
Abstract
The HNK-1 carbohydrate epitope is a 3-sulfo-glucuronyl residue attached to lactosamine structures on glycoproteins, proteoglycans, or glycolipids mostly expressed in the nervous system. Here, using monoclonal antibodies against the sulfated HNK-1 carbohydrate epitope, we first examined its distribution in developing and adult kidneys, then its expression in kidneys with tubular necrosis and renal neoplasms. This HNK-1 epitope was expressed in the human, rabbit, and rat, but not mouse kidney. It was detected within a subset of epithelial cells in the renal vesicle and in comma- and S-shaped bodies during early stages of nephrogenesis. In ureteral bud derivatives, the epitope was present transiently in the area where the collecting duct fused with the nephron. In the adult kidney, expression of the HNK-1 epitope became mainly restricted to the thin ascending loop of Henle where this epitope was carried by heparan- and chondro-proteoglycan. In pathological conditions, HNK-1 epitope expression increased dramatically in proximal epithelial tubule cells in kidneys with acute tubular necrosis. In tumors, the HNK-1 epitope was expressed in the epithelial component of nephroblastomas and in a subgroup of papillary renal cell carcinomas. These data suggest that molecules carrying the sulfated HNK-1 carbohydrate epitope may play an important role in critical stages of renal development and in the physiology of thin ascending loop of Henle.
T
Kidney organogenesis involves reciprocal inductive interactions between the ureteric bud epithelium and metanephric mesenchyme (Kuure et al. 2000; Vainio and Lin 2002; Mori et al. 2003). The tips of the ureteric bud induce nephrogenic mesenchyme to form condensates followed by pretubular aggregates, which undergo mesenchyme to epithelium transition and progress into polarized epithelia of renal vesicles and comma-and S-shaped bodies. Morphogenesis and patterning of the epithelial structures lead to the formation and functional maturation of distal and proximal tubules and the glomerulus. On the other side, the most distal segment of the primitive nephron becomes connected to the ureteric bud epithelium. The molecular mechanism by which the S-shaped body undergoes segmentation and connection with the ureteric bud epithelium is unknown at present. This process requires the fine-tuning of cell interactions. Cell-cell contacts and cell-matrix interactions mediated by cell surface recognition molecules play a key role in these processes (Kanwar et al. 2004). Glycosylation of cell recognition molecules is one of the important posttranslational modifications that regulates their functions.
Here we examined the sulfated HNK-1 carbohydrate epitope expression from earliest stages of nephron development to mature kidney. We then studied its expression in native kidneys with acute tubular necrosis and renal tumors of diverse histopathological origin. Our results indicate that the HNK-1 epitope is selectively expressed within a subset of epithelial cells during early nephrogenesis and becomes restricted to the thin ascending limb of loop of Henle in the adult kidney where this epitope is carried by heparan- and chondroproteoglycans. Moreover, this epitope may serve as a biomarker for proximal tubular injury.
Materials and Methods
Tissue Specimens
Human tissue specimens were retrieved from the Departments of Pathology at Tenon and Trousseau Hospitals (Paris, France). All tissues were collected after patients gave informed consent. Normal human adult renal tissue was obtained from the unaffected pole of 10 kidneys removed due to a renal cell carcinoma (RCC). Twelve normal human embryonic and fetal kidneys were obtained from therapeutic abortion material. Acute tubular necrosis was studied in 10 renal biopsies performed in grafted kidneys 8 to 13 days after transplantation. Renal tumors comprised 9 nephroblastomas with epithelial component, 91 oncocytomas, 208 adult RCCs (71 clear cell, 78 papillary, 48 chromophobe, and 11 low-grade tubular mucinous and spindle cell variants) diagnosed according to the WHO 2004 classification (Eble et al. 2004). All RCCs (tubular mucinous variant excepted) were studied using the tissue microarrays previously described (Allory et al. 2005). Mucinous tubular and spindle cell carcinomas were characteristically made of regular spindle or cuboid cells arranged in a myxoid alcian blue-positive stroma, with a distal tubule immunohistochemical profile (epithelial membrane antigen, cytokeratin 7, E-cadherin positive, and CD10 negative). All tissues were 10% (v/v) formalin fixed by immersion. Male Wistar rat, C57/BL6 mouse, and rabbit kidneys were fixed with 2% (w/v) paraformaldehyde in PBS, pH 7.4, by vascular perfusion followed by immersion in the same fixative for up to 2 hr. Human non-tumoral lymph nodes and mouse cerebellum were used as positive controls. For immunohistochemistry, all tissues were embedded in paraffin according to standard protocols. Five-μm sections were prepared and mounted on Superfrost Plus glass slides. For biochemical analysis, samples of human adult and rat kidney were also snap frozen in liquid nitrogen and conserved at −80C until use. Animal studies were performed according to INSERM Ethical Committee recommendations.
Antibodies
Primary MAbs used in this study were the anti-HNK-1 (IgM, clone VC1.1, 1:200 dilution; Sigma, St Louis, MO), the anti-L1CAM MAb272 (Debiec et al. 1998), the anti-CD34 (IgG, clone Tük 3, 1:25 dilution; DAKO, Glostrup, Denmark), the anti-E-cadherin (IgG, clone A42-C7, 1:100 dilution; Zymed, South San Francisco, CA), and the anti-cytokeratin 7 (IgG, clone OV-TL,1:250 dilution; DAKO). Polyclonal rabbit anti-sera used were specific for Tamm-Horsfall protein (1:2000 dilution) (Xuetal. 1997), aquaporin 1 (1:100 dilution; Chemicon, Temecula, CA), urea transporter type 2 (UTA2) (1:100 dilution), a kind gift from M.M. Trinh Trang Tan, INSERM Unit 665, Paris, France.
