
Abstract
Glomerular endothelial and mesangial cells may originate from the metanephric mesenchyme. We used the MAb Thy1.1, a mesangial cell marker in the adult rat kidney, and rat endothelial cell markers MAb RECA-1, MAb PECAM-1 (CD31), and MAb Flk-1 as potential markers to characterize the spatial and temporal distribution of mesangial and endothelial cell precursors during nephrogenesis in the rat. At early stages of glomerulogenesis, RECA-1- and Thy1.1-positive cells were detected in the metanephric blastema at 14 days post conception (dpc) embryos and 15 dpc, respectively, with Thy1.1 expression in cells surrounding the ureteric bud. At 17 and 18 dpc, both RECA-1- and Thy1.1-positive cells were found in the cleft of the S-shaped bodies and in the capillary loops of maturing glomeruli. Double staining for BrdU, a marker of proliferation, and for RECA-1 or BrdU and Thy1.1 also localize in the cleft of S-shaped bodies and in glomerular capillary loops at later stages of development. PDGFRβ co-localizes in cells expressing endothelial or mesangial markers. The data suggest that endothelial and mesangial cell precursors share common markers during the course of glomerulogenesis and that full differentiation of these cells occurs at late stages of glomerular maturation. Thy1.1- and RECA-1-positive cells may be derived from the metanephric blastemal cells at early stages of kidney development. A sub-population of these Thy1.1- or RECA-1-positive cells may be precursors that can migrate into the cleft of comma and S-shaped bodies and proliferate in situ to form glomerular capillary tufts.
M
During metanephric development, several growth factors, such as vascular endothelial growth factor (VEGF) (Breier et al. 1992; Kitamoto et al. 1997), fibroblast growth factor (FGF) (Fu et al. 1991), hepatocyte growth factor/scatter factor (HGF) (Woolf et al. 1995), and platelet-derived growth factor (PDGF) (Alpers et al. 1992; Betsholtz and Raines 1997) are known to be involved in glomerular capillary tuft formation. Among these factors, the role of PDGF is of particular interest because of the absence of glomerular mesangial cells in the PDGFB-chain and in PDG-FRβ-deficient mice (Leveen et al. 1994; Soriano 1994).
The general morphological features of renal development have been described in great detail. However, markers tracking progressive development of glomerular microvasculature are not optimal. To identify potential markers of precursors of the developing microvasculature, monoclonal antibodies (MAbs) were used to determine the spatial and temporal distribution of glomerular mesangial and endothelial cell precursors during kidney development. In addition, we mapped the expression of PDGFRβ in the developing kidney in an attempt to correlate its expression with the expression of endothelial or mesangial cell markers. We also used BrdU to identify the proliferative stages of cells during development of the glomerular capillary tuft.
Materials and Methods
Animals
Pregnant Harlan Sprague-Dawley rats were purchased at 10–11 days of gestation. At gestational day 13–19, mothers were anesthetized by IM injection of rat mixture (60% ketamine, 40% lidocaine) and embryos were collected. The age of the embryos was counted from the day of the vaginal plug (day 0). Embryos were dissected in 1 X PBS under a zoom model SZH Olympus stereomicroscope and embryonic kidneys were embedded in optimal cutting temperature (OCT) compound and frozen in liquid nitrogen. Kidneys were stored at −70C until cryosectioning.
Immunohistochemistry and Immunoflourescence
Frozen tissue was cut and prepared for indirect immunoperoxidase amplified by the avidin-biotin–peroxidase complex (ABC) method or alkaline phosphatase-anti-alkaline phosphatase technique (APAAP) as previously described (Guesdon et al. 1979; Cordell et al. 1984). Frozen sections (4 μm) were fixed in acetone and incubated with 0.6% H2O2 in methanol to block nonspecific peroxidase activity, with 0.01% avidin, 0.001% biotin to block localization of endogenous biotin activity, and with normal serum of the same species as the second antibody to block nonspecific antibody staining. The primary antibodies used include MAb RECA-1 (Serotec; Oxford, UK), PECAM-1 (CD31; Chemicon International, Temecula, CA), Thy1.1 [MAbs OX7 (Serolab; Crawley Down, UK), His50 (Pharmingen; San Diego, CA), and PDGFRβ (958) (Santa Cruz Technology, Santa Cruz, CA)]. Concentrations of primary antibodies ranges between 2 and 10 μg/ml. For the ABC method, the sections were incubated with biotinylated antibody to mouse or rabbit IgG, and then incubated with avidin-biotin–peroxidase complex (Vector Laboratories; Burlingame, CA), which was visualized by 3-amino-9-ethylcarbazole (AEC) in the presence of H2O2. The primary antibodies were omitted or replaced by normal mouse or rabbit IgG (5 μg/ml) as controls.
Dual-label Immunohistochemistry for PDGFRβ and PECAM-1 or Thy1.1
Frozen sections (4μm) were fixed in acetone and rehydrated in PBS with 1% BSA. Tissue sections were first incubated with normal nonimmune donkey IgG directly before the incubation with anti-PDGFRβ antibody, followed by an affinity-purified Cy3-labeled donkey anti-rabbit IgG absorbed with IgG derived from multiple species for dual labeling (Chemicon). Tissue sections were then incubated with anti-PECAM-1 antibody, followed by affinity-purified FITC-labeled donkey anti-rabbit IgG absorbed with IgG derived from multiple species for dual labeling (Chemicon) or direct immunostaining of Thy1.1 utilizing an FITC-labeled mouse MAb (Cedarlane; Nornby, Ont, Canada).
