
Abstract
A deregulation of several MUC genes (MUC1, MUC2, MUC3, MUC5AC, and MUC6) was previously demonstrated in breast carcinomas. Considering that recently we found the “non-mammary” MUC5B mRNA in primary breast tumors (Berois et al. 2003), we undertook the present study to evaluate the expression profile of MUC5B protein product in breast tissues, using LUM5B-2 antisera raised against sequences within the non-glycosylated regions of this apomucin. Expression of MUC5B by breast cancer cells was confirmed by immunocytochemistry, in situ hybridization, and Western blot on MCF-7 cancer cells. Using an immunohistochemical procedure, MUC5B apomucin was detected in 34/42 (81%) primary breast tumors, in 13/14 (92.8%) samples of non-malignant breast diseases, in 8/19 (42.1%) samples of normal-appearing breast epithelia adjacent to cancer, and in 0/5 normal control breast samples. The staining pattern of MUC5B was very different when comparing breast cancer cells (cytoplasmic) and non-malignant breast cells (predominantly apical and in the secretory material). We analyzed MUC5B mRNA expression using RT-PCR in bone marrow aspirates from 22/42 patients with breast cancer to compare with MUC5B protein expression in the primary tumors. Good correlation was observed because the six MUC5B-positive bone marrow samples also displayed MUC5B expression in the tumor. Our results show, for the first time at the protein level, that MUC5B apomucin is upregulated in breast cancer. Its characterization could provide new insights about the glycobiology of breast cancer cells.
M
With regard to mucin gene expression in the breast, MUC1 expression was observed on the apical surface of normal epithelium (Gendler and Spicer 1995). In breast cancers, MUC1 is frequently overexpressed and underglycosylated (Baldus et al. 2004). This mucin can act as a promoter of in vivo mammary gland transformation (Schroeder et al. 2004) and is implicated in metastases (Ciborowski and Finn 2002). Aberrantly localized MUC1 in the tumor cell cytoplasms or non-apical membrane is associated with a poor prognosis in breast cancer patients (Rahn et al. 2001). MUC1 is being evaluated as a target for tumor immunotherapy, and there are serum tumor marker(s) assays based on MUC1 detection (Vlad et al. 2004). Apomucins normally expressed in other tissues, and usually undetectable in normal breast, have also been studied in breast cancer. Expression of MUC2 was observed in 19% (Walsh et al. 1993) and 25% (Diaz et al. 2001) of invasive ductal carcinomas and in 11% of in situ mammary carcinomas (Walsh et al. 1993). It was shown that mucinous carcinomas of the breast exhibit strong expression of MUC2 (O'Connell et al. 1998; Matsukita et al. 2003). Intestinal MUC3 apomucin was also demonstrated in breast cancer (Cao et al. 1997; Diaz et al. 2001), as well as gastric-type secretory mucin expression, MUC5AC (Pereira et al. 2001; Matsukita et al. 2003), and MUC6 (De Bolos et al. 1998; Pereira et al. 2001; Matsukita et al. 2003). Considering the biological role of mucins, it has been suggested that the deregulated expression of apomucins could be significant in cancer biology (Turner et al. 2003; Baldus et al. 2004).
We recently found MUC5B expression in 16/31 primary breast tumors using a RT-PCR assay (Berois et al. 2003). The mRNA of this “non-mammary” apomucin was also found in MCF-7 but not in the four other breast cancer cell lines evaluated. We found MUC5B mRNA in 9/46 (19.5%) bone marrow aspirates obtained from patients who underwent curative surgery but not in 36 samples of normal peripheral blood mononuclear (PBMN) cells, indicating that MUC5B mRNA could be a specific marker applicable to the molecular diagnosis of breast cancer cell dissemination (Berois et al. 2003). The goal of the present study was to evaluate the expression profile of a MUC5B protein product in breast tissues, including breast cancers, normal tissues, and non-malignant lesions. We observed that MUC5B apomucin is over-expressed in a subset of breast cancer patients but was expressed very little or not at all in normal breast epithelium, suggesting that this apomucin is upregulated during the course of malignant transformation of the breast.
Materials and Methods
Cell Samples
Breast cancer cell lines MCF-7, T47D, and BT20 were purchased from American Type Culture Collection (Manassas, VA). These cell lines were cultured in DMEM medium supplemented with 10% bovine fetal serum, 2 mM
Tissues and Bone Marrow Aspirates
Paraffin-embedded breast tissues were obtained from the Department of Pathology, Pereira Rossell Hospital and the Department of Pathology, Casa de Galicia Clinic, Montevideo, Uruguay. Tissues were obtained from 42 patients with histopathological diagnosis of breast cancer and 14 patients with non-malignant breast diseases. All tumors were conventionally classified by histological type and grading was done according to the Scarff–Bloom–Richardson system with Nottingham modification (Frierson Jr et al. 1995). Normal breast tissues were obtained from reduction mammoplasties without malignant diseases. Tissue sections were applied onto silane-treated slides and used for histopathological diagnosis (stained with hematoxylin–eosin) and immunohistochemical assays. After informed consent, bone marrow aspirates were obtained from 22 patients with histological diagnosis of operable breast cancer at different stages. Our study was examined and approved by the Ethical Review Board of the Hospital de Clínicas, School of Medicine, Montevideo, Uruguay. All patients were screened for metastases by conventional staging (chest X-ray, liver ultrasound, blood tests, and bone scan in patients with stage II or III). Five-ml samples of bone marrow aspirates were collected by sternal and iliac crest punctures under anesthesia before surgery, in EDTA anticoagulant, and quickly sent to the laboratory.
In Situ Hybridization
In situ hybridization was performed on breast cancer cell lines using a MUC5B biotin-labeled oligonucleotide antisense probe (5′-TGTGGTCAGCTCTGTGAGGATC-CAGGTCGTCCCCGAGTGGAGAGGG-3′) (synthesized by Genset; Paris, France) specific for the tandem repeat domain of MUC5B (Dufosse et al. 1993). The procedure was based on a standardized protocol (Audie et al. 1993). After the hybridization step, samples were incubated with streptavidin–peroxidase. Color was developed using diaminobenzidine (DAB; Sigma, St Louis, MO) and H2O2.