Immunohistochemistry
For HNK-1 immunostaining on human and rat renal tissues, sections were processed on an automated instrument (Ventana Nexes; Ventana Medical Systems, Paris, France) using an indirect biotin-avidin system, the Ventana Basic DAB detection kit (Ventana Medical Systems) according to manufacturer's instructions, without antigen retrieval procedure. For HNK-1 immunostaining on mouse and rabbit tissue, it was impossible to use the automated instrument because of cross-reactions of the analyzed tissue with the anti-mouse and anti-rabbit immunoglobulin cocktail present in the instrument. Thus, we used a biotinylation kit (Mouse-on-Mouse Iso-IHC Kit; InnoGenex, San Ramon, CA), and we applied prelabeled immunoglobulin complexes on the rabbit and mouse tissue and then on the alkaline phosphatase-streptavidin conjugate. The reaction product was visualized with developing solution containing blue-purple 5-bromo-4-chloro-3-indoxyl phosphate with nitroblue tetrazolium chloride (BCIP/NBT) substrate according to the manufacturer's instructions. Immunostaining was performed on human renal tissue according to the procedure manual as a control to ensure that it provides similar results to those obtained with the automated procedure.
For double-staining experiments in human tissue, the first labeling was manually performed and revealed by an alkaline phosphatase-conjugated streptavidin complex (1:250; DAKO) followed by BCIP/NBT supplemented with 1 mmol/l of levamisole (to quench endogenous alkaline phosphatase activity). To block antibody cross-reactivity and retrieve epitopes, slides were microwaved twice for 5 min at 360 W in citrate buffer, pH 6.0. Sections were then processed on the Ventana automated instrument for the second immunostaining that was revealed by a horseradish peroxidase reaction. Controls included primary antibody switching between manual and automated procedure, omission of the second antibodies, replacement of the primary antibody by irrelevant monoclonal antibody of the same isotype, or preimmune rabbit serum.
Protein Extraction, Immunoprecipitation, Gel Electrophoresis, and Western Blot
Tissues were homogenized in extraction buffer [PBS, pH 7.4, 0.1% (v/v), Triton X-100, 0.05% (w/v) SDS] in the presence of protease inhibitor cocktail (Protease Arrest; Calbiochem, San Diego, CA), followed by low-speed centrifugation at 1000 × g to remove nuclei and cell debris. The supernatant was then centrifuged at 14,000 × g for 1 hr and used for further analysis. Immunoprecipitation used the IgM MAb anti-HNK-1 linked to Dynabeads M-450 rat anti-mouse IgM (Dynal; Oslo, Norway) or IMMUNOcatcher kit (CytoSignal; Irvine, CA) according to manufacturer's instructions. In some experiments, the extracted lysate or the immunoprecipitated proteins were deglycosylated with chondroitinase ABC, recombinant, heparitinase I, Flavobacterium heparinum (Calbiochem), keratanase (Seikagaku; Falmouth, MA), or neuraminidase (Sigma-Aldrich; Lyon, France) using buffers and conditions as recommended by the manufacturer.
Protein extracts were diluted in SDS sample buffer, heated for 15 min at 75C, and separated in 4% SDS-polyacrylamide gels according to the method of Laemmli (1970). For Western blotting, proteins were transferred from gels to polyvinylidene difluoride membranes (Bio-Rad Laboratories; Hercules, CA). Membranes were blocked with 10% non-fat dry milk in PBS-0.1% Tween and 0.1 mM levamisole for 45 min at 37C and then probed with MAb anti-HNK-1 (1:1000 ascite fluid diluted) or MAb272 anti-L1CAM (1.0 μg/ml) followed by alkaline phosphatase-conjugated secondary antibody (Promega; Heildelberg, Germany) diluted 1:5,000. Primary antibodies were omitted, or isotype-matched murine IgM or IgG was used as the primary antibody for control blots. The immunoreaction was visualized by enzymatic reaction with BCIP/NBT complex.
Results
HNK-1 Expression in Developing Human Kidney
Immunohistochemistry experiments performed at different embryonic ages (8, 10, 17, 18, 25, and 27 weeks of gestation) showed that HNK-1 epitope was present both in developing nephron and ureteral bud derivatives but with a very restrictive pattern. In ureteral bud derivatives, HNK-1 was transiently expressed at the tip where the elongating collecting duct splits in two parts in touch with the metanephric blastema (Figure 1A). It was also present in the limited area where the collecting duct connects with the developing nephron (Figure 1D). Of interest, we showed previously that the L1CAM cell adhesion molecule was specifically expressed in ureteral bud derivatives, including the tip and the connecting segment (Debiec et al. 1998; Allory et al. 2005). However, unlike L1CAM, HNK-1 was absent from more mature segments of collecting ducts as well as from mature connecting segments (Figure 1 and Figure 2).
