
Abstract
In this study we investigated the expression of bone morphogenetic protein (BMP)-signaling Smads in distraction osteogenesis (DO). Osteotomy of the right tibia was performed in 14 skeletally mature white New Zealand male rabbits. Lengthening was started 1 week later at a rate of 0.5 mm/12 hr and was maintained for 3 weeks. Expression of Smad proteins 1, 4, 5, 6, 7, and 8 and Smad ubiquitin regulatory factors (Smurfs) 1 and 2 was evaluated in the distracted zone using immunohistochemistry. Expression of receptor-regulated Smads (R-Smads) 1, 5, and 8 showed a significant increase during the distraction phase, followed by a gradual decrease during the consolidation phase. Smad 4 showed significant expression during both distraction and the beginning of the consolidation phase. Smad 6 and Smad 7 were highly expressed during the consolidation phase. Staining for both Smurfs 1 and 2 was maximal at the end of the distraction period. Staining for all proteins was most intense in chondrocyte and fibroblast-like cells. Expression pattern of R-Smads correlated with our previously reported expression pattern of BMPs 2, 4, and 7 and their receptors. These results therefore suggest a role for the whole BMP signaling pathway including the Smad proteins in DO.
D
Smad proteins are major intracellular signaling molecules in the BMP-induced signaling pathway (Kawa-bata et al. 1998; Canalis et al. 2003; Shi and Massague 2003; Nohe et al. 2004). They are classified into three subclasses according to their structure and mechanism of action, which include receptor-regulated Smads (R-Smads 1, 5, and 8), common-partner Smad (Co-Smad 4), and inhibitory Smads (I-Smads 6 and 7). R-Smads are activated by BMP receptors that allow heterodimerization with the CoSmad, Smad 4. I-Smads negatively regulate BMP signaling by competing with R-Smads for activated type I receptor via Smad 6 and Smad 7, by competing with R-Smads for interaction with Smad 4 via Smad 6, or by targeting BMP receptors for their degradation (Hayashi et al. 1997; Hata et al. 1998; Ishida et al. 2000; Shi and Massague 2003; Park 2005).
R-Smads are degraded by the ubiquitin-proteasome pathway (Miyazono 2000; Shi and Massague 2003). Smad ubiquitin regulatory factors (Smurfs) are implicated in the ubiquitin-mediated degradation of different components of the BMP signaling pathway (Zhu et al. 1999; Kavsak et al. 2000). They can bind to R-Smads and promote their degradation, or they can target activated type I receptors through I-Smad (Smad 7) and enhance receptor turnover (Arora and Warrior 2001).
Although Smads expression during embryological bone formation and fracture healing has been extensively investigated (Sakou et al. 1999; Kloen et al. 2002, 2003; Yu et al. 2002), there has been only one report on Smads expression (Smads 1-5) in mandibular DO (Farhadieh et al. 2004). The aim of this research was to analyze the temporal and spatial expression of Smads and Smurfs in DO and to try to correlate this expression with that of BMP ligands and BMP receptors.
Materials and Methods
Fourteen skeletally mature, 9-month-old male white New Zealand rabbits weighing 2.5-3.5 kg were used. Housing, care, and experimental protocol were approved by the McGill University Animal Care and Ethics Committee.
Operative Protocol
Operative and distraction protocols were identical to our earlier studies (Rauch et al. 2000; Hamdy et al. 2003). Rabbits were anesthetized by intramuscular administration of ketamine, xylazine, and acepromazine. Anesthesia was maintained with halothane, oxygen, and nitric oxide after intubation. A modified Orthofix uniplanar M-100 fixator (Orthofix; Verona, Italy) was applied to the right tibia under sterile conditions. The tibia was exposed subperiosteally, and the osteotomy was performed with an oscillating saw just below the fusion site between tibia and fibula. The periosteum was reapproximated and the wound closed. Unrestricted weight bearing and activity were allowed postoperatively. After a delay of 7 days, distraction was started at a rate of 0.5 mm/12 h for 3 weeks. This was followed by a period of consolidation during which the external fixator was held in place with no distraction. Rabbits were examined daily for signs of infection, weight loss, and pain.
Starting 1 week after surgery, two rabbits were sacrificed every week for 7 weeks postoperatively. Rabbits were sacrificed by intravenous injection of Euthanyl (MTC Pharmaceuticals; Cambridge, ON, Canada).