Immunohistochemical and Immunocytochemical Analysis
Samples were dewaxed and rehydrated and epitopes were unmasked by boiling 20 min in 10 mM sodium citrate, pH 6.0. Sections were first incubated with normal goat serum (to decrease background staining), and then the anti-MUC5B antisera LUM5B-2, previously characterized by Wickström et al. (1998), was incubated overnight at 4C. This antibody was generated in rabbits immunized against a synthetic peptide RNREQVGKFKMC present in the cysteine-rich domains of the MUC5B apoprotein, flanking the highly glycosylated mucin regions. After three washes in PBS, quenching of endogenous peroxidase activity was performed with 3% H2O2 in PBS for 20 min. Sections were then incubated with 1/200 biotin-labeled goat anti-rabbit immunoglobulin (Sigma) for 1 hr at room temperature. Finally, peroxidase-labeled avidin (1/200) (Sigma) was added for 45 min. The reaction was revealed with DAB, and slides were counter-stained with Mayer's hematoxylin and mounted. For every assay, negative controls using PBS without primary antibody were included. The level of staining of each tumor was categorized as follows: 0, for negative samples or stained <10% of tumor extension; 1, for stained samples between 10 and 39%; 2, for stained samples between 40 and 79%; and 3, for tumors with >80% of stained cells. Signal intensity was assigned as strong (3), moderate (2), weak (1), and negative (0). Total score results from the combination of both parameters.
Anti-MUC5B antibody was also evaluated by immunocytochemistry on three human breast cancer cell lines (MCF-7, T47D, and BT-20). Suspended cells were spun onto glass slides using 100-μl aliquots at 1 × 106 cells/ml (Shandon; Runcorn, UK) and fixed using 50% methanol/acetone (v/v). After incubation with LUM5B-2 antisera, the immunostaining protocol performed was as described for immunohistochemistry.
Preparation of Cell Lysates and Immunoblotting
Cell lysates were prepared in 10 mM Tris-HCl (pH 7.9), 150 mM NaCl, 1% NP40, and 1 mM phenylmethylsulfonyl fluoride, and nuclei were removed by centrifugation at 5000 X g for 5 min at 4C. Protein content was determined by the bicinchoninic acid method (BCA; Sigma) as recommended by the supplier. Cell lysates were resolved on SDS-polyacrylamide gel electrophoresis (Laemmli 1970), performed using a 3–10% gradient gel under reducing conditions. Proteins were transferred to nitrocellulose sheets (Amersham; Aylesbury, UK) according to Towbin and coworkers (1979) at 60 V for 5 hr in 20 mM Tris-HCl, pH 8.3, 192 mM glycine, and 10% methanol. Residual protein-binding sites were blocked by incubation with 3% bovine serum albumin (BSA) in PBS overnight at 4C. The nitrocellulose was then incubated with the anti-MUC5B antisera LUM5B-2 for 3 hr at 37C. After three washes with PBS containing 0.1% Tween 20 and 1% BSA, the membrane was incubated for 1 hr at room temperature with goat anti-rabbit immunoglobulin conjugated to peroxidase (Sigma) diluted in PBS 0.3% Tween 20 and 3% BSA. Reactions were developed by the enhanced chemoluminescence (ECL) method (Amersham) according to the manufacturer's directions. The same procedure was performed omitting antisera LUM5B-2 as a negative control.
RT-PCR Assay for MUC5B in Bone Marrow Aspirates
Bone marrow aspirates were first centrifuged at 1500 X g for 5 min and the buffy coats removed into fresh tubes. The contaminating red blood cells were eliminated by lysis buffer (154 mM ammonium chloride, 12 mM sodium bicarbonate, and 0.1 mM EDTA) under gentle agitation for 10 min. Mononuclear cells were recovered by centrifugation at 1500 X g for 5 min. Total RNA was extracted with Tri-Reagent (Sigma) from bone marrow aspirates, according to the manufacturer's instructions. First-strand cDNA was synthesized by reverse transcriptase (MMLV; Amersham) using random hexamers. MUC5B mRNA was identified using a nested RT-PCR reaction as described elsewhere (Berois et al. 2003), and β-actin RT-PCR assays were performed to control RNA integrity and cDNA synthesis in each sample. Ten μl of all PCR products were analyzed by electrophoresis on 2% agarose gels by direct visualization after ethidium bromide staining.
Statistical Analysis
Correlation between MUC5B expression and various clinicopathologic factors were analyzed by using Fisher's exact test. Univariate analysis of disease-free survival was performed using the log-rank test; p values <0.05 were considered to be statistically significant.
Results
MUC5B Expression in Breast Cancer Cell Lines
Considering that previously we detected MUC5B mRNA in MCF-7 cells but not in other breast cancer cell lines (Berois et al. 2003), we performed an immunocytochemical evaluation of these cell lines using the anti-MUC5B antibody LUM5B-2. We detected the MUC5B protein in MCF-7 cells (Figure 1), but not in T47D or BT20 cells (data not shown). The staining pattern corresponded to a cytoplasmic and cell surface localization. Expression of MUC5B apomucin in MCF-7 breast cancer cells was also confirmed by in situ hybridization, using a probe specific for the MUC5B tandem repeat (Figure 1). Western blot analysis on MCF-7 cell extracts showed that MUC5B migrated as a broad major band of ∼400 kDa and several components of higher molecular weight (part marginally entering in the separation gel) (Figure 1).
