
Abstract
Evidence suggests that angiotensin type 2 receptor (AT2R) and angiotensin-converting enzyme 2 (ACE2) play a protective role in atherogenesis. These factors have not been identified in rabbit atherosclerotic plaques. Our goal was to localize ACE2 and AT2R in rabbit atherosclerotic tissues, and determine which cell types express these factors. New Zealand White rabbits were fed either a control diet or a 0.5% cholesterol diet (n=8/group) for 12 weeks. Paraffin-fixed thoracic aorta were serially sectioned and processed for immunohistochemistry using commercially available antibodies to ACE2, AT2R, RAM 11 (to identify macrophages), and α smooth muscle cell actin (αSMC) to identify smooth muscle cells and myofibroblasts. AT2R immunoreactivity, but not ACE2 immunoreactivity, was clearly present in endothelia overlying normal wall. However, both AT2R and ACE2 immunoreactivity were clearly present in endothelia overlying neo-intima formation and atherosclerotic plaques. Within plaques, both AT2R and ACE2 immunoreactivity were observed in macrophages and αSMC actin-positive cells. Examination of serial sections showed that the majority of cells were both ACE2- and AT2R-positive. Macrophages and αSMC actin-positive cells produce ACE2 and the AT2R in atherosclerotic plaques. Determining a role for these factors in the control of atherosclerosis will require additional studies.
T
Angiotensin-converting enzyme 2 (ACE2) is a newly described enzyme that can break down angiotensin I into Ang (1–9) and Ang II into Ang (1–7) (Boehm and Nabel, 2002). Although Ang (1–7) may in itself be atheroprotective (Osei et al. 1993; Ferrario 1998), the conversion of AngII to Ang (1–7) via ACE2 can be atheroprotective simply because it removes AngII from the atherogenic milieu.
ACE and AT1R expression have been previously reported in rabbit atherosclerotic tissues (Schuh et al. 1993). Furthermore, the inhibition of either ACE activity (Candido et al. 2002) or inhibition of AT1R activation (Li et al. 1999) by selective pharmacotherapy greatly reduces atherosclerosis formation in the rabbit model of atherosclerosis, indicating a detrimental role for ACE and the AT1R in atherogenesis.
Thus it is hypothesized that in atherogenesis, ACE, ACE2, AT1R, and the AT2R are all expressed, and that the inhibition of ACE or AT1R would leave ACE2 and AT2R unopposed to impair atherogenesis. However, if credence is to be given to this theory, high levels of ACE2 and AT2R should be expressed in atherosclerotic tissues, because ACE and AT1R levels have already been shown. To this end, immunohistochemistry for ACE2 and AT2R were performed on rabbit atherosclerotic tissues, and specific cell markers were used to identify macrophages and α smooth muscle cell actin (αSMC)-positive cells (smooth muscle cells and myofibroblasts).
Materials and Methods
Male New Zealand White rabbits at 3 months of age were randomly allocated into two groups of eight rabbits and fed either a normal rabbit chow diet supplemented with 0.5% cholesterol (Chol) or a normal rabbit chow diet (Con). The animals were housed in individual cages and maintained at a constant temperature of ∼21C. Food and water was supplied ad libitum. The experiments were approved by the Monash University, Department of Anatomy Ethics Committee, and were carried out according to the National Health and Medical Research Council “Australian Code of Practice for the Care and Use of Animals for Scientific Purposes” (6th Edition, 1997).
Immunohistochemistry
Thoracic aortae were processed for paraffin. Paraffin blocks were cut at 5 μm and collected on gelatin-coated slides. Slides were dewaxed, rehydrated, and incubated with blocking serum (1% donkey serum diluted in 10 mM Tris HCl, pH 7.3). Primary antibodies were diluted (AT2R and ACE2, 1:500 dilution; Santa Cruz Biotechnologies, Santa Cruz, CA) and the slides incubated overnight. Antibody absorbtion was used as a negative control for AT2R and ACE2. Slides were then rinsed in 10 mM Tris HCl, pH 7.3, and a biotinylated donkey anti-goat polyclonal antibody (Chemicon International; Temecula, CA) was diluted 1:100 in antibody diluent (Dakocytomation, Australia) and the slides incubated for one hour. Slides were then rinsed in 10mM Tris HCl, (pH 7.3) and incubated with an ultrasensitive streptavidin-peroxidase polymer (Sigma-Aldrich, Australia) diluted 1:100 in antibody diluent (Dakocytomation; Carpinteria, CA) for 1 hr. Diaminobenzidine was used as the substrate.