Sample Preparation
Following sacrifice, the external fixator was removed and the right tibia was dissected free from soft tissues. Specimens were fixed in 4% paraformaldehyde overnight, decalcified in 20% ethylenediaminetetraacetic acid for 3 weeks, and then embedded in paraffin. Seven-um-thick sections were cut. Parallel sections were taken so that both temporal and spatial expression of different Smads were evaluated and compared with each other. After deparaffinization and hydration, endogenous peroxidase was blocked with 1% hydrogen peroxide for 10 min. Nonspecific binding was blocked by incubation in 10% normal horse serum (NHS) (Vector Laboratories; Burlingame, CA) for 10 min in a humidified chamber. For immunostaining, the following antibodies were tested: Smads 1, 4, 5, 6, 7, and 8, Smurf 1 and Smurf 2 (Santa Cruz Biotechnologies; Santa Cruz, CA). Sections were incubated with these primary antibodies in 1% NHS using a 1:40 dilution for 1 hr in a humidified chamber. Tissue sections were subsequently incubated for 1 hr with a secondary antibody (biotinylated antigoat antibody) in a 1:100 dilution (Vector Laboratories). Sections were then stained using the avidinbiotin complex method (Vector Laboratories) and 3,3′-diaminobenzidine tetrachloride. Finally, sections were counterstained with hematoxylin and mounted with Permount. For negative controls, the same procedure was followed except the primary antibodies were omitted.
Peptide Neutralization
According to data provided by the manufacturer (Santa Cruz Biotechnologies), primary antibodies used in the present study recognize mouse, rat, and human Smad and Smurf proteins. Therefore, it was necessary to determine if these antibodies also recognized specific rabbit Smads and Smurf proteins. Blocking peptides are available for all Santa Cruz Biotechnology affinity-purified rabbit and goat polyclonal antibodies and monoclonal antibodies raised against peptide antigens. Antibodies binding to antigens may be blocked/competed by preabsorption with the blocking peptide. To perform a blocking/competition process, we combined 1 μl of the primary antibody (200 μg/ml) with a 5-fold (w/w) excess of its respective blocking peptide (200 μg/ml) in a small volume of phosphate-buffered saline totaling 500 μl. This was then incubated overnight at 4C. The following morning, the immunohistochemistry protocol was followed using the same 1:40 dilution in NHS and following the same protocol described above. Treated samples showed no evidence of staining (Figure 1), thus confirming that the antibodies used in the present study were specific to the rabbit protein of interest. This technique has been previously published by the authors (Campisi et al 2003; Hamdy et al 2003).
Grading of the Immunostained Sections
Traditionally, immunohistochemistry is a qualitative technique used for the anatomical identification of the different cellular and extracellular components that stained positively. However, similar to previously published reports (Tavakoli et al. 1999; Yeung et al. 2001), we have developed in our laboratory a special method to evaluate semiquantitatively our results, based on the percentage of cells showing a positive staining. Our results with this technique have been previously published for long bone (Rauch et al 2000; Hamdy et al. 2003) and mandibular DO (Campisi et al. 2003). We have used the same technique in the present study. Each of the immunostained sections was graded blinded, the scores for intensity and percentage of cells stained being as follows: - represents no staining in the majority of cells; + represents staining in less than 25% of cells; ++ represents staining in 25-50% of cells; +++ represents staining in 50-75% of cells; ++++ represents staining in more than 75% of cells. The number of cells expressing the various proteins is assessed by cell counting. Chondrocytes, osteoblastic, and fibroblastic cells are identified morphologically. These analyses are performed separately for the callus region and the center region containing the fibrous interzone. Figure 2 provides a graphical representation of how immunohistochemistry images are evaluated semiquantitatively.