MUC5B Expression in Primary Breast Cancer
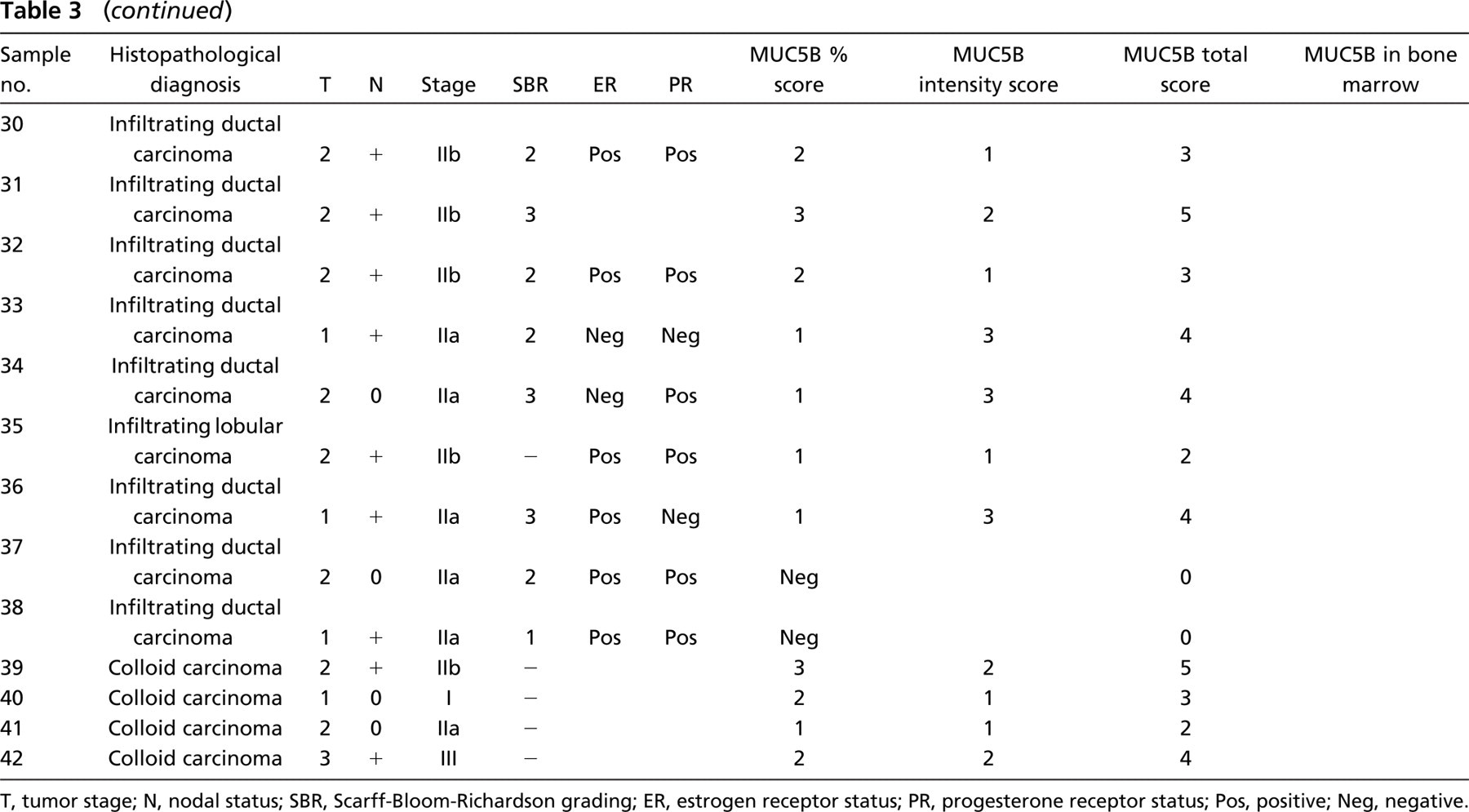
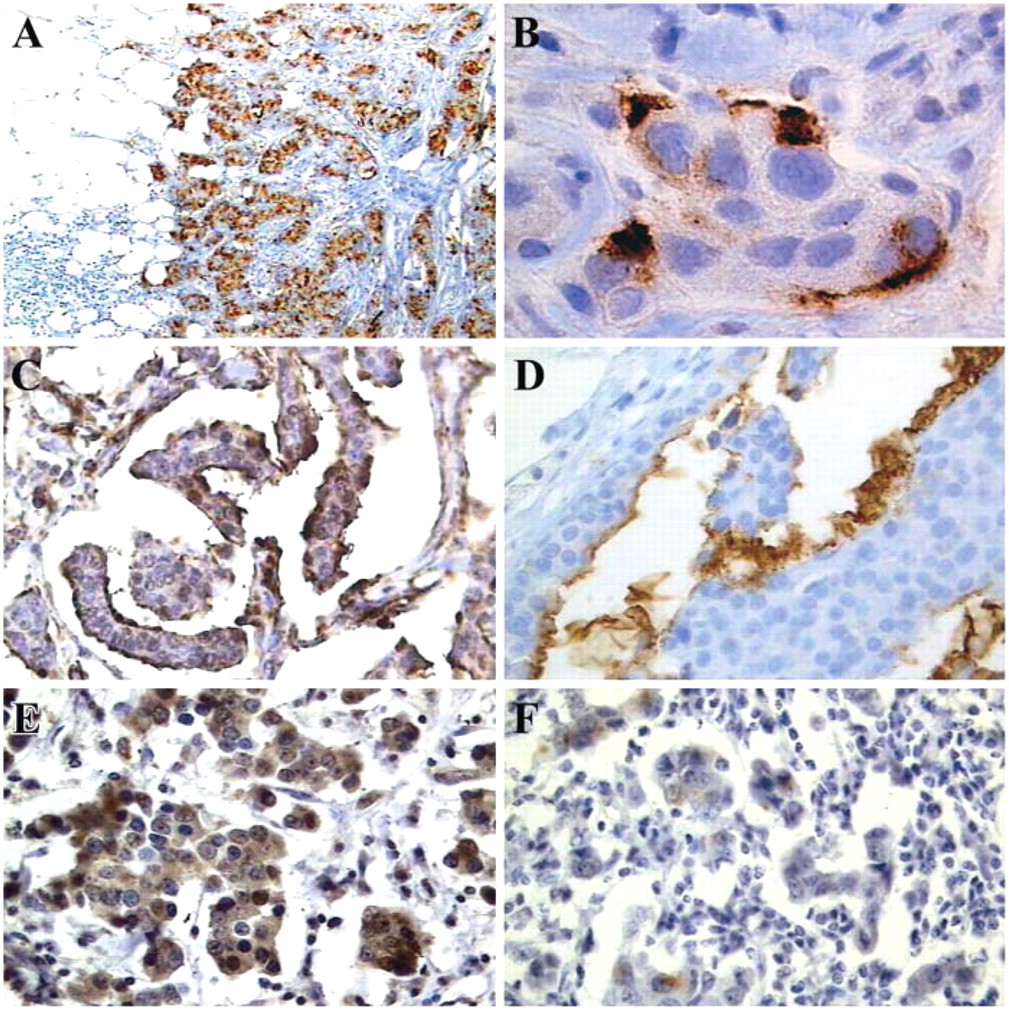
MUC5B apomucin expression was evaluated in 42 primary tumor specimens from patients with breast cancer. Characteristics of patients and tumors are indicated in Table 1. A positive staining was observed in 34/42 (81%) specimens (Table 2) with variations in the staining percentage and intensity (Table 3). In infiltrating ductal carcinoma the staining pattern was preponderantly cytoplasmic and perinuclear (Figures 2A and 2B), whereas in 4/4 colloid carcinomas the signal was predominantly apical and in the nearest secretory material from tumoral cells (Figure 2). MUC5B apomucin was strongly expressed in 2/2 invasive papillary carcinomas and was detected in 1/2 invasive lobular carcinomas evaluated. The relationship between MUC5B expression in these samples and some of the different clinicopathologic parameters such as tumor size, lymph node involvement, tumor differentiation, and hormone receptor status is shown in Figure 3. MUC5B was expressed more frequently in T1 tumors (11/12; 91.7%) rather than in T2–T3 tumors (23/30; 76.7%), and in tumors from patients N0 (15/17; 88.2%) rather than in those with metastatic lymph nodes (19/24; 79.1%). Differences were not statistically significant. Regarding stage of disease, MUC5B was expressed in 6/6 (100%) tumors from stage I patients, in 14/17 (82.3%) stage IIa, in 13/16 (81.2%) stage IIb, and in 1/2 stage III. No correlation was observed between MUC5B expression and tumor grading or with estrogen or progesterone receptors.
