti-MAGP-2 antibody localized in regions of tissues that also stained with antibodies to both MAGP-1 and fibrillin-1 (Figures 2–7). No localization of MAGP-2 with other structures was observed, suggesting that MAGP-2 was also specifically associated with these microfibrils. In nuchal ligament, skeletal muscle, and spleen, the tissue distributions of MAGP-2 and MAGP-1 were indistinguishable from one another. Some subtle differences in localizations between the two proteins were detected in lung, cartilage, skin, and heart. Major differences in the distributions of MAGP-2 and MAGP-1 were evident in fetal aorta, fetal and mature kidney, and mature ocular zonule. In aorta, MAGP-2 was detected in the adventitia and the inner few layers of elastic lamellae close to the lumen but was not found in association with the elastic fibers of the thick medial layer, which immunostained strongly for MAGP-1 and fibrillin-1. In addition, MAGP-2 was not detected in the muscular layers of small arteries and arterioles in a range of tissues. In fetal and adult kidney, MAGP-2 was detected, with strong signal, in the glomerular mesangium and in blood vessels, but showed little localization around tubules, unlike MAGP-1, which was strongly localized in this latter region. Interestingly, the ocular zonule of the adult eye showed no immunostaining for MAGP-2, in contrast to its strong staining for MAGP-1. The fibrillin-containing microfibrils are the only discernible structures in the acellular zonule, where fibrillin-1 is considered to be the only major fibrillin present (Zhang et al. 1995). Immunoelectron microscopy confirmed that MAGP-2 was specifically associated with fibrillin-containing microfibrils in those regions of tissues that stained positively by immunofluorescence. No staining of other matrix structures was observed (Figure 8).
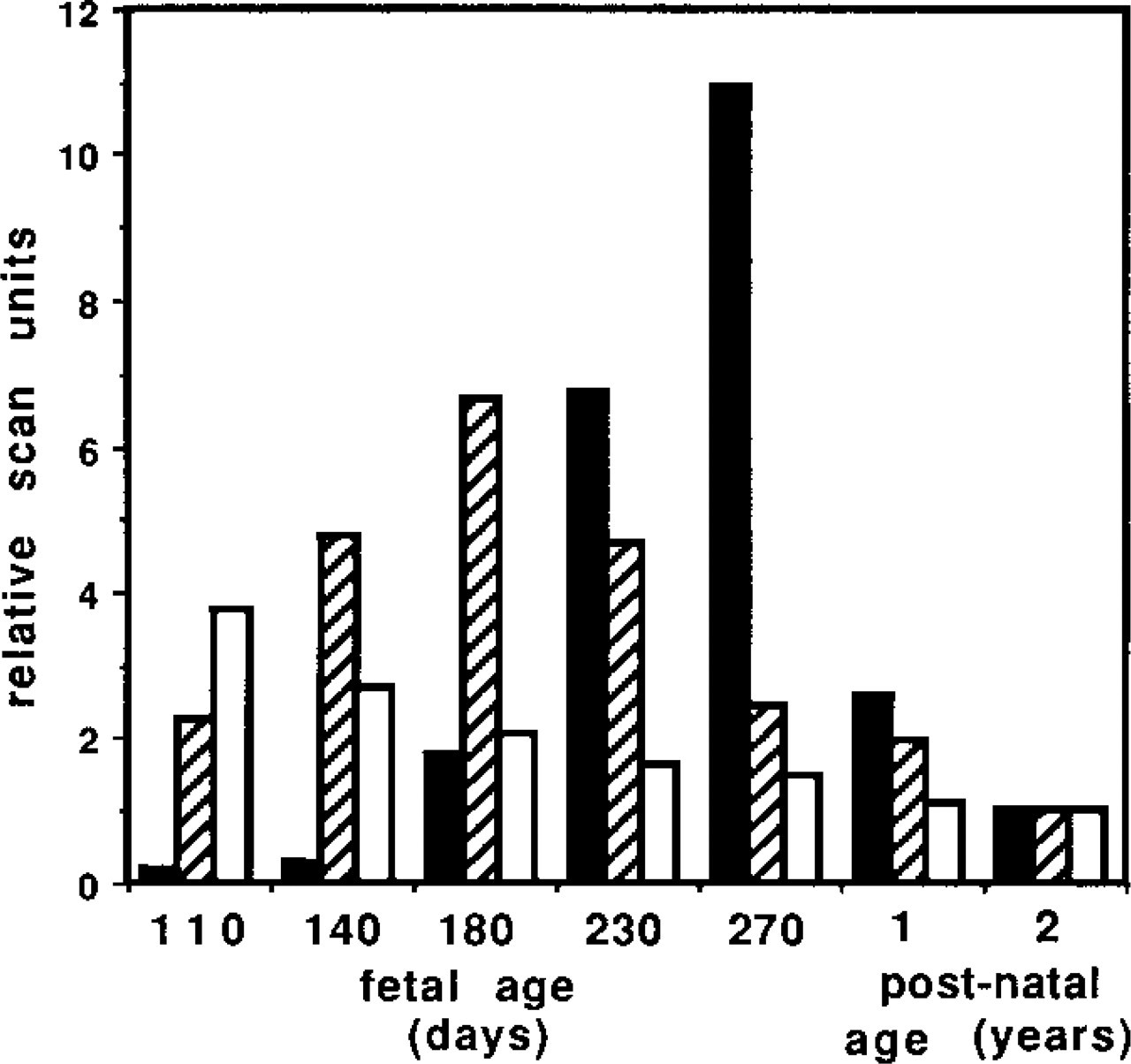
Measurement of relative steady-state mRNA levels during development of nuchal ligament. Total RNA (10 μg) from several stages of fetal and post-natal development was analyzed by northern blotting with 32P-labeled cDNAs for MAGP-2, MAGP-1, and elastin. Each signal on the autoradiographs was scanned densitometrically and normalized to GAPDH mRNA. The steady-state level for each mRNA was expressed as arbitrary scan units relative to the 2-year postnatal value (assigned value 5 = unit). Black columns, elastin mRNA; cross-hatched columns, MAGP-2 mRNA; white columns, MAGP-1 mRNA.