Dual-label Immunohistochemistry for Thy1.1 and PECAM-1 or Thy1.1 and Flk-1
Frozen sections (4 μm) were fixed in acetone and rehydrated in PBS with 1% BSA. Tissue sections were first incubated with an avidin block, then a biotin block, before blocking with normal nonimmune donkey IgG directly before the incubation with anti-PECAM-1 or anti-Flk-1 (Santa Cruz) followed by an affinity-purified Cy3-labeled donkey anti-rabbit IgG absorbed with IgG derived from multiple species for dual labeling (Chemicon). Tissue sections were then incubated with a biotinylated anti-Thy1.1 antibody followed by affinity-purified FITC-streptavidin-labeled IgG absorbed with IgG derived from multiple species for dual labeling (Chemicon). Sections were analyzed using an Olympus FV-500 confocal microscope. The excitation wavelength for the FITC is 488 nm and for the excitation wavelength for Cy3 is 543 nm.
BrdU Labeling and Double Immunostaining of BrdU with RECA1 or Thy1.1
Pregnant rats at 18 days of gestation were injected IP with BrdU (Sigma; St Louis, MO) at a dose of 1 g/kg body weight. Ninety minutes after BrdU injection, the rats were sacrificed and fresh embryonic kidneys were embedded in OCT, frozen in liquid nitrogen, and kept at −70C until use. Double immunostaining for BrdU and RECA-1 or BrdU and Thy1.1 was performed on cut frozen sections that had been fixed in acetone. RECA-1 or Thy1.1 staining with the ABC technique was performed first, followed by BrdU staining using a BrdU detection kit according to the manufacturer's instructions (Boehinger; Mannheim, Germany). Because RECA-1, Thy1.1, and BrdU are all MAbs, to avoid crossreaction between the two systems of detection, we used microwave oven treatment. Tissue sections that have been stained for the first antigen were immersed in 0.01 M sodium citrate buffer, pH 6.0, and heated (700 W microwave power output) for 8 min. This treatment denatures the primary antibodies and inactivates the peroxidase used for detection of the first antigen (Lan et al. 1995). Sections were then treated with 0.1% Triton in 0.1% sodium citrate at room temperature (RT) for 15 min before BrdU detection to increase the permeability of the nuclear membrane. Sections were then incubated with anti-BrdU MAb (1:10 dilution) at RT for 2 hr, followed by alkaline phosphatase-labeled polyclonal antibody against mouse IgG. BrdU detection was visualized by alkaline phosphatase substrate and chromogen 5-bromo-4-chloro-3-indolyl phosphate (BCIP)/nitroblue tetrazolium salt (NBT). Sections were then counterstained with methyl green. All sections were washed with PBS-BSA between all incubations. Sections were viewed and photographed with an Olympus AX70 research microscope.
Results
Thy1.1 Expression
Only sparse immunostaining of Thy1.1-positive cells could be detected in the metanephric blastema before 15 dpc surrounding ureteral branches (Figure 1A). However, at 15 dpc, cells expressing abundant Thy1.1 could also be detected surrounding the ureteric bud (Figure 1B). During comma- or S-shaped body formation, Thy1.1-positive cells localized at the cleft of the S-shaped body (Figure 1C). In addition, Thy1.1-MAb stained the cells dispersed between the differentiated epithelial structures throughout the metanephros. During capillary tuft formation of the embryonic kidney (Figure 1D) and in the adult kidney, Thy1.1 MAb strongly stained glomerular cells, predominantly in the mesangial area. However, Thy1.1 expression was not detected in any epithelial cells at any stage of nephron development.
RECA-1 Expression
At the earliest stage of kidney development and specifically at 13 dpc, RECA-1 MAb stained few cells near the periphery of the metanephric blastema. Very few RECA-1-positive cells were present within the metanephric blastema itself (Figure 2A). RECA-1-positive cells appeared within the kidney rudiment, starting at 14 dpc, scattered throughout the metanephric blastema, with cells forming cordlike structures. Faint staining was seen in the ureteric bud (Figure 2B). In isolated metanephric blastema at 15 dpc, the number of RECA-1-positive cells increased (Figure 2C). In the comma- and S-shaped body at 17 and 18 dpc, RECA-1-positive cells localized within the cleft of comma- or S-shaped bodies but were also detected in some non-epithelial structures dispersed throughout the metanephric blastema (Figure 2D). In the more mature glomeruli, strong staining of RECA-1-positive cells was found in the glomerular tuft in endothelial locations, but not in the podocytes (Figure 2E). In addition, at later stages of development, 19 and 20 dpc, arterial endothelial cells and the capillary network in the metanephric blastema strongly stained with RECA-1 (not shown). In adult kidney, in which kidney structures can easily be distinguished, the RECA-1 MAb specifically recognized vascular endothelial cells, including endothelial cells of glomeruli, peritubular capillaries, and endothelia of arteries of different diameters (not shown), confirming the specificity of this antibody for identification of rat endothelial cells (Duijvestijn et al. 1992).