Although HNK-1 was not initially observed in the metanephric blastema, it was detected throughout the successive stages of nephrogenesis, from the early primary vesicles to the late maturing nephron stage (Figures 1A-1D). A weak signal was observed within a subset of epithelial cells in the renal vesicles (Figures 1A and 1B) and comma-shaped bodies (Figure 1C). In the early S-shaped body, HNK-1 was detected in the tubular progenitors of the nephron (Figure 1D) that correspond to the future proximal tubule, loop of Henle, and distal tubule that was identified in homologous sections by E-cadherin immunolabeling (Figures 1E and 1F). The HNK-1 epitope appeared to be absent from future podocytes and Bowman's capsule cells (Figure 1D). As nephrogenesis proceeded, HNK-1 expression was downregulated in the proximal and distal tubules. To illustrate the localization of HNK-1 expression in relation to tubule development, a lower magnification image showing both the nephrogenic zone and the deeper cortex containing more mature nephrons is presented in Figure 2A. In maturing glomerulus, a strong expression of HNK-1 was observed in the neck of differentiating glomerulus (Figure 2B) and in numerous cells in the initial segment of proximal tubule (Figure 2C). At late development stage, HNK-1 was still observed in rare cells at the glomerulus neck (Figure 2D). In mature glomeruli and proximal tubules HNK-1 was absent, except in the cytoplasm of occasional isolated cells of proximal tubules (Figure 2E). Of note, scattered cells positive for cytokeratin 7 and described as plica cells have been previously reported in proximal tubules (Moll et al. 1991). By double immunostaining, these plica cells were distinct from the rare HNK-1 positive cells (Figure 2E). After the S stage, HNK-1 was not detected in distal tubules (Figure 2D), except in some elongating loops of Henle in the medulla (Figure 2F). In adult kidney, HNK-1 expression was maintained in, and virtually restricted to, these particular medullary segments of Henle's loops. HNK-1 epitope was not expressed in renal urothelium.
HNK-1 Expression in Adult Human, Rabbit, Rat, and Mouse Kidney
