
Abstract
Osteopontin (OPN) is an adhesive, matricellular glycoprotein, whose expression is elevated in many types of cancer and has been shown to facilitate tumorigenesis in vivo. To understand the role of OPN in human skin cancer, this study is designed to determine whether OPN is expressed in premalignant [solar/actinic keratosis (AK)] and in malignant skin lesions such as squamous cell carcinomas (SCC) and basal cell carcinomas (BCC), as well as in normal skin exposed or not exposed to sunlight. Immunohistochemical analyses showed that OPN is expressed in SCC (20/20 cases) and in AK (16/16 cases), which are precursors to SCC, but is absent or minimally expressed in solid BCC (17 cases). However, positive staining for OPN was observed in those BCC that manifest differentiation toward epidermal appendages such as keratotic BCC. In sunlight-exposed normal skin, OPN is minimally expressed in the basal cell layer, but in contrast to those not exposed to sunlight, OPN is more prominent in the spinous cell layer with increasing intensity toward the granular cell layer. Additionally, OPN is expressed in the hair follicles, sebaceous glands, and sweat glands of normal skin. In conclusion, these data suggest that OPN is associated with keratinocyte differentiation and that it is expressed in AK and SCC, which have metastatic potential, but minimally expressed in solid BCC.
Keywords
S
OPN is an acidic, secreted protein recently classified as a member of the SIBLING (Small Integrin-Binding, N-Linked Glycoprotein) family (Fisher et al. 2001). OPN from mice and humans are encoded by a single copy gene, Spp1, and are mapped to the Ric locus (Rickettsia tsutsugamushi) of chromosomes 5 and 4 (4q13), respectively (Fet et al. 1989; Patarca et al. 1989; Young et al. 1990). OPN has been cloned from several species and has been shown to consist of highly conserved regions. Starting from the N-terminus, OPN contains a motif with 7–10 consecutive aspartic acid residues (depending on the species), a cell-binding sequence argininylglycylaspartic acid (RGD), a thrombin cleavage site, which is six amino acid residues downstream of the RGD motif, a cryptic epitope (SLAYGLR in mouse and SVVYGLR in human) postulated to be involved in angiogenesis (Hamada et al. 2004), a calcium-binding region, and a hyaluronic acid receptor (CD44)-binding domain (Wai and Kuo 2004; Rangaswami et al. 2006). Depending on the tissue site or factors that regulate its expression, OPN can undergo posttranslational modification (Prince et al. 1987; Sorensen et al. 2003).
The functions of these conserved motifs in OPN have been shown mainly through in vitro and a few in vivo studies to possess calcium binding, chemotaxis, cell adhesion, cell signaling, cell proliferation, and cell survival capabilities. OPN was initially found prominently in bone, kidney tissue, and body fluids. Immunohistochemical studies have since shown its expression in human luminal surfaces of the epithelial cells of several tissues/organs such as the gall bladder; lung; breast; gastrointestinal, urinary, and reproductive tracts; and salivary and sweat glands (Brown et al. 1992; Qu et al. 1994; Ogbureke and Fisher 2004). Moreover, as mentioned above, its high inducibility has resulted in the detection of its elevated expression in several pathological conditions, one of which is cancer (Rittling and Chambers 2004; Rangaswami et al. 2006).
Human cancer of various tissue types such as brain, breast, mouth, salivary glands, thyroid gland, lung, stomach, endometrium, ovary, cervix, kidney, colon, and prostate are reported to express OPN within the cancerous tissues, the stromal cells and infiltrating macrophages (Brown et al. 1994; Bellahcene and Castronovo 1995; Hirota et al. 1995; Saitoh et al. 1995; Chambers et al. 1996; Tunio et al. 1998; Devoll et al. 1999; Shijubo et al. 1999; Thalmann et al. 1999; Agrawal et al. 2002; Kim et al. 2002; Coppola et al. 2004). OPN expression is also elevated in benign tumors (Tiniakos et al. 1998; Devoll et al. 1999; Kusafuka et al. 1999). Additionally, increased serum levels of OPN have also been detected in cancer patients, with some studies suggesting that the level of OPN is inversely correlated to the length of patient survival (Singhal et al. 1997; Fedarko et al. 2001; Hotte et al. 2002; Kim et al. 2002; Koopmann et al. 2004). Collectively, the expression of OPN in both malignant and benign tumors suggests its association in tumorigenesis and metastasis.