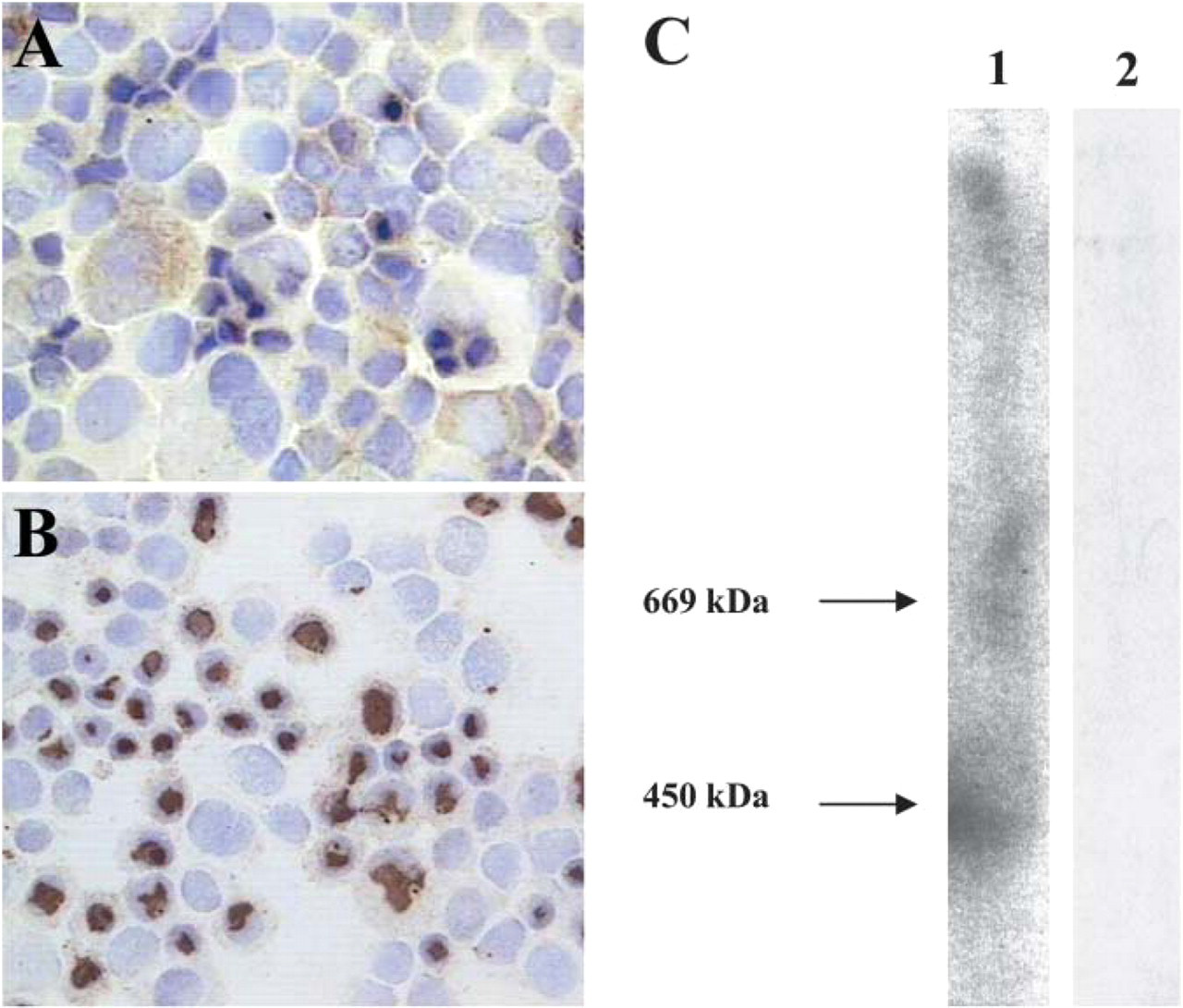
MUC5B expression in the human breast cancer MCF-7 cell line. (
Adjacent ductal carcinoma in situ (DCIS) was observed in 13/42 cases, and 7 showed MUC5B immunoreactivity. In these cases the staining was predominantly cytoplasmic with an apical pattern (Figure 2D). Immunoreactivity was also observed in the secretory material. Interestingly, we observed a total correspondence between DCIS and the invasive form. All seven MUC5B-positive DCIS samples were present in tissues corresponding to MUC5B-positive breast carcinomas, and all six MUC5B-negative DCIS samples corresponded to MUC5B-negative breast carcinomas.
The previous observation that MUC5B mRNA is expressed by breast cancer cells and not at all by PBMN cells (Berois et al. 2003) led us to test the immunohistochemical expression of this marker in regional lymph nodes obtained from 10 patients with breast cancer, evaluating whether there is a correlation with the respective primary tumor. Lymph nodes without tumor involvement showed no expression of MUC5B, even when the primary tumors had a strong signal for this protein. In contrast, metastatic lymph nodes (by conventional histology) were clearly identified using the anti-MUC5B antibody only when the primary tumor was also positive for MUC5B. In most samples a stronger staining pattern was observed in the tumor compared with the respective lymph nodes (Figures 2E and 2F).