Distribution of Thy 1.1 immunostaining in the developing rat kidney. (
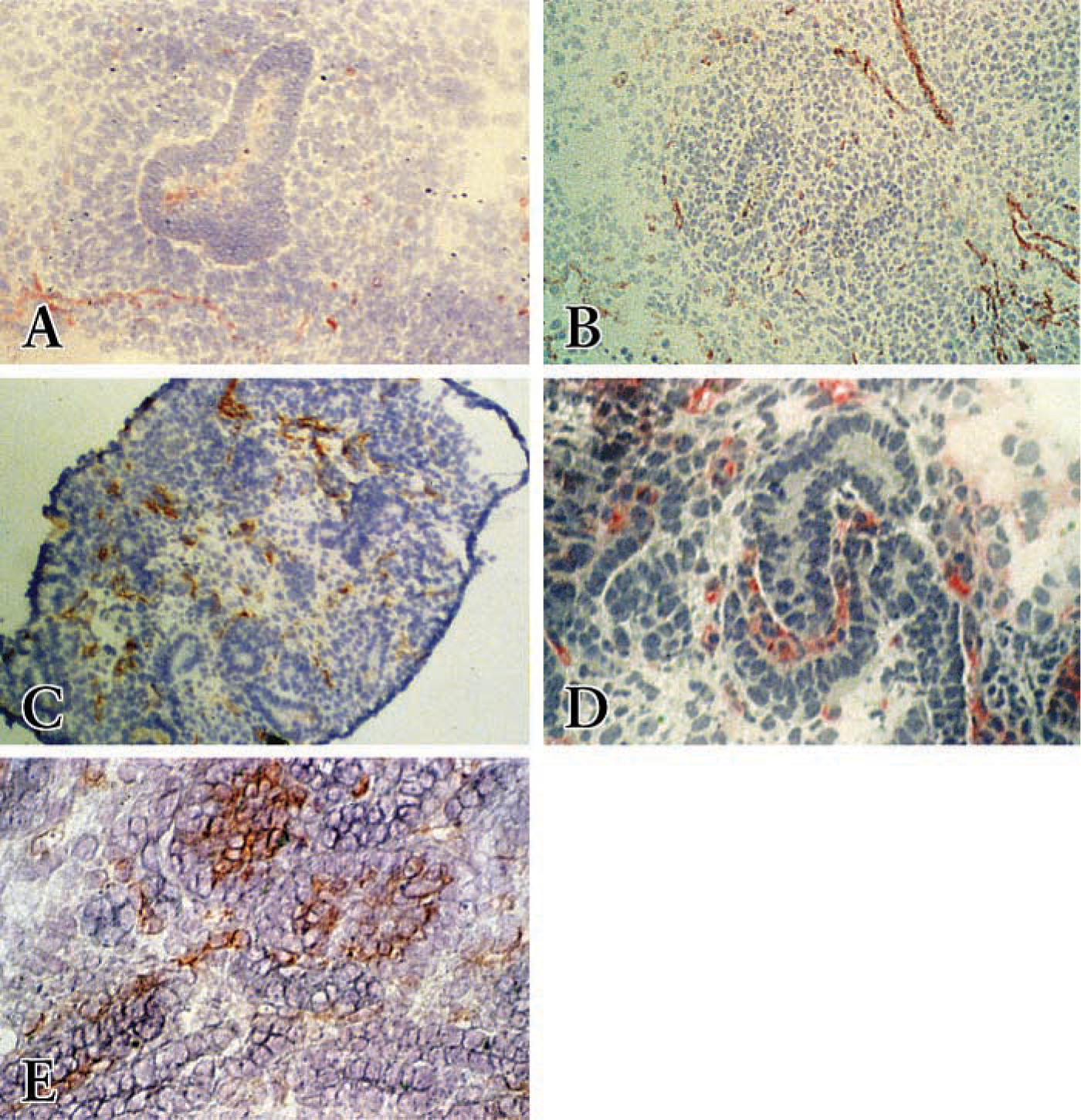
Distribution of RECA-1 immunostaining in the developing rat kidney. (
PDGFRβ Expression
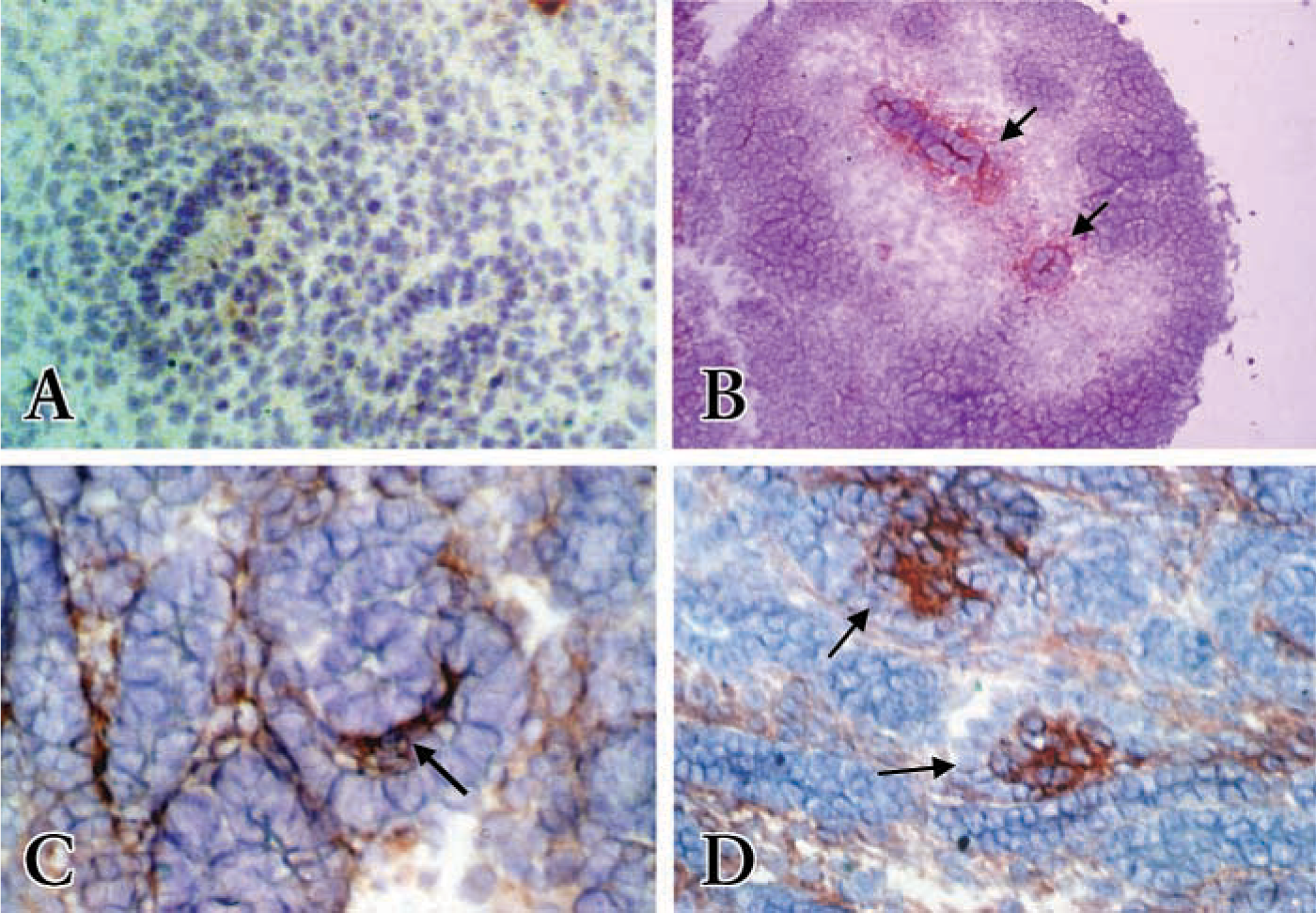
PDGFRβ is essential for mesangial cell development (Soriano 1994). In the early stages of nephron development, PDGFRβ is expressed throughout the undifferentiated metanephric blastemal cells and also faintly in the ureteric bud. As development continues, epithelial cell condensates form around the tips of the ureteric bud and eventually form comma- and S-shaped bodies. These epithelial cell condensates are PDGFRβ-positive (not shown) (Arar et al. 2000). As the nephron develops into comma- or S-shape bodies, PDGFRβ-positive cells decrease in the epithelial comma- or S-shape-bodies. However, PDGFRβ now localizes to cells within the cleft of comma- or S-shaped bodies (Figures 3B and 3H, red stain). In capillary loops of forming glomeruli, endothelial and mesangial cell precursors strongly stain for PDGFRβ, whereas glomerular epithelial cells are negative (Figures 3E and 3K, red stain). In addition, some interstitial cells dispersed between differentiated epithelial structures are positive but stain with less intensity. In adult kidneys, we detected only very faint PDGFRβ immunostaining in glomeruli within the mesangium (Figure 3N).
PECAM-1 Expression
Platelet endothelial cell adhesion molecule-1 (PECAM-1, CD31) is expressed on the surface of platelets, myeloid cells, and certain lymphocyte subsets (Watt et al. 1995) and is expressed over the entire plasma membrane of endothelial cells (Scholz and Schaper 1997). PECAM-1 has been used as a marker for developing endothelial cells during cardiovasculature development (Baldwin et al. 1994), lung development (Buck et al. 1996; Marszalek et al. 2000), and development of endothelial cells from ES cells (Redick and Bautch 1999). During nephron development, PECAM-1-positive cells show the same staining pattern that we earlier showed with RECA-1 (Figure 2). Staining with the PECAM-1 antibody provides strong and bright fluorescence compared with RECA-1 antibody staining. Therefore, PECAM-1 was used in dual immunofluoresence staining. In the comma- and S-shaped