Numerous in vitro and in vivo studies by others and our laboratory implicate OPN's role in tumorigenesis, or more specifically, in tumor promotion (Senger et al. 1989; Rittling and Novick 1997; Wu et al. 2000; Chang et al. 2003). More recently, we have shown in vivo that the lack of induced OPN expression in OPN-null mice significantly suppresses benign squamous papilloma development relative to wild-type mice when subjected to the two-stage (initiation-promotion) mouse skin chemical carcinogenesis model (Hsieh et al. 2006), therefore directly supporting the role of OPN in facilitating tumorigenesis.
To better understand the role of OPN in human skin cancer, this study was undertaken to determine whether OPN is expressed in premalignant [solar/actinic keratosis (AK)] and malignant skin lesions such as squamous cell carcinomas (SCC) and basal cell carcinomas (BCC), as well as in normal skin exposed or not exposed to sunlight. Although previous studies have examined normal skin (Baccarani-Contri et al. 1995) and a few cases of skin cancers for OPN expression (Coppola et al. 2004), we offer a more comprehensive study of OPN expression in skin tumors.
Materials and Methods
Chemicals
Monoclonal antibody (MAb 53) to human OPN has previously been characterized and provided by author Ann Chambers (University of Western Ontario) (Bautista et al. 1994; Allan et al. 2006). The isotype control antibody to MAb 53 is IgG 2a, κ, and this was purchased from BD Biosciences Pharmingen (San Diego, CA).
Specimens
Institutional Review Board approval was obtained for the use of archival paraffin-embedded human specimens from the Veterans Hospital. Paraffin-embedded specimens were sectioned at 5μm and stained with hematoxylin and eosin (H&E). The specimens diagnosed with AK, SCC, and BCC were derived from male subjects with an age range of 36–90 years (Table 1). Kidney tissues were used as a positive control for OPN expression. Sunlight-exposed normal skin was obtained from thin skin region or those areas surrounding the AK, SCC, and BCC. The foreskin samples, which are considered unexposed to sunlight, were obtained by signed consent of healthy adults and/or guardians for children younger than 18 years of age from Thailand.
Immunohistochemical Analyses
In addition to routine H&E stain, all skin specimens were analyzed by immunohistochemistry using the alkaline phosphatase detection method. As a positive control and to test the specificity of the OPN antibody, normal kidney tissues were used. In the latter analyses, the horseradish peroxidase detection method was used instead of the alkaline phosphatase method to detect OPN in kidney tissues, because it contains high levels of endogenous alkaline phosphatase (Weiss et al. 1986).
Age range, mean age, and sex of patients with skin lesions
Three consecutive sections of 5 μm from each specimen were placed on superfrosted slides (Fisher Scientific; Pittsburgh, PA). Sections were deparaffinized with xylene for 20 min at 48C and transferred to 100% ethanol for 10 min, and the latter process was repeated one more time. Slides were rehydrated by rinsing stepwise with decreasing concentration of ethanol (95%, 75%, and 50%), followed by a water rinse, and transferred to a Tris-buffered solution. The skin sections were treated with pronase, antigen retrieved by steaming for 1 hr in Glyca solution (BioGenex; San Ramon, CA), and blocked with Cyto-Q-biotin-free solution (Innovex Biosciences; Richmond, CA) for 25 min. Slides from each specimen were incubated with a 1:150 dilution of monoclonal antibody to human OPN (MAb 53) or the same concentration as MAb 53 of isotype control antibody (IgG2a, κ) overnight at 4C, and this incubation was followed by the addition of secondary antibody conjugated to biotin for 30 min. To determine whether, nonspecific binding occurs from secondary antibody conjugated to biotin (BioGenex), an additional consecutive slide from each specimen was incubated with only the secondary antibody conjugate to biotin. All these slides were incubated with streptavidin-alkaline phosphatase (BioGenex) for 30 min and rinsed thoroughly before developing them with the addition of Fast Red chromogen (BioGenex). Positive staining appears pink. Slides were counterstained with hematoxylin and mounted with Supermount (BioGenex).
Kidney sections used as a positive control were treated with pronase and antigen retrieved as described above, followed by incubation with 3% hydrogen peroxide and blocked with Cyto-Q-biotin free blocking solution. After incubation with the primary antibody and secondary antibody conjugated to biotin, the specimens were incubated with streptavidin conjugated to horseradish peroxidase and developed using 3, 3′-diaminobenzidine substrate (BioGenex). Positive staining appears brown.