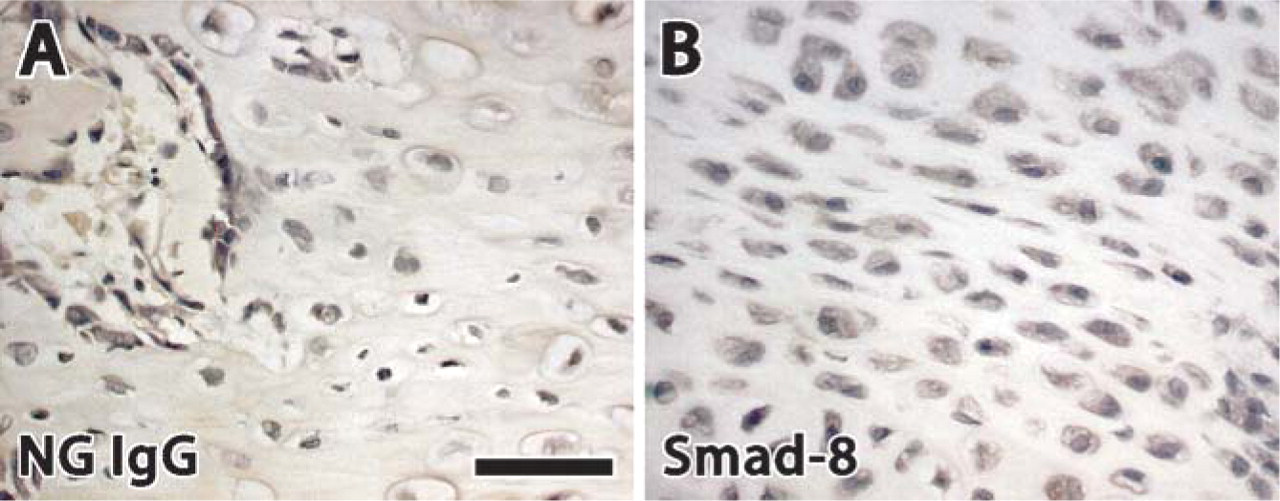
This image verifies antibody specificity for rabbit antigens. (A) Normal goat IgG. (B) Blocking peptide for Smad 8. The absence of staining indicates that the primary antibody is specific to reacting with the protein of interest. Bar = 50 μm.
Results
Expression of Smads 1, 4, 5, 6, 7, and 8 and Smurfs 1 and 2 was studied at weekly intervals. Tables 1 and 2 summarize the results obtained from immunohisto-chemistry. Distraction was started at week 1, and the consolidation phase began at week 4. Figures 3–6 provide representative examples of immunostained sections.

Schematic representation of semi-quantitative evaluation of immunohistochemistry images. (
Among the receptor-activated Smads, Smad 1 and Smad 5 exhibited a similar level of expression in chon-drocyte and fibroblast cells, staining intensely during the distraction period and less intensely 1 week after distraction, whereas Smad 8 showed less expression at all time points.
Smad 4 was highly expressed during distraction and at the beginning of the consolidation phase in both chondrocytes and fibroblasts, at the center and in the callus region. During the last 2 weeks of consolidation, about half of the chondrocytes still stained for Smad 4, but there was no positive staining in fibroblasts.
Inhibitory Smads, i.e., Smad 6 and Smad 7, showed increasing staining as the distraction progressed in both chondrocytes and fibroblasts, with the highest level of expression at the end of distraction and at the beginning of consolidation. Later in the consolidation phase, Smad 6 expression decreased slightly in chondrocytes and decreased abruptly in fibroblasts. In contrast, in the same period Smad 7 manifested a high level of expression in chondrocytes but was also absent in fibroblasts. Smad immunostaining revealed a low signal in osteoblastic cells in both center and callus regions, with the exception of I-Smads (Smad 6 and Smad 7) that were more intensely expressed in these areas during distraction and consolidation phases.
Smurfs 1 and 2 were maximally expressed in chondrocytes. Staining was at its maximum at the end of distraction and at the beginning of consolidation.
Discussion
In previous studies, we analyzed the expression of BMP-2, −4, and −7, as well as the expression of BMP receptor proteins during DO (Rauch et al 2000; Hamdy et al 2003). We found that the expression of BMP-2, −4, and −7 was maximal during the distraction phase of DO, but their expression tapered off during the consolidation phase. Similarly, the most intense staining for BMP receptors was evident during the second week of distraction and the expression level gradually diminished toward the end of distraction and during the consolidation phase. Results of these studies suggested that BMPs may play an important role in the signaling pathways that link the mechanical forces created by distraction to biological responses. In the present study, we investigated the expression pattern of the major intracellular signaling pathway proteins of BMP, i.e., the expression of Smad proteins. Although the expression of these proteins during embryological bone formation and fracture repair has been extensively investigated, their expression in DO remains largely unknown.