body, PECAM-1-positive cells localized within the cleft of comma- or S-shaped bodies (Figure 3A, green stain). In the more mature glomeruli, PECAM-1 positive cells are seen at the periphery of the glomerular tuft but not in the podocytes (Figure 3D, green stain). In the adult kidney, PECAM-1 antibody preferentially stains the endothelial cells in the glomerulus (Figure 3M).
Dual immunostaining of PDGFRβ with PECAM-1 (
Dual Immunostaining of PDGFRβ with PECAM-1 or Thy1.1
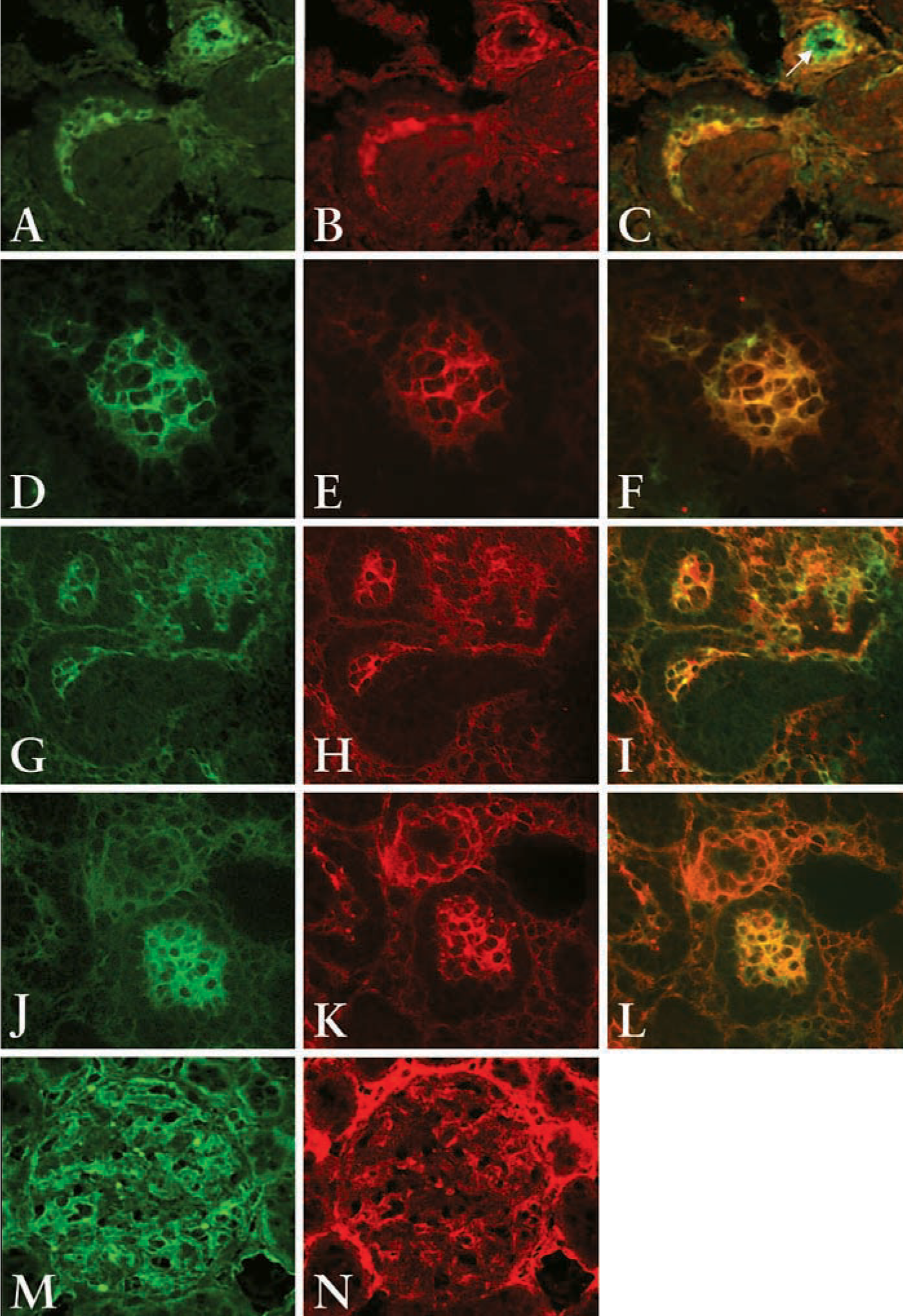
To verify if PDGFRβ co-localizes with PECAM-1 or Thy1.1, dual-label immunohistochemistry was employed using two separate fluorescence tags. At the comma- and S-shaped body stage, the cells in the cleft of S-shaped bodies double stained with PECAM-1 and PDGFRβ antibodies (Figure 3C, yellow-orange stain) or Thy1.1 and PDGFRβ antibodies (Figure 3I, yellow-orange stain) suggesting that at this stage of glomerulogenesis cells in the cleft of the S-shaped bodies express both PDGFRβ and endothelial or mesangial cell markers. Note the specificity of PECAM-1 (green stain only) to detect endothelial cells in the arteriole (arrow, Figure 3C). In glomerular capillary loops, the PDGFRβ antibody and PECAM-1 preferentially double stained the cells at the periphery of the glomerular loop but not the podocytes (Figure 3F, yellow-orange stain). PDGFRβ and Thy1.1 also double stain cells of the glomerular capillary loop except podocytes (Figure 3L, yellow-orange stain). However, there is some specific staining of Thy1.1 and PECAM-1 that does not co-localize with the PDGFRβ in the maturing glomerulus, suggesting that the cells in the maturing glomerulus are beginning to differentiate into their respective cell types.
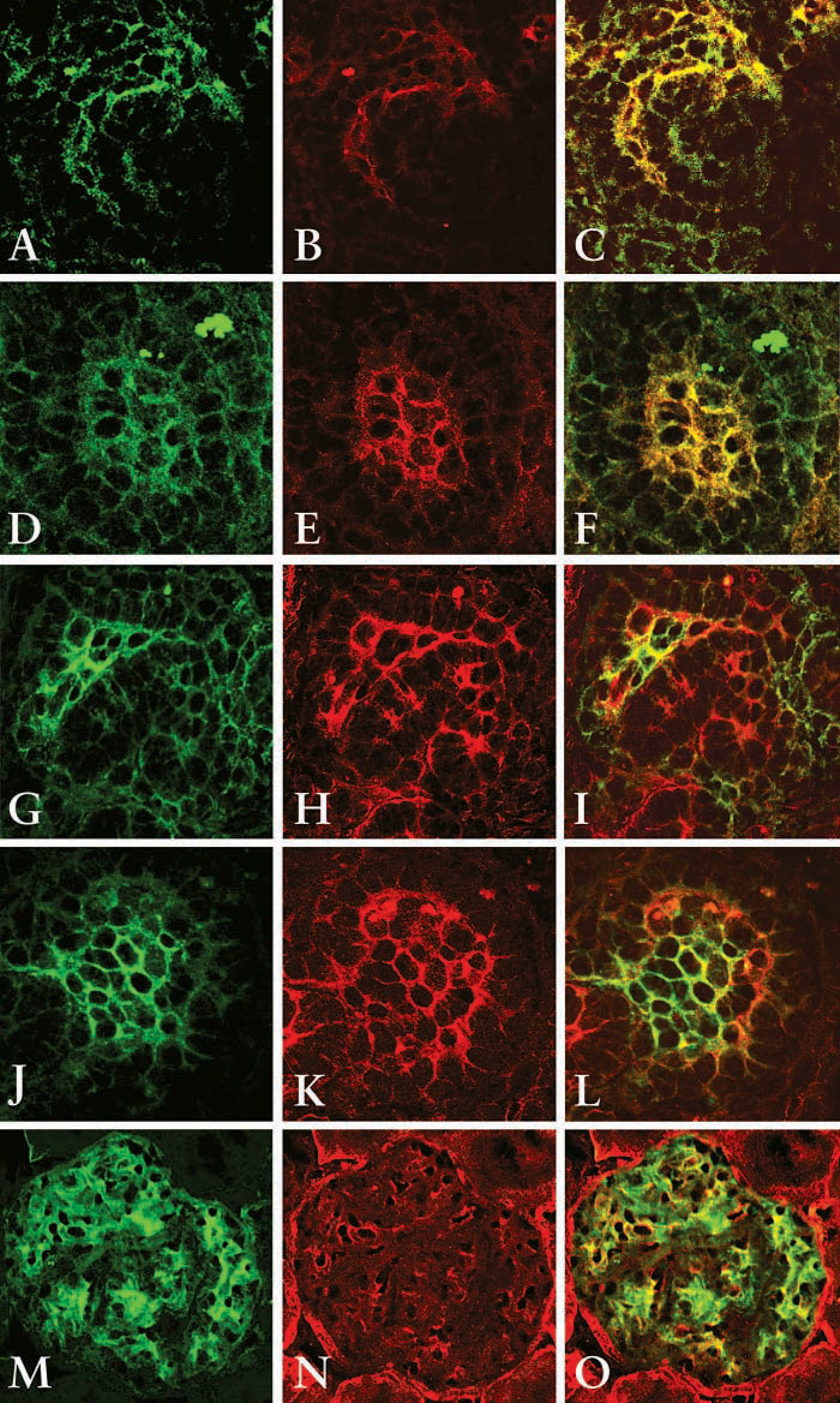
To further confirm that the cells in the S-shaped bodies express both mesangial and endothelial cells markers, dual staining of Thy1.1 and PECAM-1 or Flk-1 (a marker for mature and developing endothelial cells) was performed and analyzed using confocal microscopy. Both PECAM-1 and Flk-1 positive cells localize within the cleft of the S-shaped body (Figures 4B and 4H, respectively, red stain). Thy1.1 also localizes to the cleft of the S-shaped body (Figures 4A and 4G, green stain). Dual labeling