RT-PCR
Paraffin-embedded samples were melted and cut into smaller tissues before treating them with lysis solution as previously described (Chomczynski and Sacchi 1987). The total RNA that was extracted was transcribed to cDNA by RT using oligo (dT). Primers specific for human spp1 were 5′-TGG CCG AGG TGA TAG TGT G-3′ (forward) and 3′-CGG GGA TGG CCT TGT ATG-3′ (reverse), which generated a 400-bp pair PCR product (Kadkol et al. 2006). To normalize expression of OPN, the forward primers 5′-CCG TGC CTC CAA GAT GAC AAA-3′ and the reverse primer 5′-GTT CGG TCC TTG CGG GCT TTC AC-3′ were used to amplify a 300-bp segment of the housekeeping gene, human S26 (Pan et al. 2003). Eppendorf Mastercycler gradient was used to amplify (42 cycles) the DNA fragments, with each cycle consisting of 90C for 20 sec and 59C for 35 sec.
Results
Immunohistochemical analyses of all normal and skin lesions were stained with MAb 53 to human OPN. To show the specificity of this antibody to human OPN, normal kidney specimens were used as a positive control, because it has been previously reported to consistently express OPN in specific regions (Verhulst et al. 2002; Ogbureke and Fisher 2005). In agreement with previous observations, normal kidney tissue expresses OPN in the distal convoluted tubules, and likewise, no expression was observed in the glomerulus (Figures 1A and 1B).
To determine whether OPN is expressed in cutaneous SCC, immunohistochemical studies were performed on 20 SCC cases. Three consecutive slides of each case were incubated with monoclonal antibody to human OPN, its isotype control antibody, IgG 2a,κ, or secondary antibody conjugated to biotin. All 20 cases of SCC were shown to express elevated levels of osteopontin (Figures 1C, 1D, 1F, and 1G). In most SCC samples, OPN staining was localized in the cytoplasm (Figure 1G), and in some cases, in the extracellular matrix immediately surrounding the cancer cells (Figure 1D). The other stromal regions of SCC were basically negative for OPN. All SCC cases stained with isotype control antibody (Figures 1E and 1H) or with secondary antibody conjugated to biotin (data not shown) were negative.
The premalignant skin lesions AK, known as precursors to the development of SCC, also expressed OPN in all 16 cases (Figures 1I and 1K). The basal cell layer in the AK showed mild staining for OPN with increasing intensity of cytoplasmic OPN expression as the cells become more differentiated, such as in the upper spinous cell layer and the granular cell layer. The faint positive staining observed in the large keratin region (Figures 1I and 1J, arrowhead) of AK is nonspecific staining that came from secondary antibody conjugated to biotin, because consecutive slides incubated with isotype control antibody (Figures 1J and 1L) and secondary antibody conjugated to biotin (data not shown), respectively, showed patches of positive staining in these regions.
In contrast to SCC and AK, solid or undifferentiated basal cell epitheliomas showed minimal to no expression of OPN (Figures 2A and 2C). However, for those BCC that differentiated toward the epidermal appendages, such as differentiation toward hair (keratotic BCC), positive staining for OPN was observed (Figures 2E and 2G).
Besides its expression in the skin lesions, OPN was also detected in non-tumor or what appeared to be “normal” epidermal regions (Figure 2A) and sun-exposed normal skin (Figure 3A). More specifically, basal cells showed minimal OPN expression with positive staining in the spinous cell layer and in the granular cell layer. OPN expression was also detected in the sweat glands, which seem to be localized in the myoepithelial cells (Figure 3C). Additionally, hair follicles and sebaceous glands are immunoreactive for OPN (Figures 3E and 3F). The presence of OPN in sweat glands and hair follicles has also been previously reported in humans and rats, respectively (Brown et al. 1992; Yu et al. 2001; Rivet et al. 2006; Ogbureke and Fisher 2007).