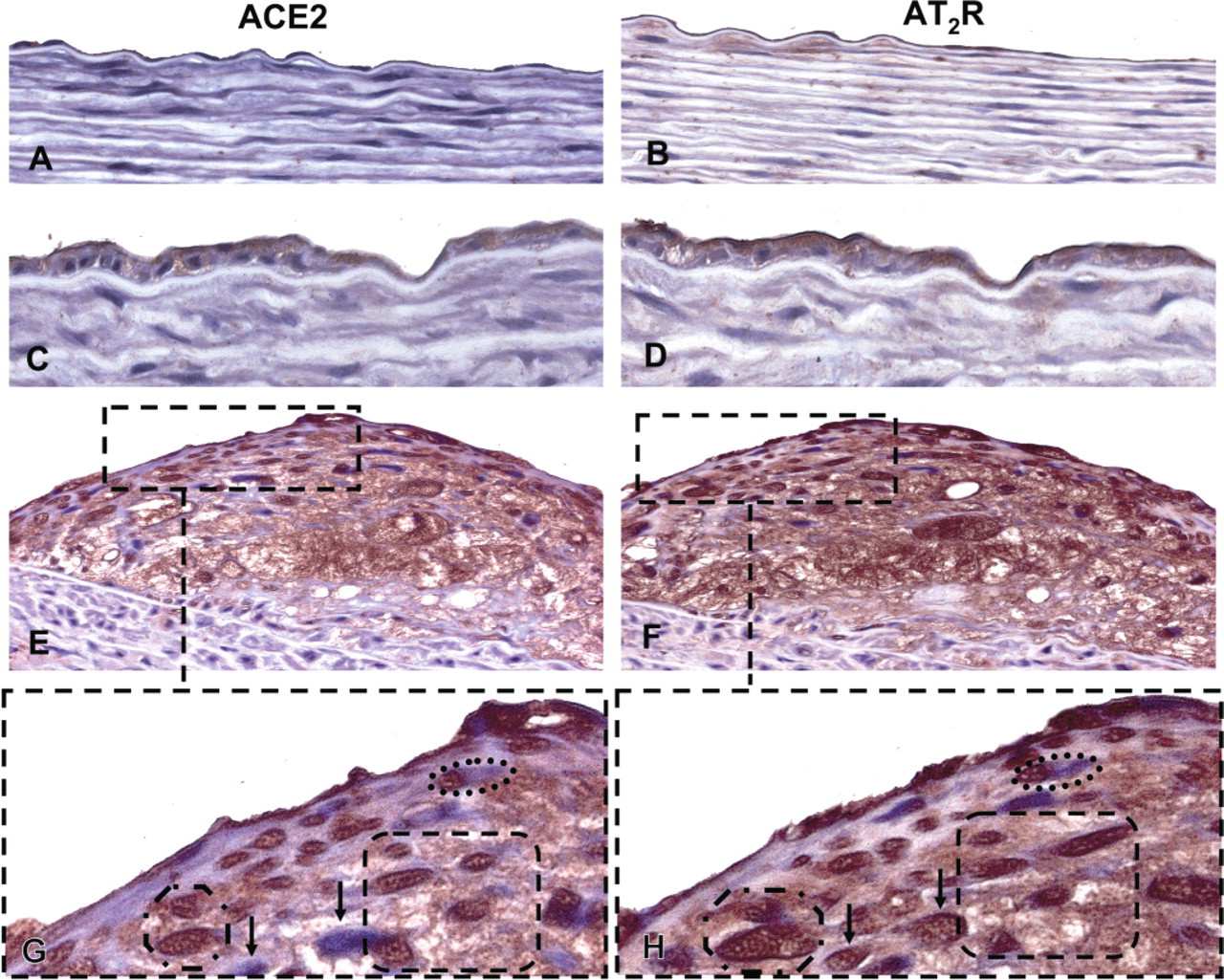
Photomicrographs of serial adjacent sections of rabbit thoracic aorta demonstrating angiotensin-converting enzyme 2 (ACE2) and angiotensin II type 2 receptor (AT2R) immunoreactivity. ACE2 immunoreactivity was not clearly evident in normal thoracic aorta endothelia (
Monoclonal antibodies to RAM-11 and αSMC actin (Dakocyotmation) were used to identify macrophages and smooth muscle cells/myofibroblasts. In this case, immunohistochemistry was performed using the Envision Kit (Dakocytomation) as previously described (Zulli et al. 2003, 2005).