We further characterized the segments of loop of Henle that were HNK-1 positive in adult kidney. First, this expression was restricted to the inner medulla (Figure 3A). Whereas the collecting ducts, which expressed L1CAM, were entirely HNK-1 negative, a fraction of thin tubules in inner medulla (30-50%) showed a strong HNK-1 expression. This expression was cyto plasmic, sometimes with a more intense staining at the apical side. Some thin tubules were completely negative, and others displayed both positive and negative cells or only positive cells (Figures 3B and 3C). Double-staining experiments with HNK-1 and CD34 used as a vascular marker confirmed that HNK-1-positive tubule sections were distinct from vasa recta capillaries (Figure 3D). These HNK-1 tubules were also stained with the cytokeratin 7 antibody, providing further evidence for their epithelial nature (not shown). Tamm-Horsfall protein, which is specific for the thick ascending part of the loop of Henle, and HNK-1 immunostainings were mutually exclusive (Figure 3E). As we observed both HNK-1 positive and negative HNK-1 thin tubules, we postulated that HNK-1 might be confined either to descending or ascending part of the thin loop of Henle. We used aquaporin 1 and UTA2, which are specific for the descending part. We showed that most of the HNK-1-positive thin tubules were not stained by these antibodies (Figure 3F). These observations suggested that HNK-1 was specifically expressed in ascending part of the thin loop of Henle. We also observed some thin tubules with HNK-1-positive cells admixed with distinct aquaporin 1-positive cells. This result is consistent with the fact that descending and ascending cell types are admixed together at the tip of the loop of Henle.
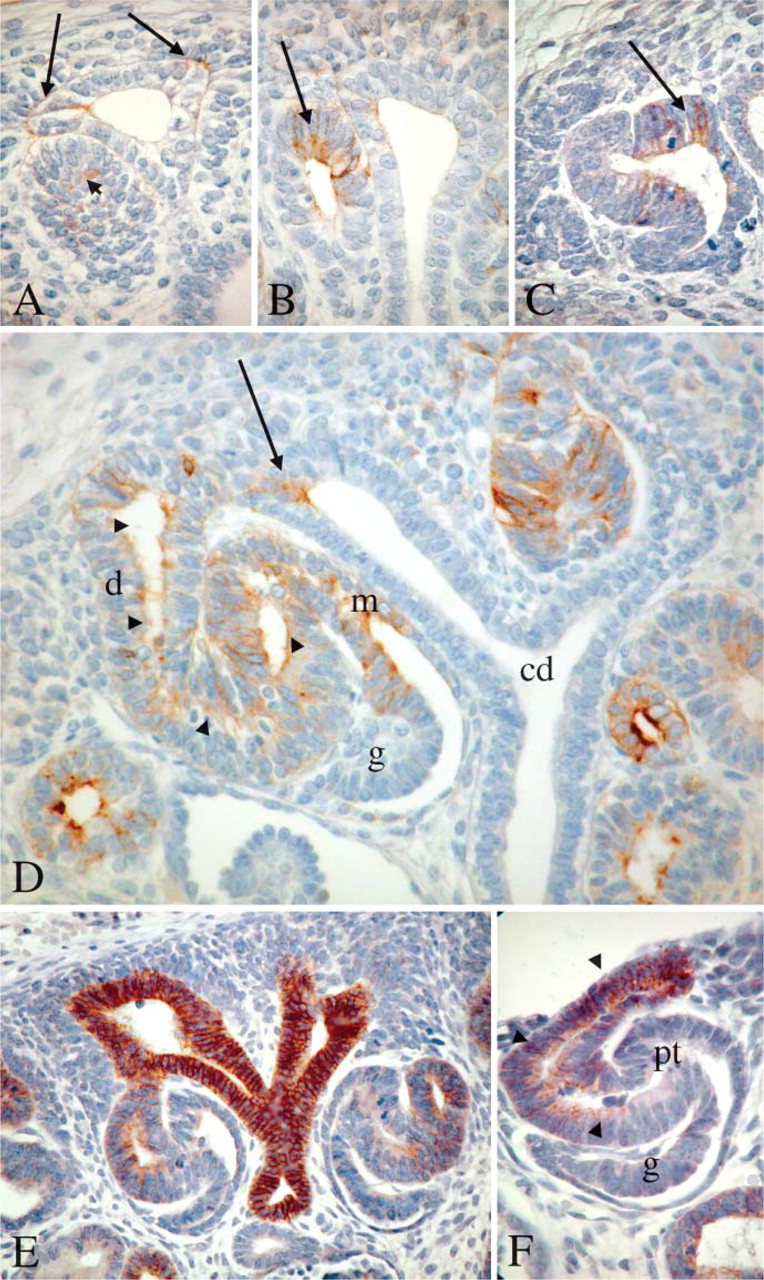
HNK-1 expression in early developmental stages of human kidney. (
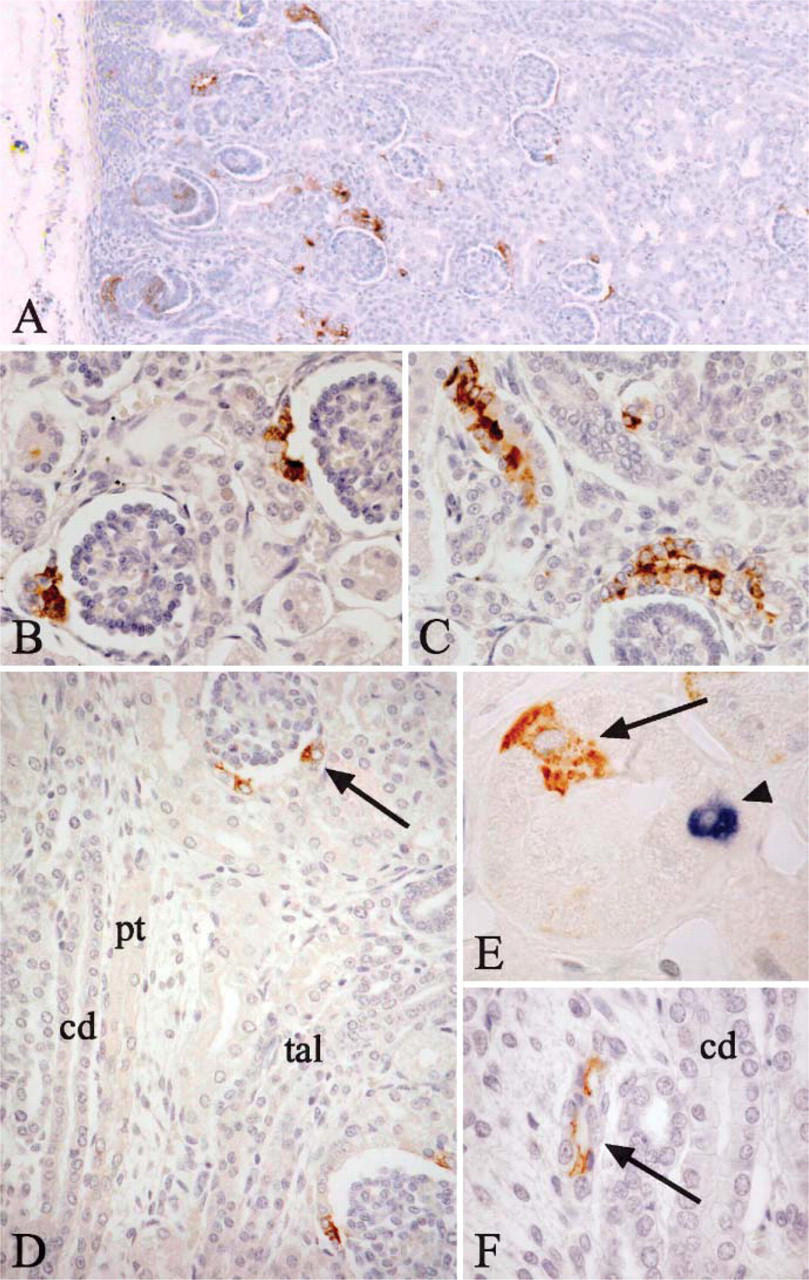
HNK-1 expression in late developmental stages of human kidney. (
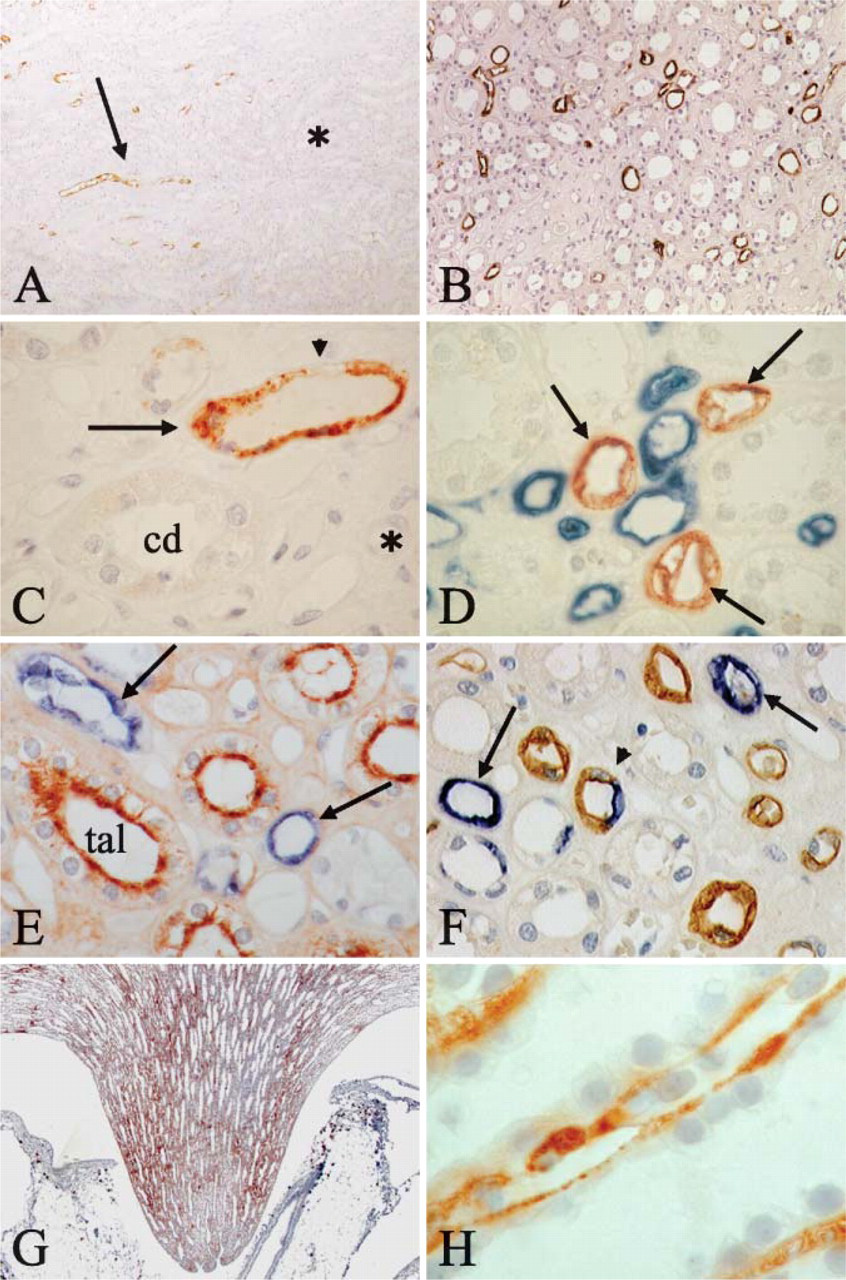
HNK-1 expression in human and rat adult kidney. (
HNK-1 immunohistochemical analysis was also performed in rabbit, mouse, and rat kidney. In rat, we observed a strong labeling of thin tubules in the medulla as described in human kidney (Figures 3G and 3H). The same restricted pattern was observed during development in collecting duct tips and nascent proximal tubules (not shown). In rabbit medulla, HNK-1 was also detected as in human kidney in thin tubules, but the expression seems to be confined only to a small fraction of these tubules. In mouse, a weak staining by HNK-1 was only present in exceptional tubules (not shown).