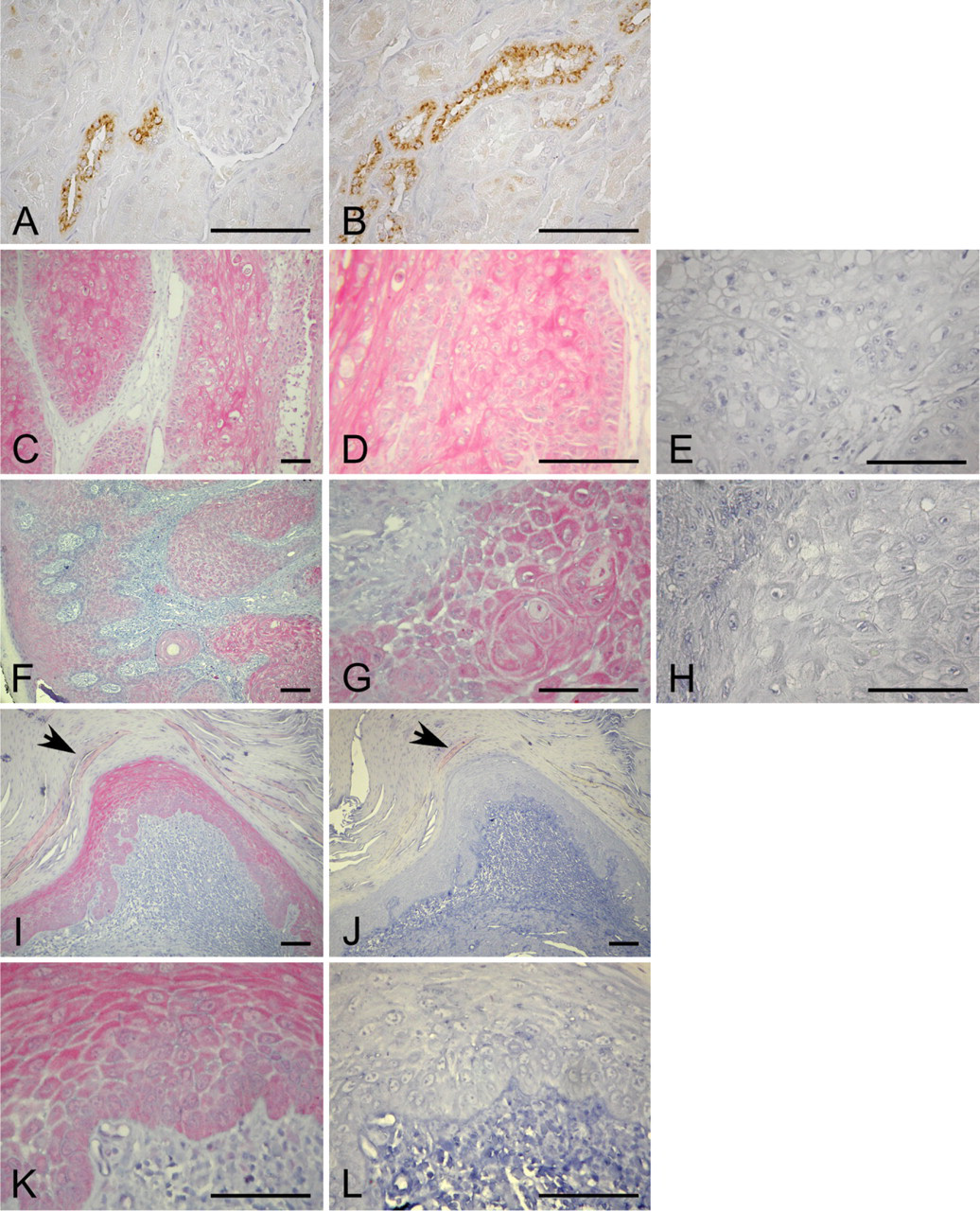
Immunohistochemical analyses of kidney, cutaneous squamous cell carcinomas (SCC), and solar keratosis (AK) for osteopontin (OPN). (
Because OPN expression is observed in the epidermis of skin that has been exposed to UVB irradiation, we questioned whether OPN is also expressed in skin that has not been chronically exposed to sunlight. We assume that the foreskin should be the least exposed to sunlight and used this tissue to analyze for OPN expression. Again, minimal to no OPN expression was detected in basal cell layer of the foreskin, whereas the upper layers of the epidermis were positive for OPN (Figure 3G). However, of 11 samples of foreskin from various ages, we found that the intensity of OPN expression in the spinous and granular cell layers was much lower than in those skin samples that were frequently exposed to sunlight (compare Figure 3A to 3G). Semiquantitative RT-PCR showed variable OPN expression in the epidermis of the foreskin from the age range of 19- to 56-year-old subjects (Figure 4).
Discussion
Although elevated levels of OPN have been observed in several types of tumors, its expression in the premalignant and malignant non-melanoma epidermal skin lesions has not been reported in detail (Coppola et al. 2004). To the best of our knowledge, this study is the first to comprehensively show the localization of OPN expression in AK and cutaneous SCC and BCC. Previous studies looked at four cases of melanomas and two cases of BCC. None of the melanomas expressed OPN, but BCC were found to be positive for OPN (Coppola et al. 2004). Contrary to their results for BCC, we observed that OPN was minimally or not expressed in all the solid BCC. However, those BCC that histologically manifested differentiation toward epidermal appendages such as hair follicles were positive for OPN. Whether the two BCC cases reported in the previous study were of the differentiated form was not mentioned (Coppola et al. 2004).