Results
AT2R, but not ACE2, immunoreactivity, was clearly visible on the endothelial layer overlying normal vessel wall (Figure 1B and 1A, respectively). However, both proteins were present on the endothelial cell layer overlying neo-intima and atherosclerotic lesions (Figure 1D and 1C, respectively). Both ACE2 and the AT2R were present in oval and spindle-shaped cells within plaques, as well as in macrophages (RAM-11 positive) both overlying and within atherosclerotic plaques (Figures 11-E1). A magnification of Figures 1E and 1F (dashed box) clearly shows that the majority of cells in plaques were positive for both proteins (dashed circles); however, there were some cells that were only positive for AT2R, but not ACE2 (Figures 1G and 1H depicted by arrows).
To determine whether macrophages or αSMC actin-positive cells expressed both ACE2 and AT2R, we used serially sectioned adjacent sections and performed immunohistochemistry for all four antibodies. We observed that a high proportion of macrophages and SMC actin-positive cells within atherosclerotic plaques expressed both ACE2 and AT2R, depicted by circles (Figure 2).
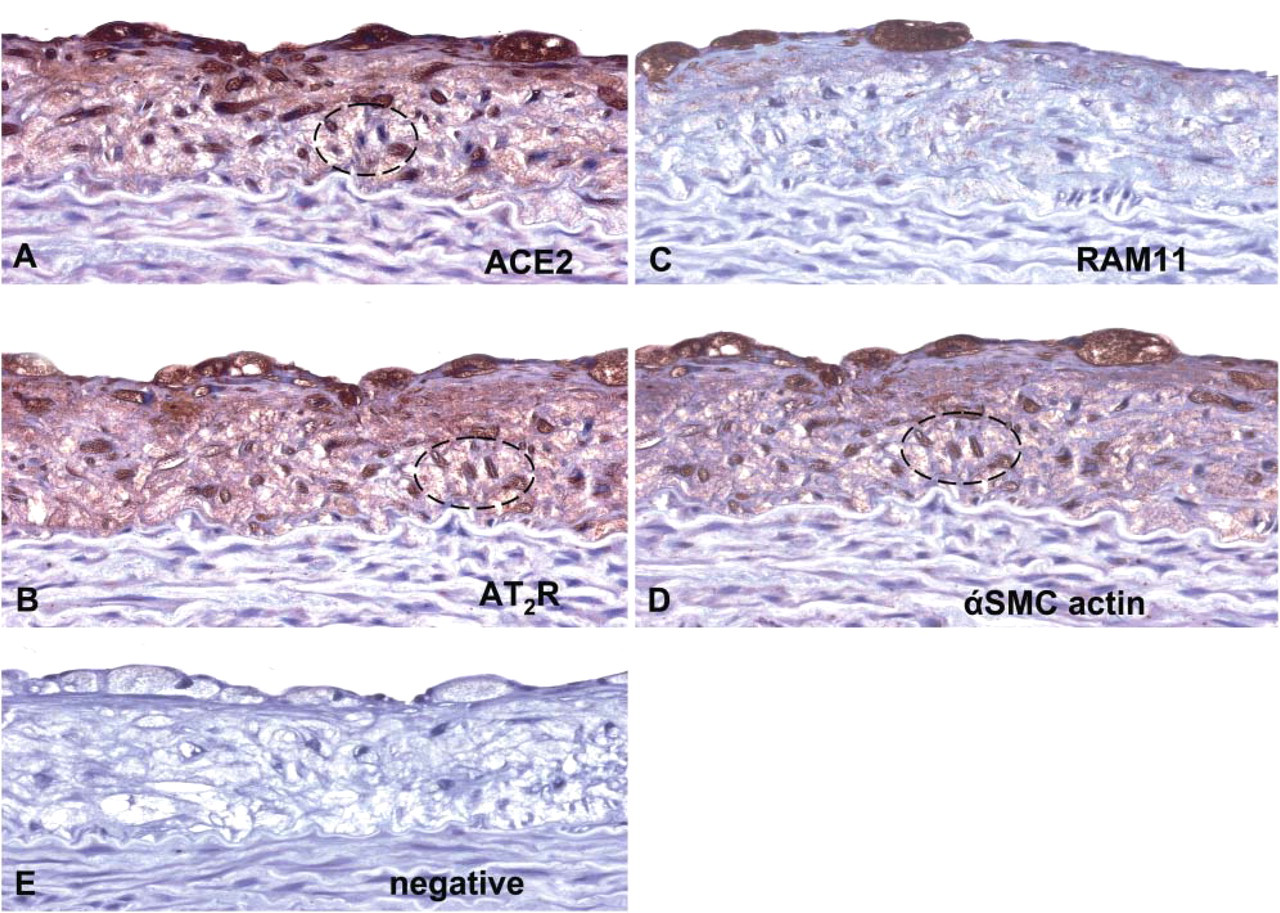
Photomicrographs of serial adjacent sections of rabbit thoracic aorta demonstrating (
Discussion
We have demonstrated by immunohistochemistry that macrophages and αSMC actin-positive cells within atherosclerotic plaques express high levels of ACE2 and AT2R proteins. In combination with results published previously by other authors, the results presented in this study add to the growing body of evidence that suggests that all the factors of the renin angiotensin system can be coexpressed in atherogenesis.