Biochemical Analysis of the HNK-1-bearing Proteins
In the nervous system, the HNK-1 epitope can be detected on different proteins, including adhesion molecules like L1CAM or NCAM, and proteoglycans. Because of the specific but restricted pattern of expression during renal organogenesis, it was not possible to perform analysis to identify the molecules bearing HNK-1 at the various ontogenic stages. Thus, we focused on the adult stage and prepared medulla protein extracts from adult human and rat kidney. The experiments in rat kidney showed a broad smear of high molecular weight, suggesting a high level of glycosylation. Because this pattern was reminiscent of the one previously described for HNK-1-associated proteoglycans in the central nervous system (Krueger et al. 1992), we tested the hypothesis that HNK-1 was associated with proteoglycans in the loop of Henle. We submitted the HNK-1 immunoprecipitate to keratanase, chondroitinase, and heparitinase treatment. After combined chondroitinase and heparitinase treatment, three distinct protein bands appeared at 250 kDa and above, confirming that the renal HNK-1 was mainly carried by a heparan- and chondro-proteoglycan. A further treatment with neuraminidase, after chrondroitinase and heparitinase, enhanced the protein bands, suggesting that polysialic acids were also present on this HNK-1-positive protein. Keratanase treatment showed no effect (not shown). In human medulla protein extracts, a faint high molecular weight smear was present before enzymatic treatment, whereas one band above 250 kDa could be detected after mixed chondroitinase and heparitinase treatment, suggesting that also in the human adult kidney HNK-1 is associated with a proteoglycan (Figure 4).
HNK-1 Expression in Human Acute Tubular Necrosis and Renal Tumors
Because of the relatively diffuse expression of HNK-1 in developing proximal tubule that becomes restricted to a few cells in adult proximal tubule, we investigated whether HNK-1 could be re-expressed during acute tubular necrosis. HNK-1 expression was induced in proximal tubular cells in 8/10 patients with acute tubular necrosis after renal transplantation. HNK-1 was observed in numerous (three cases) or only a few tubules (five cases), but in each positive tubule most of the cells expressed HNK-1. The staining predominated at the apical membrane and was often cytoplasmic (Figure 5A).