The differential expression of OPN between SCC and solid BCC is intriguing. The fact that OPN is consistently expressed in all cases of cutaneous SCC, which are capable of metastasis, and not expressed in the solid BCC, which have no potential of metastasis, suggest the potential role of OPN in facilitating metastasis. Recent studies support this possibility. Injection of breast cancer cells overexpressing OPN into the fat pad of nude mice resulted in increased metastasis to the lymph node and lung (Allan et al. 2006).
In addition to OPN expression in SCC, it has also been observed in AK in the basal cell layer and more intensely toward the upper differentiated keratinocytes. A previous report indicated that 10% of AK can transform to malignant SCC (Marks et al. 1988). Whether OPN plays a causal role in the progression of AK to SCC remains to be addressed.
The observation that OPN is not expressed in the solid BCC is consistent with the staining pattern of sunlight-exposed and -unexposed skin (foreskin), where the basal cell layer shows minimal OPN expression. However, OPN is present in the form of BCC with differentiation toward epidermal appendages, implicating its association with keratinocyte differentiation. OPN's role in cell differentiation of HL-60 cells was initially suggested by Somerman et al. (1995) but was later refuted by the same group (Atkins et al. 1997). Recent studies support the requirement of OPN expression in maintaining macrophage function and its differentiated state (Nystrom et al. 2007). Additionally, OPN expression usually correlates with the intermediate and later stages of osteoblasts differentiation from mesenchymal stem cells (Ibaraki et al. 1992; Lecanda et al. 1997). OPN expression in esophageal squamous carcinoma also suggests its association with differentiation (Liu et al. 2007). These findings, taken together with our observations, implicate the potential role of OPN in differentiation. Generation of mice which lacks or is deficient in OPN expression (Liaw et al. 1998; Rittling et al. 1998), however, did not support OPN's role in differentiation, because these mice show normal embryogenesis, which requires a series of cell proliferation and differentiation process. There is also the possibility that an important process such as differentiation might have redundant matricellular proteins in place, and therefore, the lack of only OPN does not manifest abnormal embryonic development.
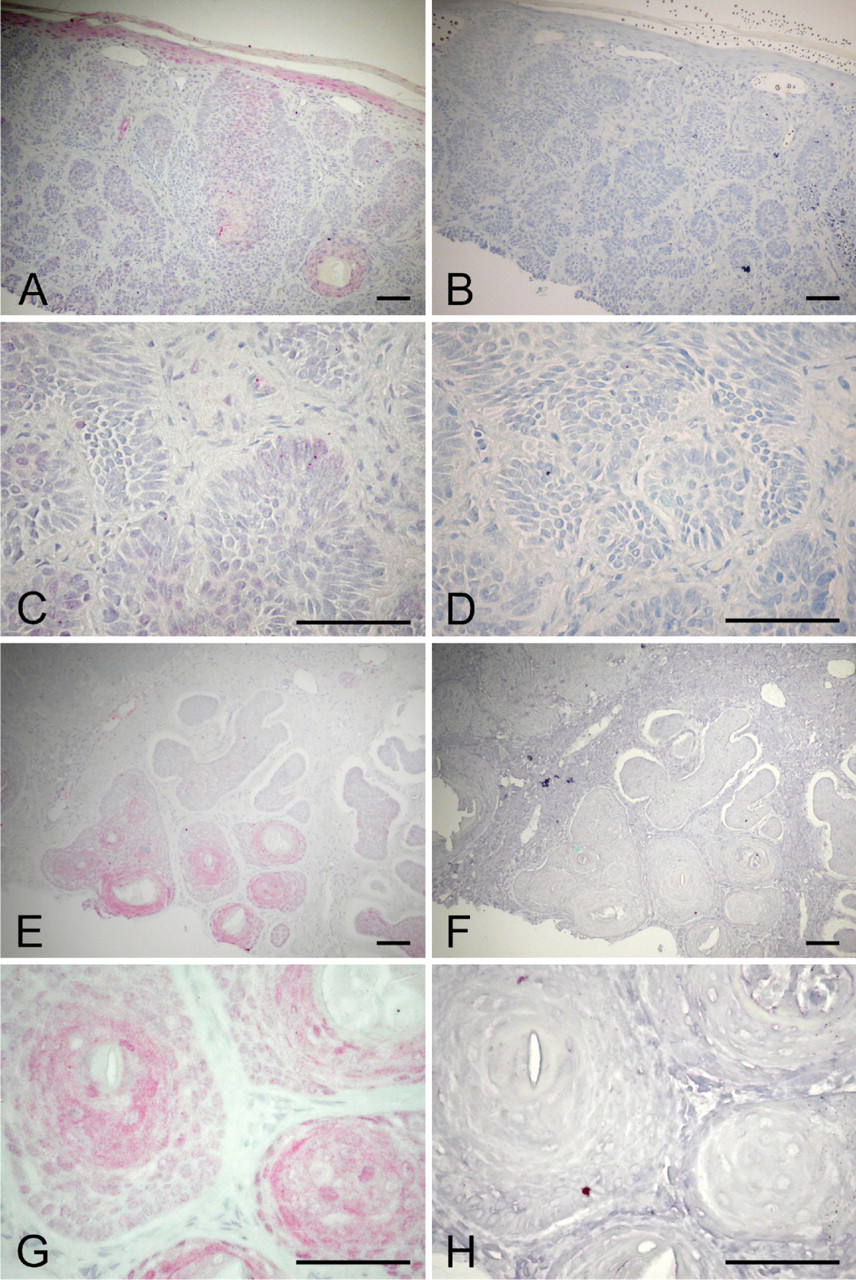
Immunohistochemical analyses of basal cell carcinomas (BCC) for OPN. Two cases of BCC: (