shows that cells in the cleft of the S-shaped body co-express Thy1.1 and PECAM-1 (Figure 4C, yellow) or Flk-1 (Figure 4I, yellow), suggesting that cells at this stage of glomerulogenesis share a common phenotype with mesangial cells and endothelial cells. At the more mature glomerulus stage, Thy1.1 is expressed in the periphery of the capillary loop (Figures 4D and 4J). PECAM-1 and Flk-1 are also localized in the periphery of the capillary loop (Figures 4E and 4K, respectively). In the maturing glomerulus, cells in the capillary loop express both Thy1.1 and PECAM (Figure 4F, yellow stain) or Thy1.1 and Flk-1 (Figure 4L, yellow stain), suggesting that cells at this stage of glomerulogenesis also share a common phenotype with mesangial cells and endothelial cells. However, there appear to be cells expressing only one of the markers (red or green stain alone), indicating that cells may have begun to fully differentiate into either endothelial cells or mesangial cells. In the adult kidney, where the cells are fully differentiated, Thy1.1 stains cells predominately in the mesangial area (Figure 4M, green stain), whereas Flk-1 stains endothelial cells within the glomerulus as well as peritubular endothelial cells (Figure 4N, red stain). In the merged image, there is some co-expression of Thy1.1 and Flk-1 (Figure 4O, yellow stain). However, there is clear specificity of the antibodies (green or red stain alone).
Dual Immunostaining of BrdU with RECA-1 or Thy1.1
In the fetal kidney at 18 dpc, BrdU staining was localized to undifferentiated metanephric blastema cells, some epithelial cells of the comma- or S-shaped bodies, cells in S-shaped bodies, and some glomerular cells in the capillary loops of forming glomeruli (Figures 5A and 5B). However, the epithelial podocytes in the capillary loops of forming glomeruli were generally negative for BrdU immunostaining (Figure 5C). BrdU double staining with RECA-1 or Thy1.1 revealed that cells in the cleft of S-shaped bodies double stained with BrdU and RECA-1 (Figure 5A) or with BrdU and Thy1.1 (Figure 5B), indicating that the cells in the cleft of the maturing glomerulus as well as other cells dispersed between epithelial structures throughout the metanephros are proliferating during the course of development. However, there is more proliferation in the glomerular tuft during glomerular maturation (Figure 5C).