Immunohistochemistry results of R-Smads and CoSmad protein expression
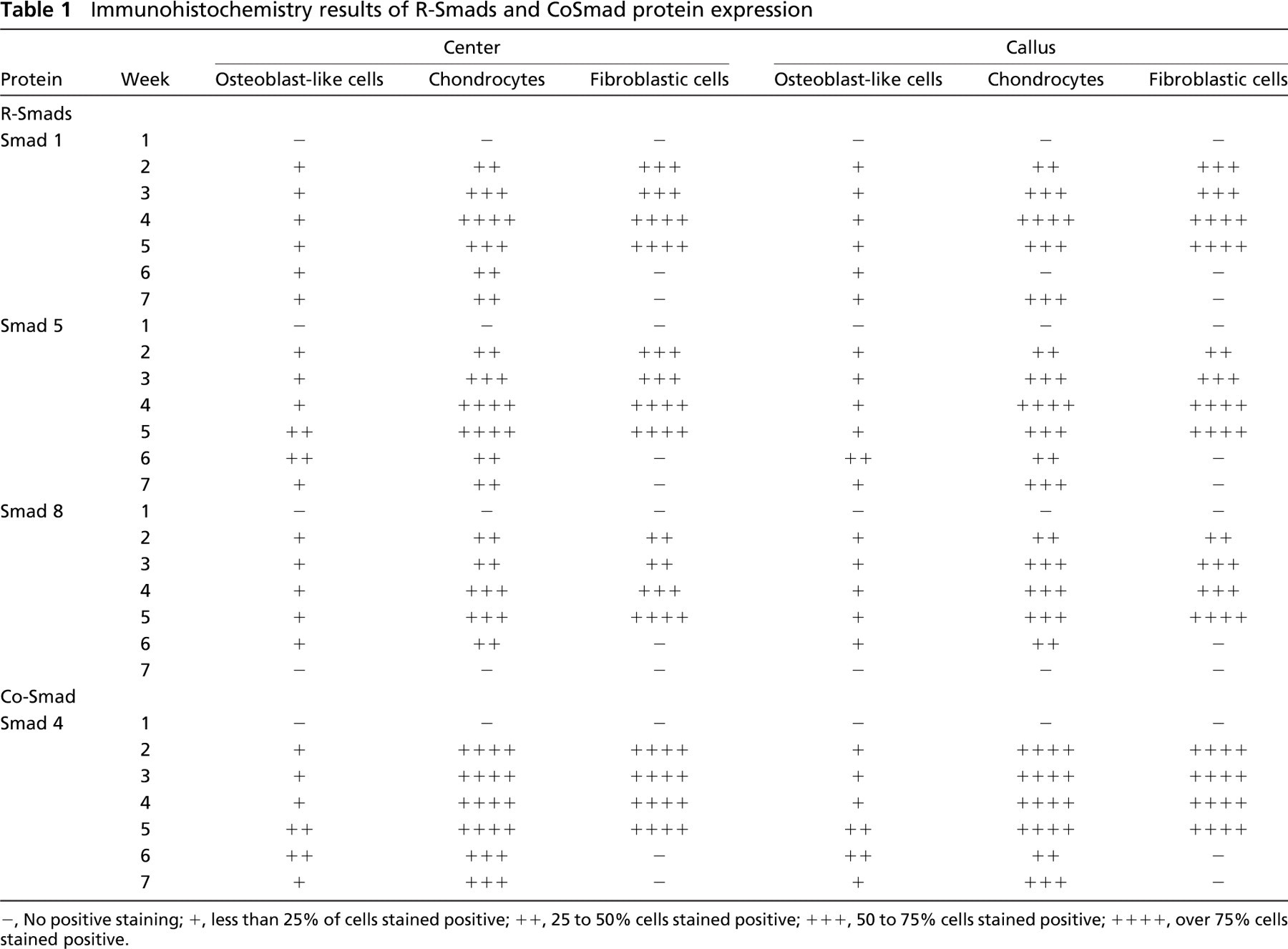
-, No positive staining; +, less than 25% of cells stained positive; ++, 25 to 50% cells stained positive; +++, 50 to 75% cells stained positive; ++++, over 75% cells stained positive.