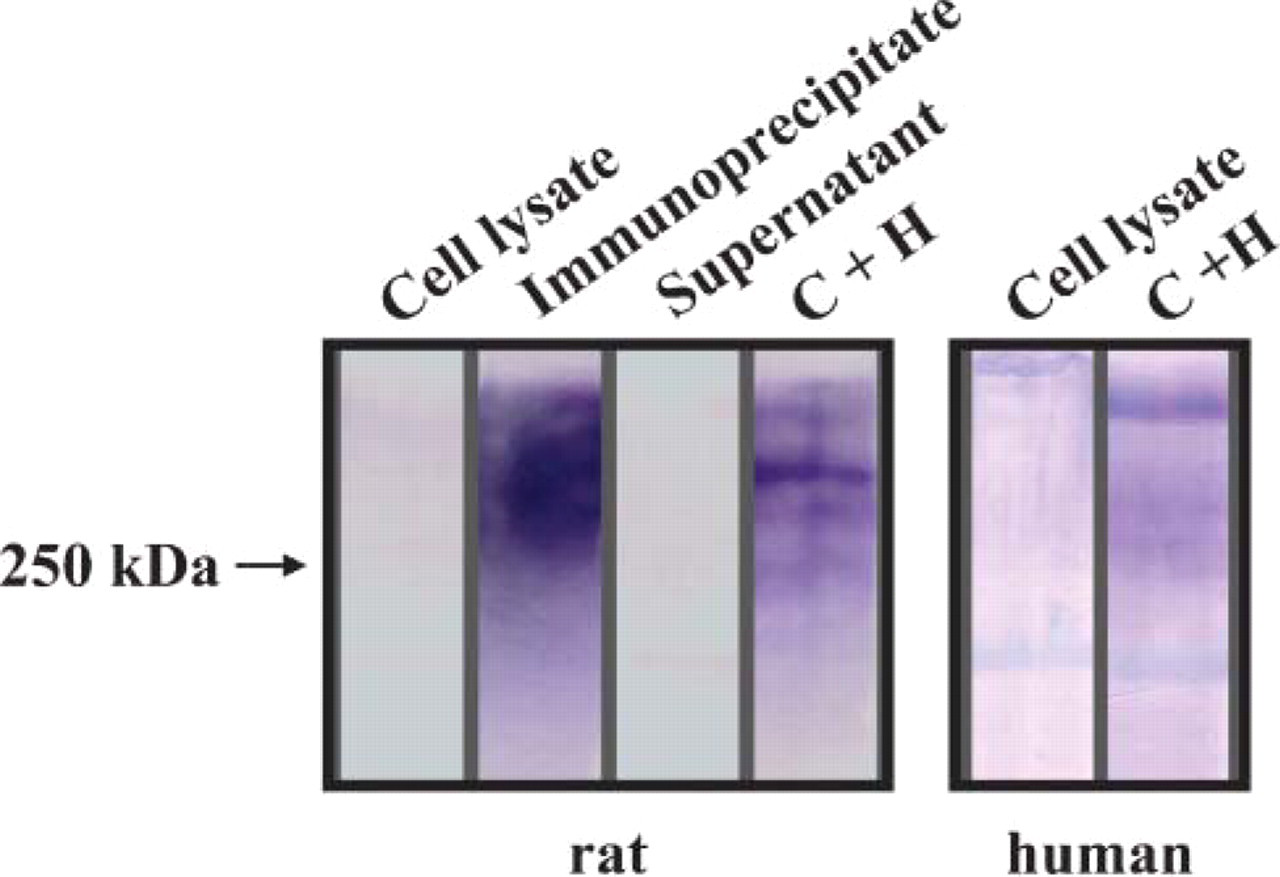
Protein extracts from rat and human medulla immuno-precipitated with HNK-1 antibody and analyzed by Western blot using HNK-1 antibody. Note the strong and broad protein smear above 250 kDa in rat HNK-1 immunoprecipitates, which partially resolved in distinct bands after combined treatment with chondroitinase and heparitinase (Lanes C + H). There was only faint staining in native lysate before immunoprecipitation and no specific staining in supernatant. The faintly stained protein smear in human renal cell lysate resolved in more distinct bands after combined heparitinase and chondroitinase treatment.
In nephroblastoma, a focal, heterogeneous HNK-1 expression was observed in the epithelial component in 6/9 cases (Figures 5B and 5D). In adult renal epithelial tumors, HNK-1 was strongly expressed in 21/78 papillary carcinomas (Figures 5C and 5D). It was absent from oncocytoma and occasionally detected in clear cell (1/71), chromophobe cell (2/48), and low-grade tubular mucinous (2/11) variants (Figure 5D). In renal papillary carcinomas, HNK-1 expression was observed more frequently in type 1 (13/33) than in type 2 (8/45) (p=0.04, Fisher exact test). Among these 21 HNK-1-positive tumors, 19 also expressed cytokeratin 7. Expression of HNK-1 was not correlated with nuclear grade, pTNM stage, or MUC-1 and Ki67 immunolabelings.
Discussion
Most studies regarding the expression and function of the HNK-1 epitope were performed in the nervous system (Schachner et al. 1995). Here we report the first description of sulfated HNK-1 carbohydrate epitope in the developing and mature kidney. The HNK-1 expression pattern differed in the adult from that observed during development, suggesting a developmental regulation of synthesis and a role in the fine-tuning of function of carbohydrate-bearing molecules.