Discussion
In this study we used immunohistochemical techniques and markers of mature mesangial cells, Thy1.1, endothelial cell markers, RECA-1, PECAM-1, and Flk-1, and PDGFRβ to identify the spatial and temporal distribution of these cell types or a subset of their precursors in the developing microvasculature and during glomerulogenesis.
With anti-Thy1.1 MAbs, which detect normal glomerular mesangial cells in the adult rat kidney, we detected positive staining of cells in the metanephric blastema only at 15 dpc. During early stages of development, Thy1.1-positive cells were located at areas surrounding the ureteric bud, suggesting that these cells are induced to express Thy1.1 by ureteric bud epithelium or, alternatively, that these cells migrate along the ureteric bud. Transplantation experiments clearly indicate that mesangial cells develop from the grafts and are therefore derived from the metanephric mesenchyme and not from an extrarenal origin (Hyink et al. 1996). These observations suggest that Thy1.1-positive cells in our studies are induced by ureteric bud. It is of interest that this pattern is similar to that recently reported by Robert et al. using endothelial cell markers, including VEGF receptor (Flk1) (Robert et al. 1996,1998). At later stages of nephron development, Thy1.1-positive cells are present primarily in the cleft of the S-shaped body. In the capillary loops of forming glomeruli, Thy1.1-staining was mainly detected in the mesangial area, perivascular cells, and peritubular cells, as reported in the adult rat kidney. Thy1.1-positive cells may differentiate from metanephric blastemal cells during ureteric bud invasion and may then be distributed at the interstitial/peritubular areas where Thy1.1 positive cells remain in the interstium. Therefore, at some stages of development Thy1.1 staining may not be specific for mesangial cells because a relatively large number of interstitial/stromal cells stain positive. However, it is likely that a subpopulation of these Thy1.1-positive cells migrate into the comma- and S-shaped bodies to mature into mesangial cells in the glomerular capillary tuft.