MUC5B Expression in Normal Breast and Non-malignant Breast Diseases
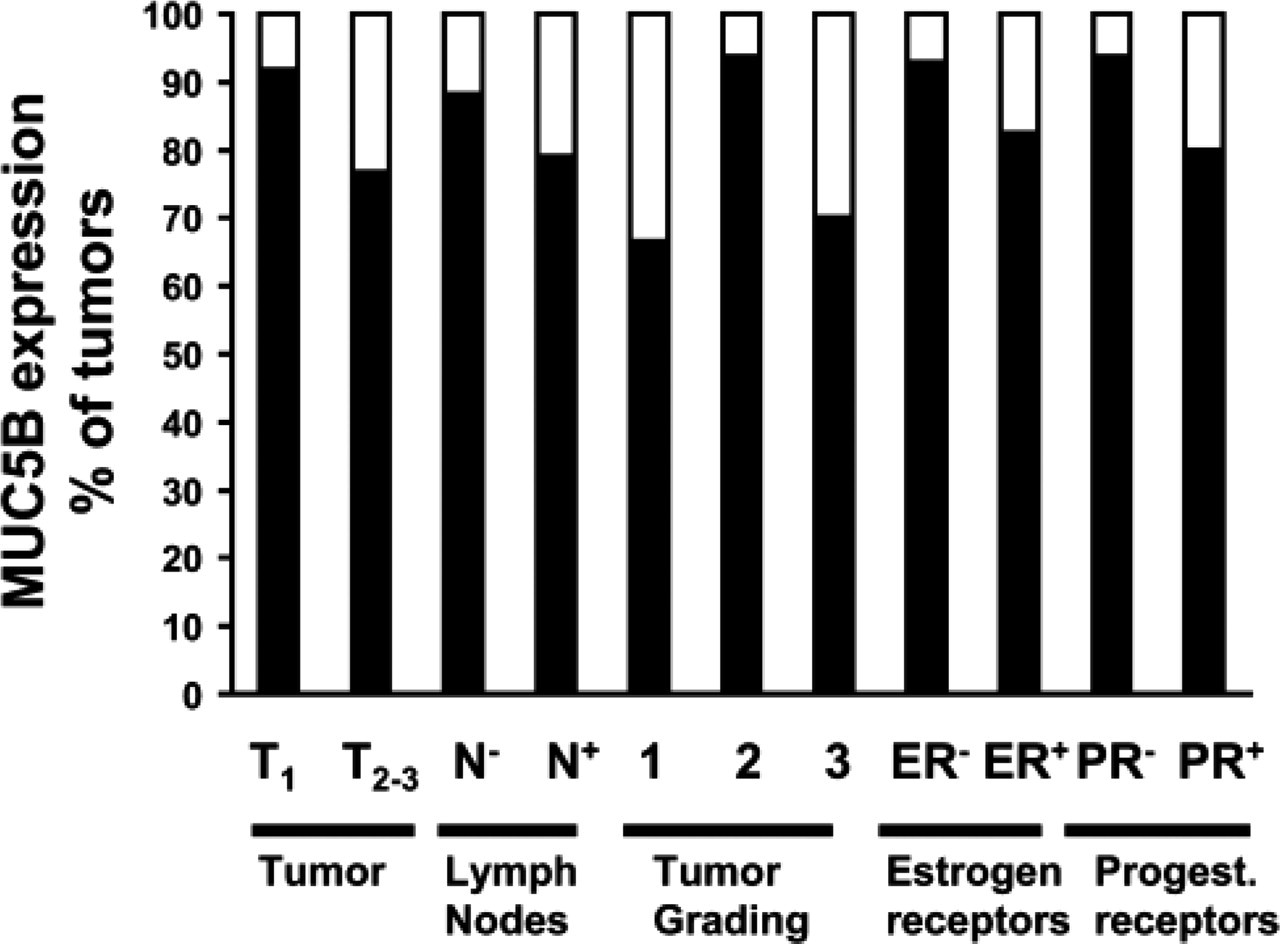
Analysis of five normal breast samples showed no staining or a weak signal in a small cell population (<5%) of some samples. In these cases an apical staining pattern of the normal lobules was observed with positivity in the secreted material (Figure 4). Normal-appearing epithelia adjacent to breast cancer was evaluated in 19/42 specimens. In 11/19 (57.9%) cases, non-malignant cells were negative; in 3/19 (15.8%) samples, MUC5B was focally expressed with weaker intensity relative to adjacent malignant epithelia; and in 5/19 (26.3%) cases, staining was also observed in almost all non-malignant cells adjacent to cancer. In these samples the staining pattern was predominantly apical and in the secretory material, but in some cases the staining was also observed at the cytoplasm.
Characteristics of the patients and tumors
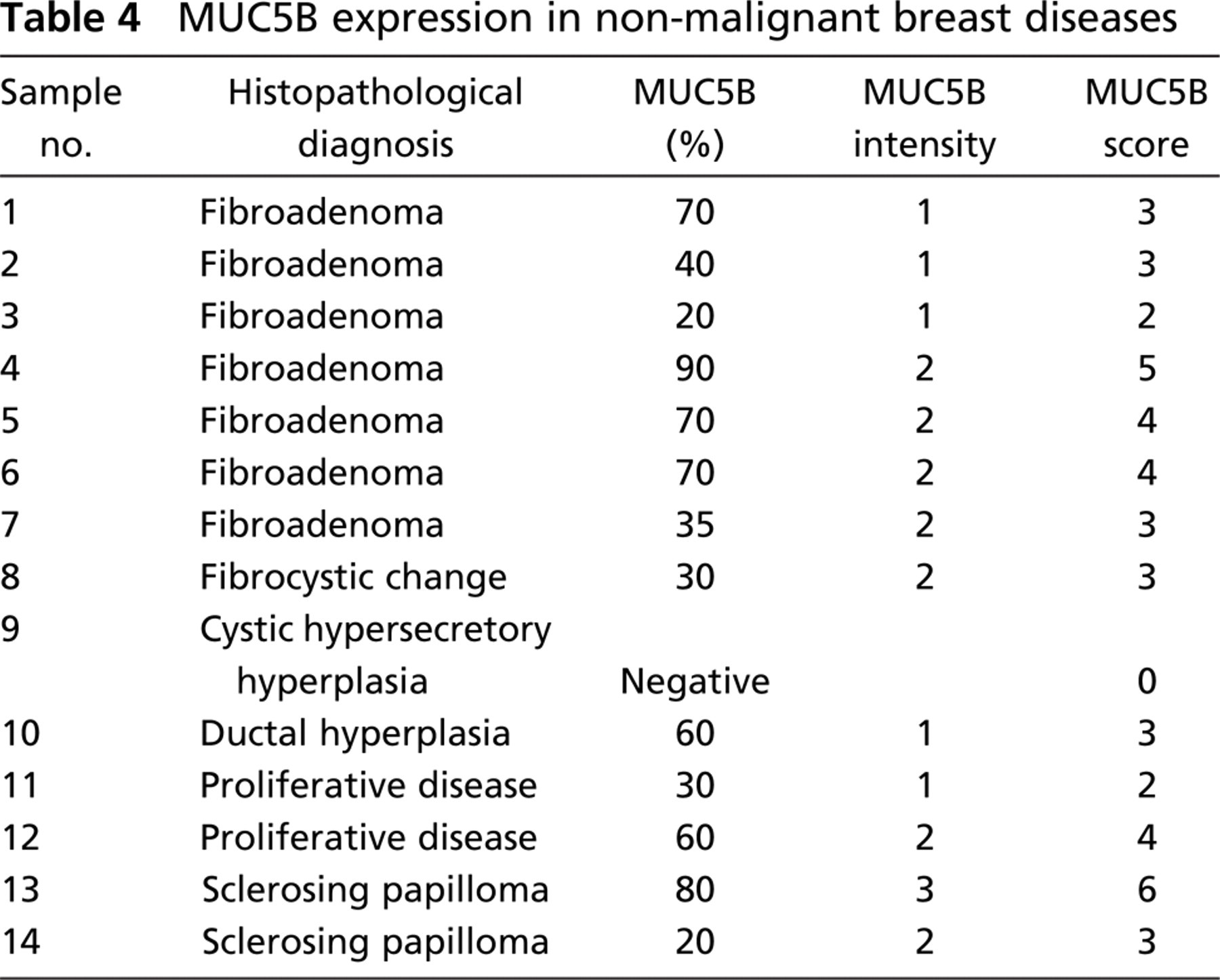
Regarding non-malignant breast diseases, MUC5B expression was observed in almost all (13/14; 92.8%) samples (Table 4), although the staining pattern was different compared with that observed in breast carcinomas. In common hyperplasia we found the same pattern as in normal lobules (Figure 4). In fibroadenomas the reactivity was seen especially in the ducts, whereas normal lobules were negative or showed a weak signal (Figure 4). MUC5B immunoreactivity was not observed in cystic hypersecretory hyperplasia with lactational changes (Figure 4D).