The reason(s) for the differential staining patterns of the two MAGPs is likely to reflect differences in function between the two proteins. In contrast to the proposed structural roles for MAGP-1, MAGP-2 may be involved in the interaction of microfibrils with particular cells, via an integrin surface receptor, in tissues such as nuchal ligament and lung. It may be that in some tissues in which the cells become separated from the microfibrils by basement membranes, e.g., dermal epithelial cells and vascular smooth muscle cells, or in acellular tissues, e.g., zonule, there is no direct interaction of cells and adjacent microfibrils. An interaction of the cell with MAGP-2 may not be required in such circumstances and therefore the protein is not found in association with these microfibrils.
The close correlation of immunolocalization patterns of MAGP-1 and fibrillin-1 in developing and mature tissues, and the apparent absence of both proteins from the elastic fibers of developing ear cartilage, are consistent with a specific association of MAGP-1 with fibrillin-1-containing microfibrils. MAGP-2, like fibrillin-2, has more restricted expression patterns during tissue development. We have previously suggested that MAGP-2 may be associated exclusively with fibrillin-2-containing microfibrils because there appeared to be some correlation of expression patterns in tissues such as kidney, lung, and ocular zonule (Gibson et al. 1996). To answer this question, we raised antibodies using synthetic peptides corresponding to the unique glycine-rich “C-region” of fibrillin-2 (see Zhang et al. 1994). Thus far we have been unable to purify these antibodies free of crossreactivity to elastin, which has a similar glycine-rich structure, and therefore the question remains unresolved. However, MAGP-2 appears to be absent from the elastic tissue of ear cartilage and the media of elastic blood vessels, both of which tissues are apparently rich in fibrillin-2. This suggests that a ubiquitous co-distribution of the two proteins does not occur.
Distinctions were also found in the tissue expression patterns of mRNAs for MAGP-2 and MAGP-1 in a range of tissues from the 210-day-old fetus, as determined by Northern blotting (Figure 9), although absolute quantitation of each mRNA was not possible by this method. In terms of its overall tissue expression profile, MAGP-2 mRNA appeared to show higher expression in nuchal ligament, and possibly in heart and skeletal muscle, than MAGP-1 mRNA, which appeared to have higher expression in aorta and kidney. The higher expression of MAGP-1 mRNA in the latter two tissues correlates well with the immunofluorescence findings. However, relatively high MAGP-2 expression in heart and muscle was not apparent in the immunofluorescence study. Expression differences between MAGP-2 and MAGP-1 were also found during development of nuchal ligament in fetal calves (Figure 10). The steady-state mRNA level for MAGP-2 peaked at around 180 days, which correlates with the period of onset of elastinogenesis in this tissue. This transient increase in MAGP-2 expression appeared to match the similar elevation in the expression of fibrillin-1 and fibrillin-2 previously observed at this stage of ligament development by Mariencheck et al. (1995). In agreement with the above study, MAGP-1 mRNA expression, although high throughout ligament development, showed no transient elevation during the elastinogenic period (Figure 10). The findings suggest that MAGP-2 may have an important function during the early stages of elastinogenesis, when tropoelastin molecules are being deposited and aligned on the microfibril scaffold close to the cell surface. However, it should be noted that significant levels of MAGP-2 mRNA were found in relatively mature ligament tissue (from a 2-year adult bovine), suggesting that MAGP-2 may also have a more enduring role in microfibril biology.
Overall, the present study shows that the immunolocalization and expression patterns of MAGP-2 are distinct from those of other microfibrillar components. This is consistent with a unique role for MAGP-2 in modulation of the function of fibrillin-containing microfibrils in certain tissue-specific matrices. This function may involve mediation of microfibril interactions with cell surfaces and matrix molecules at particular stages of development and differentiation, including early elastinogenesis.
Footnotes
Acknowledgements
Supported by the National Health and Medical Research Council of Australia.
We are indebted to Denise Yates for skilled technical assistance.