Dual immunostaining of Thy1.1 and PECAM-1 (
In addition, using endothelial cell markers RECA-1, PECAM-1, and Flk-1, we found that these antibodies recognize only endothelial cells of vascular structures, including glomerular and peritubular capillaries in the adult kidney, confirming the specificity of these antibodies for vascular endothelial cells. We demonstrate that, at early stages of kidney organogenesis, cells scattered throughout the metanephric blastema stain faintly with RECA-1 and become more prominent as development progresses. The RECA-1-positive cells are seen lining up in a cord-like manner at 15 dpc, as described by Tufro et al. (1999) using Flk-1 as a marker for endothelial cells. RECA-1-, PECAM-1-, and Flk-1 positive cells are found at later stages in the cleft of the S-shaped body and are contiguous with other endothelial cells throughout the metanephros. These antibodies also recognized cells within the capillary loop of formed glomeruli at later stages of development, as well as endothelial cells of mature glomeruli and blood vessels of the adult kidney.
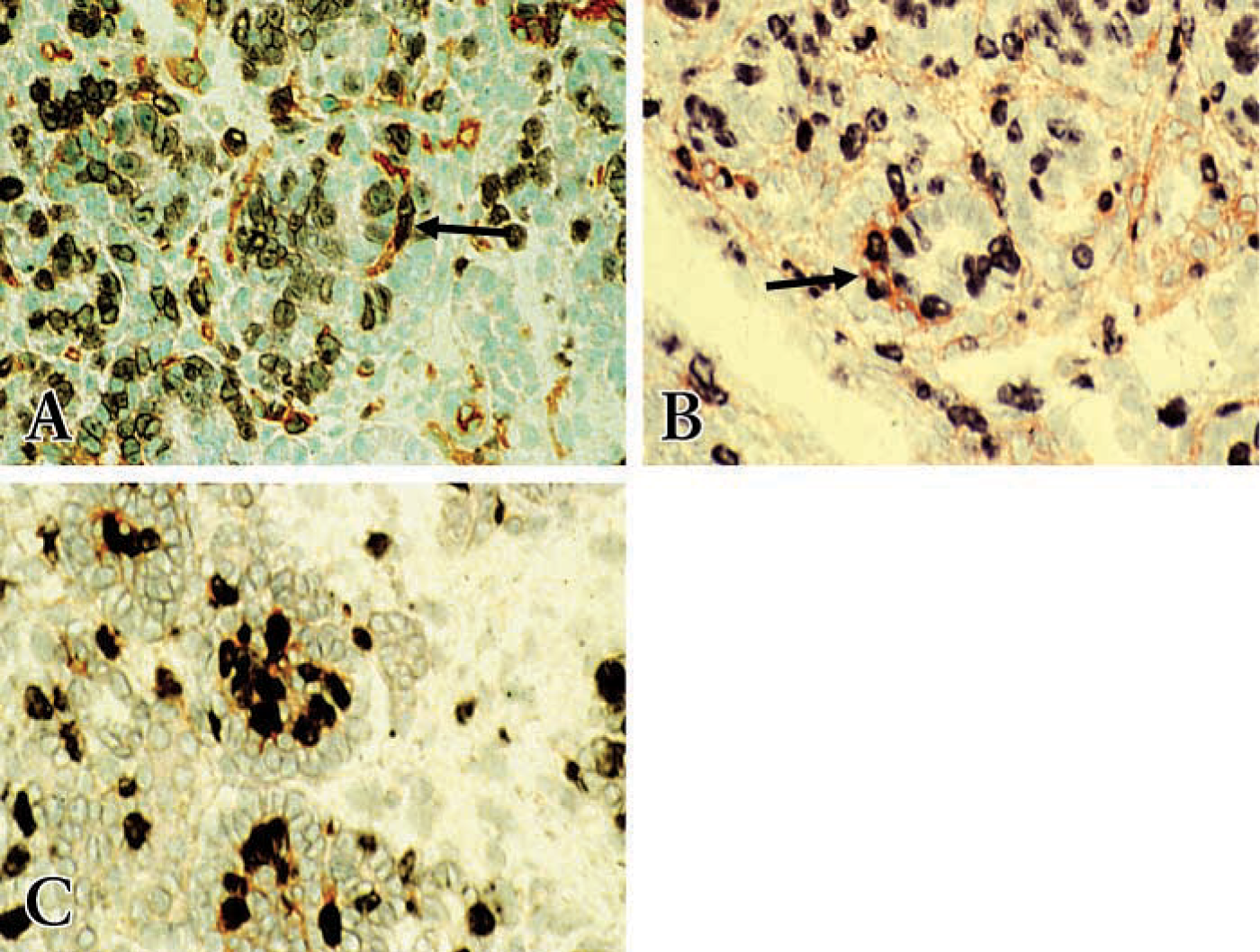
Dual immunostaining for BrdU and RECA-1 or BrdU and Thy 1.1. Proliferating cells stain dark purple by immunostaining with anti-BrdU with alkaline phosphatase reaction product BCIP/NBT. RECA and Thy1.1 stain reddish brown using immunoperoxidase with AEC substrate; see Materials and Methods for details. (
PDGF plays an important role in cell proliferation and migration during embryogenesis (Betsholtz and Raines 1997). We also mapped PDGFRβ expression at different stages of kidney development. Strong expression of PDGFRβ by the undifferentiated metanephric blastemal cells is similar to the observations of Alpers et al. (1992, 1993) and Arar et al. (2000). Shinbrot et al. (1994) showed PDGFRβ mRNA expression in mesenchymal components of many developing organs, with no expression in the epithelial components, suggesting a role for PDGFRβ in mesenchymal-epithelial interactions during organ development. Moreover, PDGFRβ mRNA was detected in mesenchyme surrounding the endothelium, which may implicate PDGFRβ in the growth and development of blood vessels (Shinbrot et al. 1994). Interestingly, we find co-localization of PDGFRβ not only with Thy1.1 but also with PECAM-1 in the cells in the cleft of comma-and S-shaped bodies, suggesting that the PDGFB-chain may play a role in cell migration and/or proliferation of precursors of both glomerular endothelial and mesangial cells. PDGFRβ also stained glomerular endothelial cells and mesangial cells in the capillary loops of forming glomeruli in the developing kidney, as demonstrated by dual immunostaining. In fact, PDGFB-chain expression by glomerular visceral epithelial cells at the comma- and S-shaped body stage, and by glomerular mesangial cells in capillary loops of formed glomeruli of the human embryonic kidney as shown by Alpers et al. (1992), indicates that the PDGFB-chain can have paracrine or autocrine effects to induce migration of glomerular endothelial and mesangial cell precursors into the cleft of comma- and S-shaped bodies as well as their proliferation in situ. BrdU staining of these cells in the cleft of S-shaped body and in capillary loops of formed glomeruli confirm the proliferative activity of these cells. However, PDGFRβ staining was dramatically decreased in glomeruli of adult kidneys, implying a role for the PDGFB-chain in glomerular endothelial and mesangial cell development. Whereas PDGFRβ plays an obligatory role in mesangial cell development and maturation, such a role has not been demonstrated for endothelial cell development. There are other reports of mesangial and glomerular endothelial cells gradually losing other fetal immunohistochemical phenotypes, such as α-smooth muscle actin, vimentin, and L-CD, in the human (Naruse et al. 2000), and neuropilin-1 in the mouse (Robert et al. 2000).