Correlation between MUC5B Expression in Tumors and Bone Marrow Samples
Considering that MUC5B mRNA could be detected in disseminated breast cancer cells using a RT-PCR assay (Berois et al. 2003), we compared mRNA expression in bone marrow with MUC5B staining in breast primary tumors. We evaluated bone marrow aspirates from 22/42 patients with operable breast cancer (samples 1 to 22 of Table 3). We found 6/22 (27.3%) bone marrow samples positive for MUC5B, and all also displayed MUC5B expression in the primary tumor. In 6/22 (27.3%) patients, both the tumor and bone marrow were negative for MUC5B expression, whereas 10/22 (45.4%) patients had positive tumor and negative bone marrow.
Discussion
The regulation of mucin expression is tissue specific, both in mucin core-peptide expression and in glycosylation. An abnormal expression of the MUC genes has been demonstrated in several malignant diseases (Ho et al. 1993; Kim et al. 1996). In the immunohistochemistry-based study reported here using LUM5B-2 antisera raised against sequences within the non-glycosylated regions of MUC5B (Wickström et al. 1998), we found this apomucin in 81% of breast cancer tissues. Antibodies specific for this region have an advantage: they recognize the precursor as well as the mature, fully O-glycosylated mucin (van Klinken et al. 1998b). MUC5B characterization could be improved by the use of a specific monoclonal antibody (Rousseau et al. 2003), which has been recently used in immunohistological studies in carcinoma of other organs (Pinto de Sousa et al. 2004). The expression of MUC5B apomucin by breast cancer cells was confirmed using immunocytochemistry, in situ hybridization, and Western blot on MCF-7 cancer cells. Mammary tumorigenesis is a multistep process. MUC5B was detected in 8/19 (42.1%) samples of normal-appearing breast epithelia adjacent to cancer but in 0/5 normal control breast samples. This fact, as well as the observation of total correspondence between adjacent DCIS and the invasive form in the MUC5B immunoreactivity, suggests that MUC5B expression could be an early event during human breast carcinogenesis. These results agree with the observation that some mucin-type O-glycosylated antigens are early biomarkers of malignant transformation in breast epithelium prior to histopathological detection of mammary carcinoma, both in humans (Cho et al. 1994) and in animal models (Babino et al. 2000). MUC5B apomucin was present in most benign breast diseases, although the staining pattern was very different compared with that observed in breast carcinomas (where MUC5B displayed cytoplasmic and non-apical membrane distribution). These results are similar to those observed for MUC1 expression pattern in breast cancer compared with non-malignant cells (Vlad et al. 2004). Taking into account that mucins are normally present at the cell surface playing an important role in cell adhesion, several questions could be raised at the functional level regarding the cytoplasmic distribution of some mucins in cancer cells. It was recently demonstrated that MUC1 binds directly to p53 and promotes p53-dependent cell fate selection of the growth arrest and survival responses to genotoxic anticancer agents (Wei et al. 2005).