HNK-1 epitope exhibits a dynamic, segmental expression pattern during nephron development. One of the major features of nephrogenesis is the coordinate development of the ureteric bud and the mesenchymally derived epithelium (Vainio and Muller 1997). These interactions culminate when ureteric bud-derived collecting duct connects with mesenchymal-derived epithelium to form the continuous tubule of nascent nephron. Our results show that HNK-1 accumulates at the sites of fusion and therefore could mediate the epithelial reorganization that occurs when the renal vesicles fuse to the ureteric bud. The HNK-1 carbohydrate added transiently to the numerous cell recognition molecules expressed at this place might synchronize signaling mechanisms that regulate the fusion event. However, the exact mechanism of fusion remains to be established.
Progression from pretubular aggregates to comma-and S-shaped bodies requires the conversion and patterning of mesenchymal cells into highly organized and polarized epithelia (Horster et al. 1999). The epithelium of the nephron is composed of a variety of specific cell types. Determination of cell identity is specified early in development, but molecular mechanisms involved in this process are unknown (Al-Awqati and Oliver 2002). Appearance of the HNK-1 epitope expression in mesenchymal derivatives is correlated with formation of the polarized epithelium. Its selective expression only within a small population of cells in renal vesicles and comma-shaped bodies provides evidence for discrete stages in cellular differentiation and/or for a subset of cells with unique functional properties for which HNK-1 epitope could be a marker. Later in development, the HNK-1 epitope was expressed in the region of the S-shaped body destined to become the distal tubule, the loop of Henle, and the proximal tubule, respectively. However, its expression in the distal and proximal tubules was transient, disappearing in more mature stages. HNK-1 signaling pathways control cell identity in a variety of systems. A wealth of studies have suggested that HNK-1 is involved in organization of early neuronal setting and axonal growth patterns (Cooke et al. 1994; Sela-Donenfeld and Kalcheim 1999; Jeffery et al. 2004). Once neurons reach their destination, HNK-1 is downregulated and virtually disappears. Because a parallel evolution is observed in the kidney proximal and distal tubules, we suggest that HNK-1 epitope might be implicated in cell organization and growth in the renal tubule.
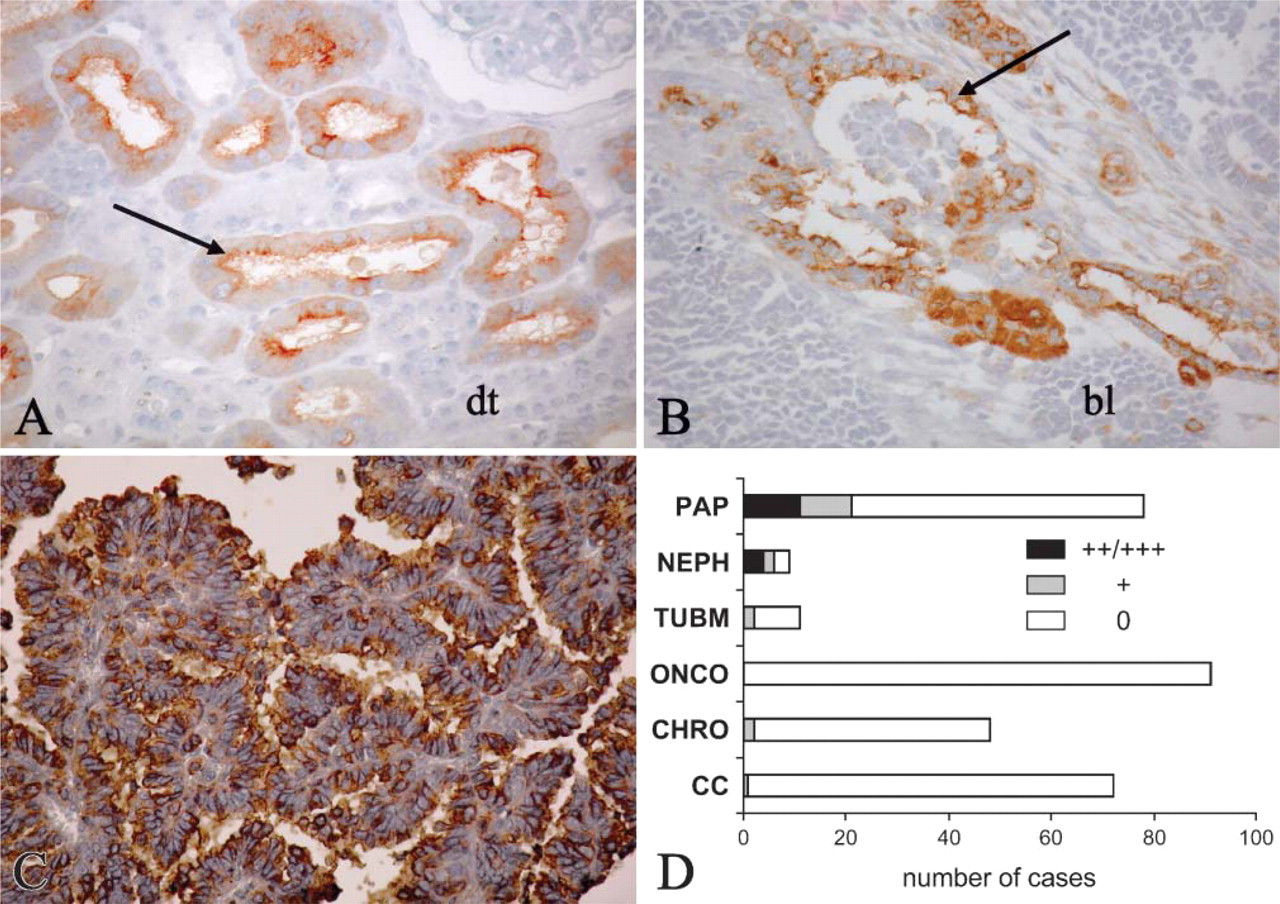
HNK-1 expression in pathology. (
In the adult kidney, the HNK-1 epitope was expressed predominantly on the apical membranes of the thin ascending loop of Henle in the inner medulla. This carbohydrate structure was carried by high molecular weight heparan- and chondro-proteoglycans as shown by Western blot analysis. Although the role of the continuous expression of the HNK-1 epitope in the mature thin ascending limb is not understood, its highly restricted expression pattern suggests that this epitope might be involved in a critical function of this nephron segment. In contrast, unlike the nervous system, the adult kidney does not feature the HNK-1 epitope on the cell adhesion molecule L1, which has a different topography of expression (Allory et al. 2005).