Immunohistochemistry results of I-Smad and Smad ubiquitin regulatory factor (Smurf) protein expression
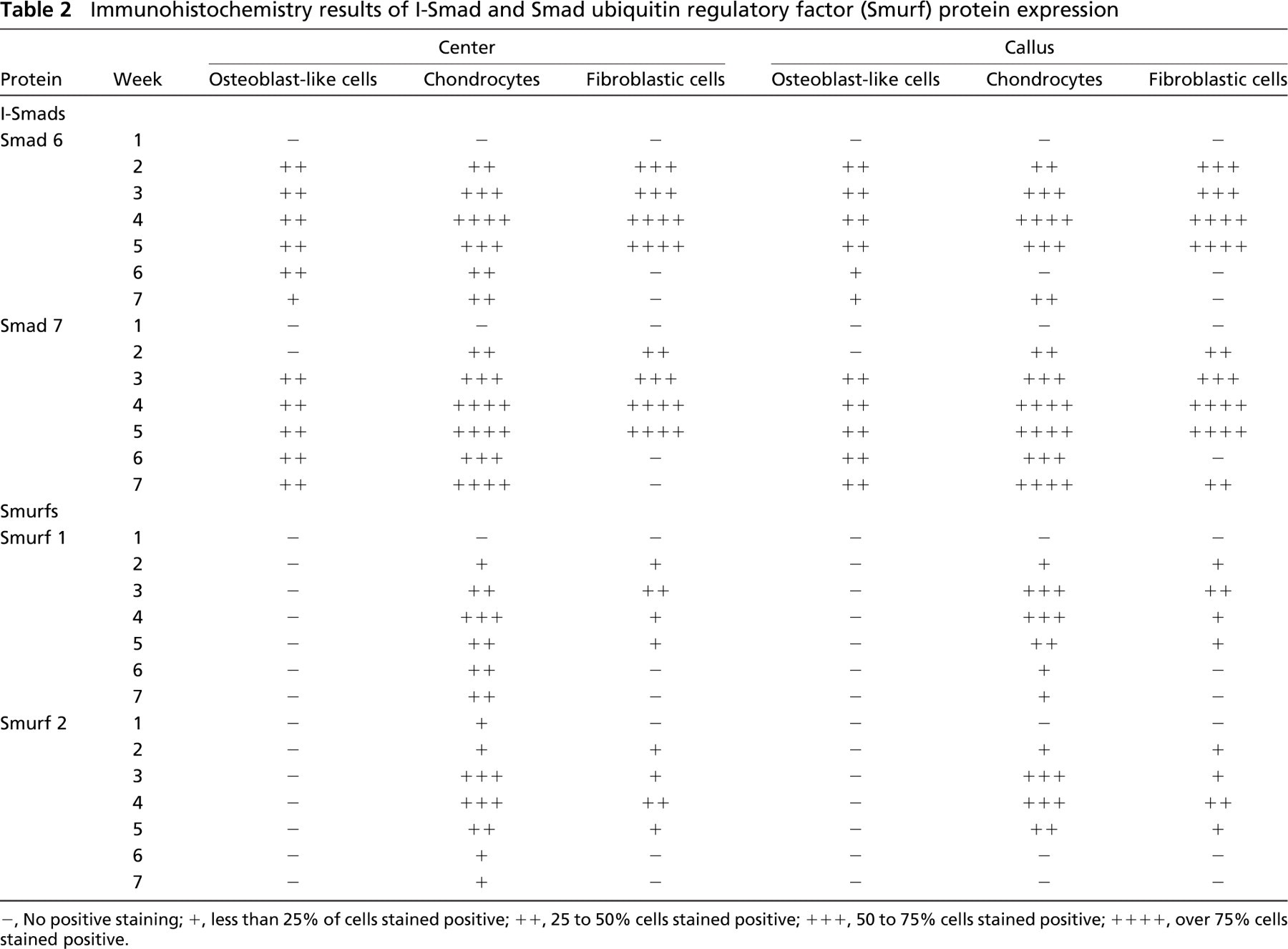
-, No positive staining; +, less than 25% of cells stained positive; ++, 25 to 50% cells stained positive; +++, 50 to 75% cells stained positive; ++++, over 75% cells stained positive.
The three subgroups of Smads (receptor activated, inhibitory, and common-partner Smads) differed in their level and time of expression. Staining for all investigated Smad proteins was negative at 1 week after surgery, before starting distraction. All Smad proteins were strongly expressed by chondrocytes and fibroblasts during distraction and less expressed during the consolidation phase. The receptor-activated Smads (Smads 1, 5, and 8) showed a temporal and spatial pattern of expression similar to the one we previously reported for BMP ligands 2, 4, and 7 and the BMP receptors I and II (Hamdy et al. 2003). Positive immunostaining for R-Smads suggests that the BMP receptors of these cells are activated. Inhibitory Smads, which are antagonists of R-Smad/Co-Smad complex signaling, showed a different pattern of expression. They were present soon after starting the distraction, and their level of expression increased toward the end of distraction. They were also intensely expressed after the distraction was concluded. These findings confirm their inhibitory role in BMPs.