Analysis of MUC5B a expression in human breast tissues
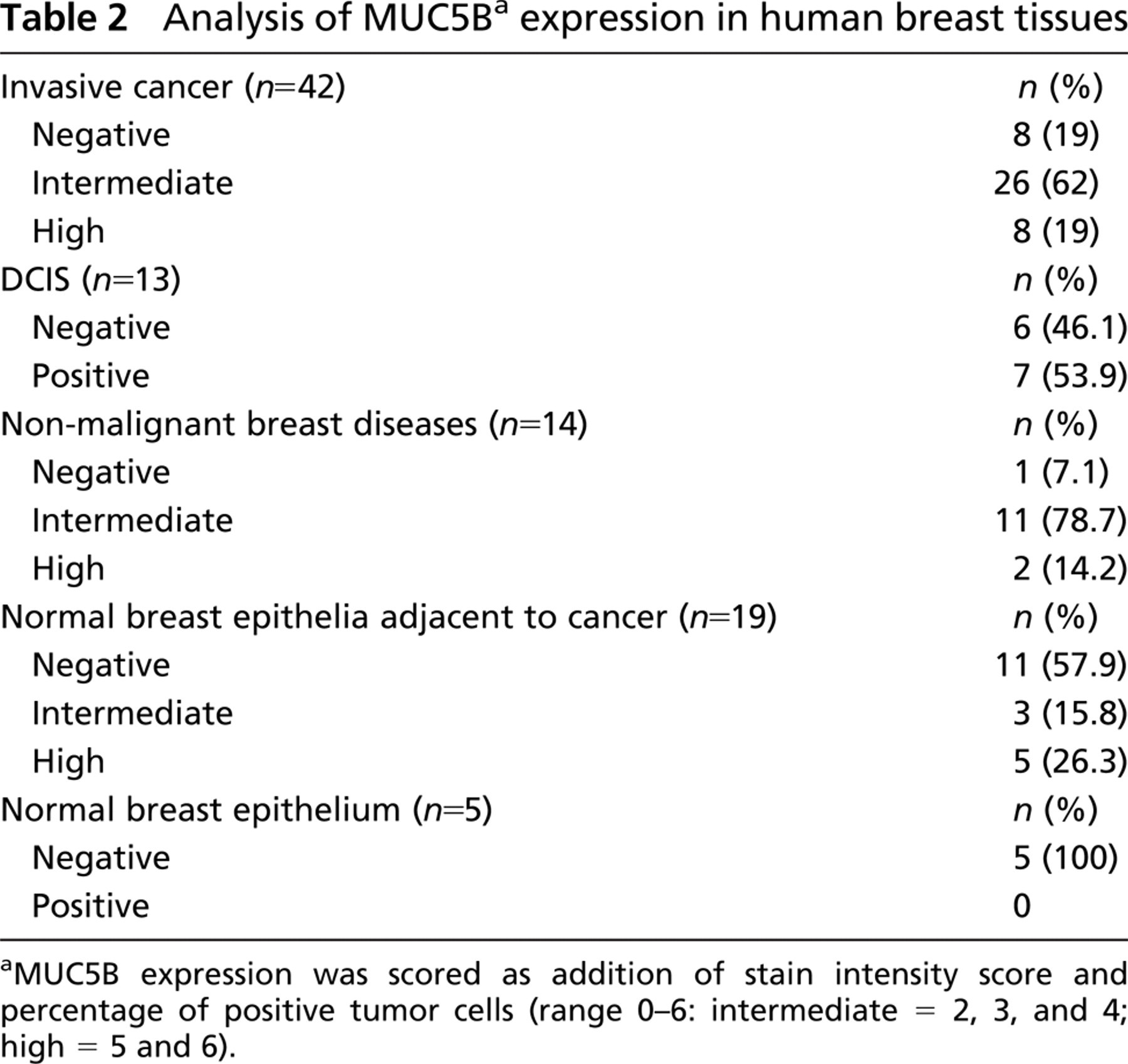
aMUC5B expression was scored as addition of stain intensity score and percentage of positive tumor cells (range 0–6: intermediate = 2, 3, and 4; high = 5 and 6).
Relationship between MUC5B expression and various clinicopathological factors
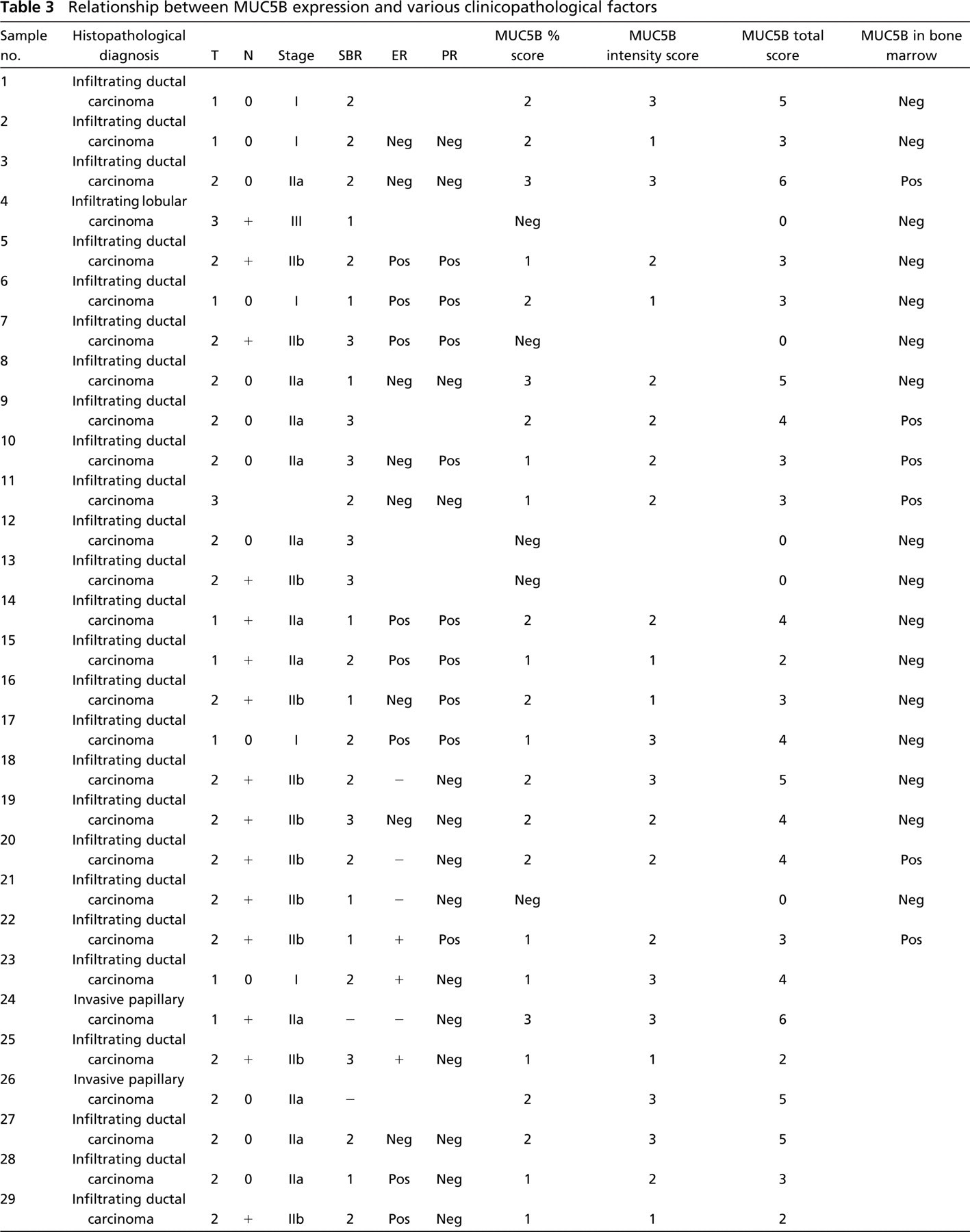
T, tumor stage; N, nodal status; SBR, Scarff-Bloom-Richardson grading; ER, estrogen receptor status; PR, progesterone receptor status; Pos, positive; Neg, negative.