It has been previously shown that inhibition of ACE (Schuh et al. 1993) or AT1R (Candido et al. 2002) with selective pharmacotherapy greatly reduces atherosclerosis. Although the mechanisms of decreased atherogenesis in this model has been suggested to be because of a decrease in AngII formation and AT1R inhibition, other atheroprotective mechanisms may be stimulated. For example, ACE inhibitors do not inhibit ACE2 activity. Thus it could be suggested that ACE inhibition may not only decrease AngII formation, but also leave angiotensin I to form Ang(1–9) via ACE2, and as we have shown, ACE2 is highly expressed in atherosclerotic plaques. Although there is no direct evidence to suggest that Ang(1–9) is atheroprotective, Ang(1–9) is converted to Ang(1–7), which has antiatherogenic properties (Osei et al. 1993; Ferrario 1998).
Another possible atheroprotective mechanism of AT1R blockade could be that “free” AngII could bind to the AT2R. Stimulation of the AT2R has been shown to inhibit neo-intimal formation (Janiak et al. 1992), decrease smooth muscle cell proliferation (Nakajima et al. 1995) and migration (Chassagne et al. 2002), and inhibit fibroblast proliferation (Tsuzuki et al. 1996). Moreover, AngII can be converted to Ang (1–7) by ACE2 (Boehm and Nabel 2002). Thus we suggest that in the development or regression of atherosclerosis, blockade of the proatherogenic actions of ACE or the AT1R could unmask the inhibitory actions of ACE2 and the AT2R.
In conclusion, ACE2 and AT2R were identified in macrophages and smooth muscle cells/fibroblast within atherosclerotic plaques and the majority of cells expressed both proteins. Although the role of ACE2 and the AT2R in atherogenesis is suggested to be atheroprotective, the specific role of these factors in atherogenesis warrants further study.
Footnotes
Acknowledgements
This project has been supported by the Austin Hospital Medical Research Foundation.