Maximum expression of all Smads and Smurfs was found in chondrocytes and fibroblastic cells compared with osteoblastic cells. This is in accordance with our previously reported results on the expression of BMP ligands and BMP receptors, which were maximally expressed in chondrocytes in the same rabbit model of DO (Hamdy et al. 2003). The presence of Smads in chondrocytes has also been reported in previous studies on BMP signaling in endochondral bone formation (Sakou et al. 1999; Tsumaki and Yoshikawa 2005). In a rat model for fracture healing, however, Smads 1-6 were also localized in osteoblasts, in addition to proliferating liferating chondrocytes and mesenchymal cells (Yu et al. 2002). The difference between these findings and our observed results may be attributed to the presence of mechanical stress applied during distraction, which can lead to a different biological signaling from that of fracture healing—where no distraction forces are applied. Our study did, however, show the expression of inhibitory Smads (Smads 6 and 7) in osteoblasts. This correlates with their function in inhibiting BMP signaling in osteoblasts as suggested in previous studies (Miyazono 2000).
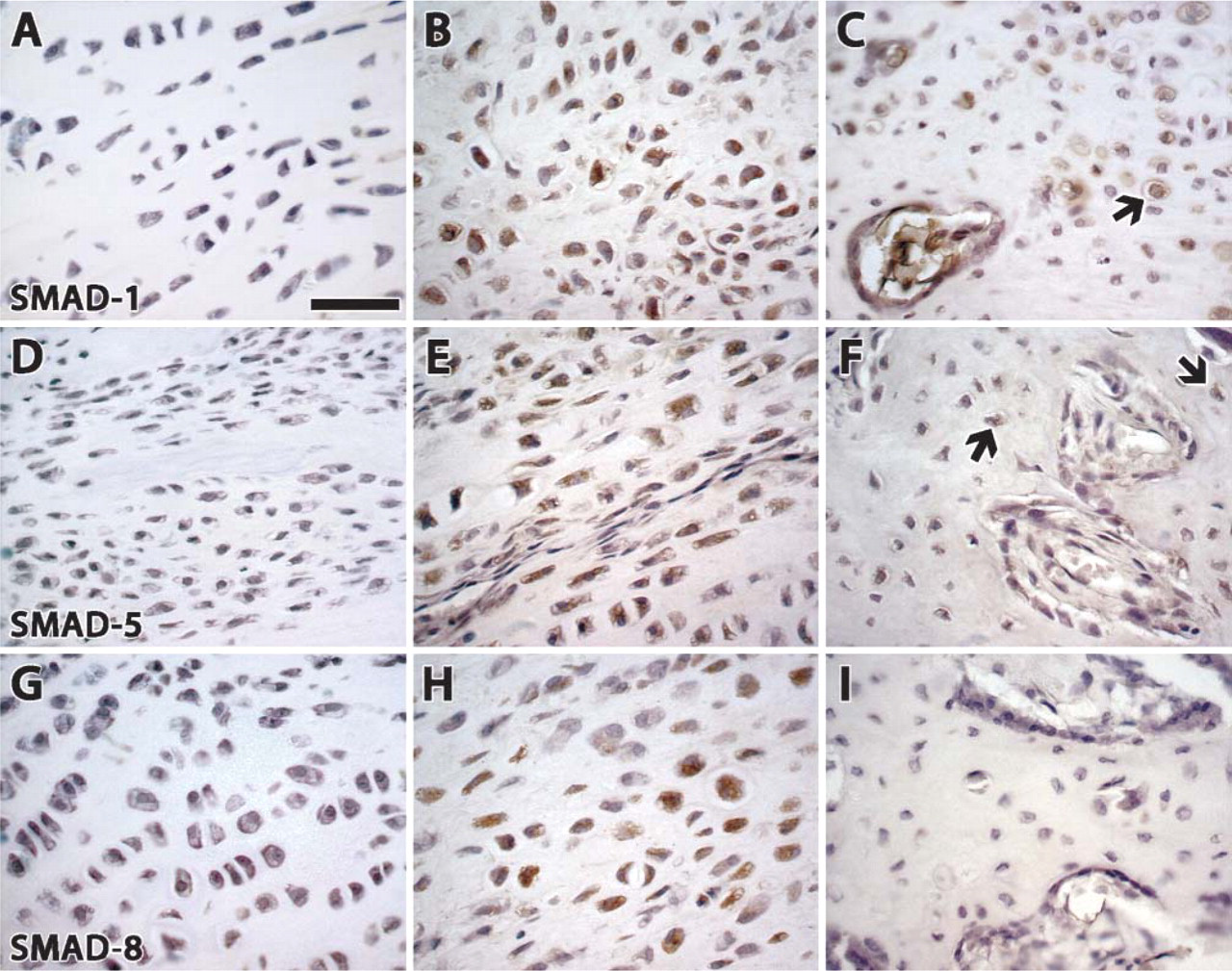
Immunostaining of receptor-activated Smads (1, 5, and 8). (
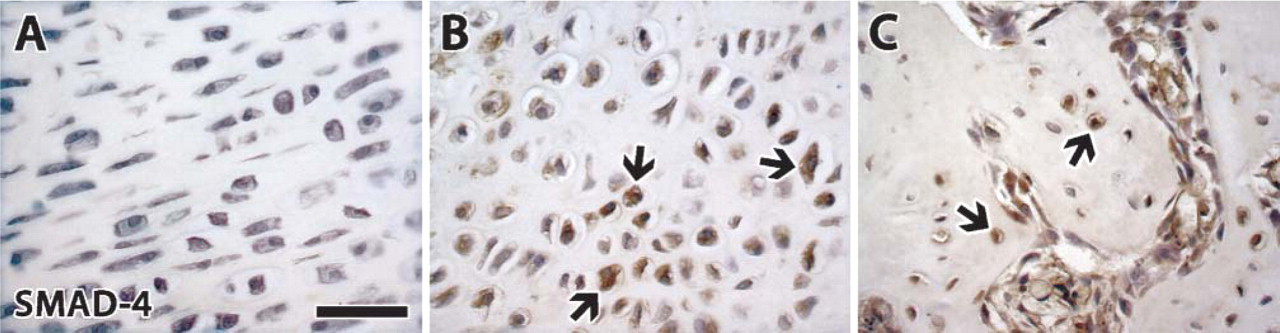
Immunostaining of common-partner Smad 4. (
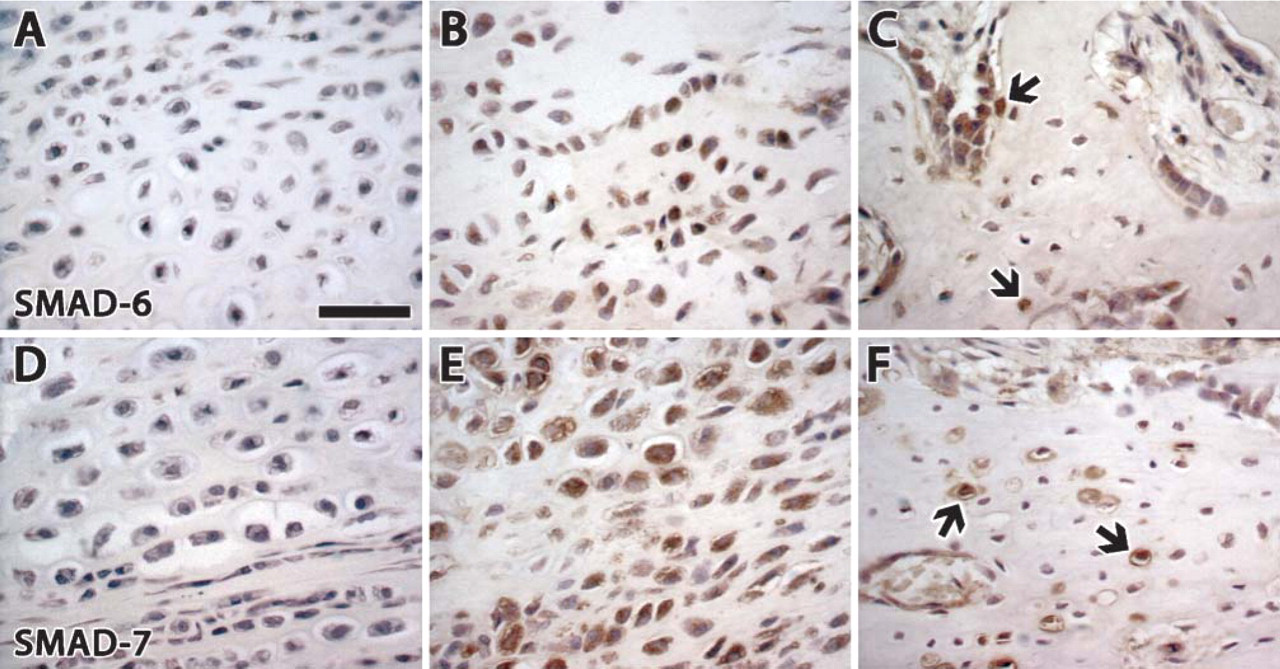
Immunostaining of inhibitory Smads 6 and 7. (
A review of the literature revealed only one published report on Smad signaling in DO. In that study, Farhadieh et al. (2004) found that Smads 1-5 were expressed in osteoblasts, osteocytes, and periosteal lining cells. However, in that study DO was performed on the mandible, which is a flat bone and, therefore, new bone formation occurs predominantly through an intramembranous pathway. Hence, we believe that it is the absence of any chondrocytes in mandibular bone formation that may explain the lack of expression of Smads in chondrocytes. This may be the reason why the Smad expression profile observed in our study of tibial DO varied from mandibular DO as investigated by Farhadieh et al. (2004).