MUC5B mucin is expressed mainly in mucous cells of airway submucosal glands (Sharma et al. 1998), salivary glands (Audie et al. 1993), gallbladder (van Klinken et al. 1998a), endocervix (Gipson et al. 1999), pancreas (Balague et al. 1994), and submucosal glands of the esophagus (Guillem et al. 2000). In cancer, MUC5B has been shown to be aberrantly expressed in lung adenocarcinomas (Yu et al. 1996; Copin et al. 2001), in gastric carcinomas (Buisine et al. 2000; Pinto de Sousa et al. 2004), and in HT-29 colon carcinoma cells resistant to methotrexate (Leteurtre et al. 2004). Human MUC5B gene is mapped clustered with MUC6, MUC2, and MUC5AC to chromosome 11p15.5 (Pigny et al. 1996). They encode large secreted gel-forming mucins that contain transforming growth factor-β-like domains in their C-terminal end (Porchet et al. 1999). They have evolved from a common ancestor gene (Desseyn et al. 1998) and have some common regulatory mechanisms (van Seuningen et al. 2001). Interestingly, in past years it has been observed that all of these “non-mammary” mucins are expressed in breast cancer (De Bolos et al. 1998; Diaz et al. 2001; Pereira et al. 2001; Matsukita et al. 2003).
Our results show no statistically significant correlation between MUC5B apomucin expression and tumor size, nodal status, histologic grade, or hormone receptors status. To extend our observations it is now important to evaluate a larger tumor collection of breast cancers and to determine its prognostic significance by patient survival evaluation. We observed a very good correlation between MUC5B protein expression in primary tumors and MUC5B mRNA in bone marrow samples of the same patients. These results support the usefulness of MUC5B for disseminated breast cancer cell detection in bone marrow samples. Recently, we have observed that operable breast cancer patients expressing MUC5B mRNA in bone marrow samples had a favorable clinical outcome (Varangot et al. 2005). This fact agrees with our preliminary observation that this apomucin was expressed with lower intensity in metastatic lymph nodes than in primary tumors, suggesting that MUC5B could be associated with a less aggressive subpopulation of breast cancer cells. Scarce data are available about the role of MUC5B in cancer biology. It was reported that its expression was associated with an absence of venous invasion in gastric carcinoma (Pinto de Sousa et al. 2004). MUC5B expression could be responsible for a phenotype related with a better outcome in breast cancer. A detailed characterization of this non-mammary apomucin is in progress to determine its role in breast cancer biology.
Immunohistochemical evaluation of MUC5B apomucin in breast cancer. (
Relationship between MUC5B expression and different clinicopathologic parameters.
For the elucidation of molecular mechanisms that control mucin gene expression in breast cancer, it is imperative to identify transcription factors that target MUC genes during malignant transformation. It was demonstrated that MUC5B expression in gastric cancer cells is governed by a highly active distal promoter that is upregulated by protein kinase C and that repression is under the influence of methylation (Perrais et al. 2001). The fact that MUC5B is expressed in MCF-7 but not in other cell lines analyzed indicates that these cells could be used as a model for studying expression of this gene. It would be very interesting to analyze MUC5B expression in this cell line under hormonal influence, like progesterone and estrogens, because MCF-7 has both kinds of receptors. Concerning MUC5B-negative cell lines, BT20 does not possess hormone receptors, and T47D, even if it has receptors, is less sensitive than MCF-7 to hormones. In recent years it has been determined that MUC5B promoter has potential estrogen and progesterone binding sites (Perrais et al. 2001), and hormonal regulation of MUC5B expression has been observed in the endocervix (Gipson et al. 1999). The other members of the 11p15.5 cluster are also hormonally regulated (Freedman and Luisi 1993; Gollub et al. 1995; De Bolos et al. 1998). However, in the preliminary evaluation performed here on 31 patients with breast cancer, we did not observe any correlation between MUC5B expression and estrogen or progesterone receptor status.
Immunohistochemical evaluation of MUC5B apomucin in non-malignant breast tissues. (
MUC5B from human saliva, respiratory tract secretions, and cervical pregnancy mucus was shown to be a large oligomeric mucin composed of subunits linked by disulfide bonds. Although the apoprotein is the same, there are MUC5B glycoforms that are produced by different populations of cells (Wickström et al. 1998). In airway diseases there are significant increases in low-charge forms of the MUC5B mucin in cystic fibrosis and chronic obstructive pulmonary disease (Kirkham et al. 2002). The results presented in our work show that the heterogeneous electrophoretic behavior of MUC5B from breast cancer cells is very similar to that observed for this mucin in colon cancer cells (Leteurtre et al. 2004). Although it is unknown whether MUC5B glycosylation is modified by malignant transformation, we can speculate that aberrant expression of MUC5B apomucin by breast cancer cells could lead to the production of other MUC5B glycoforms, probably less glycosylated (as suggested by our Western blot results), which have different functional properties. It is possible that the aberrant expression of a specific type of apomucin could lead to the accumulation of simple O-glycan tumor-associated antigens such as Tn. Evaluating soluble apomucins in breast cancer pleural effusions, we observed that Tn is expressed not only on MUC1, the major breast mucin, but also on the other three non-mammary apomucins evaluated (MUC2, MUC5AC, and MUC6) (Freire et al. 2003). Peptides based on the sequence of the MUC6 tandem repeat have been GalNAc-glycosylated in vitro using ppGalNAc-Ts (Reis et al. 2000). Recently, we demonstrated that MUC6 apomucin expression could be responsible, at least in part, for Tn antigen expression in MCF-7 breast cancer cells (Freire et al. 2005). This observation could be explained by two results: (a) an amino acid sequence derived from MUC6 was a very good acceptor substrate for ppGalNAc-Ts (which are the enzymes catalyzing the Tn antigen synthesis) and (b) the MUC6-Tn glycopeptide was a poor acceptor substrate for core 1 β3Gal-T, the next enzyme involved in the saccharide chain biosynthesis (Freire et al. 2005).
MUC5B expression in non-malignant breast diseases
In this first evaluation performed at the protein level of MUC5B in breast tissues, we observed that this apomucin is overexpressed in 81% of breast cancers but is generally undetected in normal breast epithelium. The association of MUC5B expression with breast cancers at a lower stage remains to be confirmed in a larger population study. Considering the important role of mucins in invasiveness and metastasis, as well as in modulation of immune response against cancer, it will be important to determine the relationship between MUC5B expression and breast cancer biology.
Footnotes
Acknowledgements
This work was partially supported by ECOS France–Uruguay Program.
We thank Dr. I. Carlstedt, who kindly provided the LUM5B-2 antibody; Dr. C. Pressa and Dr. R. Laviña (Casa de Galicia Clinic, Montevideo, Uruguay), who obtained bone marrow aspirates from operable breast cancer patients; and Dr. C. Herrera for her helpful contribution.