Double staining with BrdU and RECA-1 or BrdU and Thy1.1 of the cells in the cleft of the S-shaped bodies and in the capillary loops of forming glomeruli demonstrate that these cells are proliferating at later stages of kidney development. These data suggest that, at early stages of kidney development, the scattered RECA-1-positive cells throughout the metanephric blastema are most likely precursors of endothelial cells (angioblasts). They can be derived from the undifferentiated metanephric mesenchymal cells, which then migrate into the cleft of the S-shaped body and proliferate in situ to develop into glomerular capillary tuft cells. These data, however, do not exclude the possibility of an extrarenal origin for RECA-1-positive cells at early stages of kidney development. Angiogenic sprouts can migrate from the vascular network into glomerular and peritubular capillary tufts, as initially proposed using embryonic kidneys grafted into avian or quail chorioallantoic membrane (CAM) and into the anterior eye chambers of newborn rats (Ekblom et al. 1982; Sariola et al. 1983). However, grafts of embryonic kidneys into the anterior eye chambers of adult animals strongly indicate the predominantly endogenous origin of glomerular endothelial and mesangial cells (Kloth et al. 1994; Hyink et al. 1996).
In this morphological study using mesangial and endothelial cell markers, we detected Thy1.1- and RECA-1-positive cells that had distinct staining patterns at very early stages of kidney development. Cells positive for Thy1.1 or RECA-1 may be derived from metanephric blastemal cells. At this stage in development, it is likely that the cells that are positive for Thy1.1 represent not only mesangial cell precursors but also other interstitial/stromal cells. However, during later stages of glomerulogenesis these Thy1.1- and RECA-1/PECAM-1/Flk-1-positive cells may represent precursors of mesangial and endothelial cells that migrate into the cleft of comma- and S-shaped bodies and proliferate in situ to form glomerular capillary tufts. During nephrogenesis, the interaction of the PDGFB-chain and PDGFRβ may have important roles in microvascular cell proliferation and migration, especially during glomerular maturation.
In conclusion, at early stages of nephron development we find differential staining of Thy1.1 and endothelial cell markers. At the S-shaped stage, we find co-localization of mesangial and endothelial cell markers with PDGFRβ and with BrdU, suggesting that mesangial and endothelial cell precursors in the cleft of the maturing glomerulus are proliferating during the course of glomerular microvascular development. In maturing glomeruli, there is still much overlap in the expression of these markers, although specificity of staining is seen in more mature glomeruli, indicating full differentiation of mesangial and endothelial cells in the mature glomerulus. Collectively, our data suggest but do not conclusively demonstrate that endothelial and mesangial cell precursors share common markers during glomerulogenesis. The selective absence of mesangial cells in the PDGFRβ-deficient mouse indicates that this receptor and its ligand are essential for mesangial but not glomerular endothelial cell development. However, this conclusion awaits full characterization of endothelial cells in the PDGFRβ-deficient mouse.
Footnotes
Acknowledgements
Supported by National Institutes of Diabetes and Digestive and Kidney diseases Grant DK-33665 (to HEA), and in part by the Research Enhancement Award Program, Dept. of Veterans Affairs Medical Research Service (HEA), American Heart Association, Texas Affiliate Grants-in-Aid 97G-439, Clinical Scientist Award from NKF (to MA). Y-CX was supported in part by NKF Texas Affilate.