Despite similar expression of sulfated HNK-1 epitope in human, rabbit, and rat kidney, we failed to identify that epitope in the mouse kidney. On the other hand, in the adult mouse kidney the non-sulfated form of HNK-1 carbohydrate has been recently shown to be expressed on the brush border of kidney proximal tubules and in thin ascending limb of Henle's loop in inner medulla (Tagawa et al. 2005). In the brush border of proximal tubule cells, the non-sulfated HNK-1 epitope was expressed on two major metalloproteases, aminopeptidase N (CD13) and meprin α, suggesting that this carbohydrate may be involved in the regulation of their activity (Tagawa et al. 2005). However, the protein-bearing non-sulfated HNK-1 in the mouse inner medulla was not identified. In our experiments the sulfated HNK-1 carbohydrate was carried by a heparan- and chondro-proteoglycan of high molecular weight, whereas in the mouse inner medulla the non-sulfated HNK-1 seemed to be associated with lower molecular weight glycoproteins (Tagawa et al. 2005).
Analysis of changes in the HNK-1 epitope expression under pathological conditions may be helpful to find out both its physiological role and its significance in renal disease. During recovery from injury, surviving epithelial cells dedifferentiate and proliferate, eventually replacing the irreversibly injured tubular epithelial cells. The dedifferentiation of renal tubular cells that recapitulate gene expression patterns typical of the developing nephron has implications for the regulation of renal repair. Because the HNK-1 epitope is markedly upregulated on the injured proximal tubules in acute tubular necrosis, we suggest that this epitope might be involved in proximal epithelial cell repair.
Nephroblastoma is a malignant renal tumor in childhood derived from the remnant of immature renal tissue. We and other investigators (Satoh et al. 2000) detected the HNK-1 epitope only on the epithelial component, whereas the blastema cells and the fibrous stroma were negative. Metanephric adenoma, another renal tumor that usually behaves in a benign fashion, also expresses the HNK-1 epitope (Muir et al. 2001; Skinnider et al. 2005). Expression of the HNK-1 epitope in both tumors is reminiscent of the antigen expression pattern seen in the developing nephron. However, the HNK-1 epitope was not expressed in highly aggressive clear cell RCCs, which are known to originate from the proximal tubule (Motzer et al. 1996).
Metanephric adenoma has morphologic similarities to papillary carcinomas, but the immunohistochemical pattern of these two tumors is distinct (Muir et al. 2001; Skinnider et al. 2005). A combination of HNK-1 and CK7 was previously proposed as being helpful in distinguishing metanephric adenoma (typically HNK-1+ or CK7- or only focally+) and papillary carcinomas (typically HNK-1-, CK7+) (Muir et al. 2001). However, we found that 21 papillary carcinomas expressed HNK-1, among which 19 also expressed CK7. Our observation that HNK-1 is expressed in 21/78 papillary carcinomas from our series supports previous cytogenetic and clinical observations suggesting that metanephric adenoma and papillary carcinoma are closely related (Brown et al. 1997; Hes et al. 2003), as well as the existence of a continuum between the two types of tumors.
Although the HNK-1 epitope is present in the prospective loop of Henle in the S-shaped body and throughout the normal adult thin ascending limb of Henle, it is absent from tumor cells in mucinous tubular carcinoma. Mucinous tubular and spindle cell carcinomas are recently described tumors that are thought to originate from the loop of Henle (Parwani et al. 2001; Ferlicot et al. 2005). The absence of the HNK-1 epitope from those carcinomas suggests that either these tumors originate from cells of the thick loop of Henle that do not express the epitope, or that HNK-1 is lost during malignant transformation.
In summary, our work provides a comprehensive expression map of sulfated HNK-1 epitope during early nephron development in adult and in pathological conditions. Our results suggest that molecules carrying the HNK-1 epitope may play an important role in critical stages of renal development and in the functioning of thin ascending loop of Henle. Further identification of those molecules that bear the HNK-1 epitope may shed light on important aspects of kidney physiology and pathobiology.
Footnotes
Acknowledgments
This work was supported by grants from Institut National de la Santé et de la Recherche Médicale (INSERM), Paris 6 University, and Association pour l'Utilisation du Rein Artificiel. Y.A. was a recepient of a Poste d'Accueil INSERM.