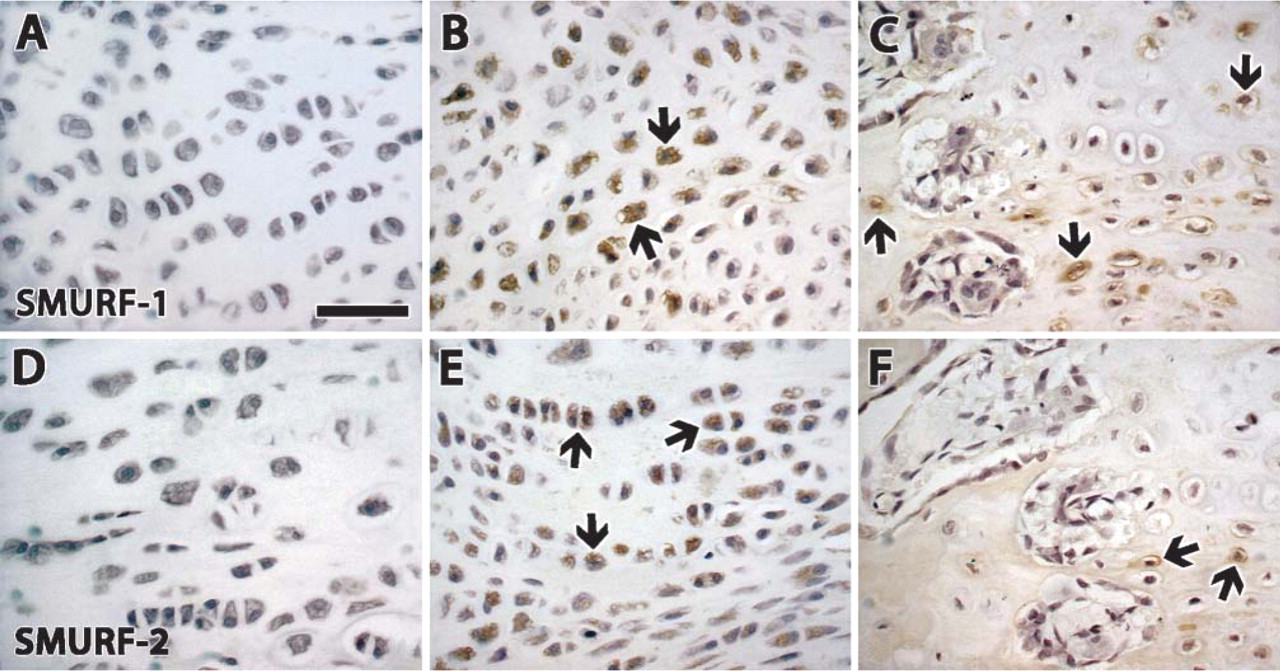
Immunostaining of Smurf 1 and 2. (
We were unable to find any previous studies on the expression of Smurfs during DO or fracture healing. As Smurfs generally operate by binding to Smad proteins (Ying et al. 2003; Tsumaki and Yoshikawa 2005), the minimal staining of Smads in osteoblasts observed in our study could explain the absence of staining of Smurfs in osteoblasts—and conversely the high expression in chondrocytes.
In conclusion, results of this study confirm postreceptor activity for the BMP signaling pathway during DO in the rabbit and suggest that the BMP pathway may play a major role in the process of new bone formation in DO. This is supported by the strong similarity in the temporal pattern of expression and in the spatial colocalization to the same morphological type of cells of BMP ligands, BMP receptors, and their intracellular Smad proteins. Furthermore, our results suggest that the whole BMP signaling pathway is upregulated during the distraction phase of the lengthening process. This is most likely a result of the mechanical forces caused by distraction. These findings may help in the development of novel therapeutic approaches for accelerating bone formation in DO by the application of exogenous growth factors early in the distraction phase of the lengthening procedure or the manipulation of the BMP signaling pathway.
Footnotes
Acknowledgments
This work was supported by Shriners of North America operating grant, Fonds de la Recherche en Santé du Québec, and the National Science and Engineering Research Council of Canada.
We thank Ms. Guylaine Bedard and Mr. Mark Lepik for the illustrations.