Whether OPN plays a critical role in directing human keratinocyte differentiation remains to be examined. In addition to its possible role in differentiation, it is also likely that the induction of OPN expression before or during the differentiation process is important for enhancing cell survival, especially during the time of external or internal stress, because numerous in vitro and in vivo studies support its role in preventing stress-induced cell apoptosis. Using the two-stage (initiation by 7, 12-dimethylbenz(a)anthracene (DMBA) and tumor promotion by 12-0-tetradecanoylphorbol-13-acetate (TPA) mouse skin chemical carcinogenesis model in addressing OPN's role in tumor promotion, we have shown that the ablation of OPN resulted in significant decrease in papilloma development (Hsieh et al. 2006). DMBA-induced apoptosis assays showed significant apoptosis of the basal cells in the skin of OPN null mice compared with that of wild-type mice, suggesting that DMBA- and TPA-induced OPN secretion into the microenvironment of initiated cells could enhance their survival, and consequently, more papillomas are observed in the wild-type mice (Hsieh et al. 2006).
With respect to skin cancer development, chronic exposure of UVB (280–320 nm) radiation is the major risk factor. However, its mechanisms are still not fully understood. Unlike the two-stage mouse skin chemical carcinogenesis model where the dorsal skin is initiated with the carcinogen only once, followed by continuous application of tumor promoter, UVB acts as both an initiator and a tumor promoter simultaneously.
As an initiator or a carcinogen, UVB can generate mutations in RAS oncogene and tumor suppressor genes, p53 and PTCH, found in skin cancer (Pierceall et al. 1991; Brash et al. 1996; Gailani et al. 1996). As a tumor promoter, UVB is postulated to stimulate clonal expansion of initiated cells through several mechanisms and subsequently the development of premalignant and/or malignant tumors. Some of the possible mechanisms of UVB-induced tumor promotion are (1) the generation of reactive oxygen species, which could stimulate RAS and mitogen-activated protein kinase, leading to activation of activating protein-1 (AP-1) and nuclear factor-κB, both known to contribute to cell proliferation and tumorigenesis, and (2) the activation of epidermal growth factor receptor shown to be important in keratinocyte proliferation and mediate UV-induced epidermal hyperplasia (El-Abaseri et al. 2005).
In addition to UVB inducing tumor promotion by promoting clonal expansion, we postulate that UVB may also contribute to enhancing the survival of initiated cells through their interaction with altered matrix microenvironment such as the presence of induced secreted OPN (Hsieh et al. 2006). Whether OPN expression is induced by sunlight will need to be determined. We observed that sunlight-exposed skin has higher expression of OPN than foreskins normally not exposed to UVB. There are several lines of evidence suggesting that UVB could indirectly stimulated OPN expression. Activated RAS has been found in both actinic keratoses and cutaneous SCC (Spencer et al. 1995). The human OPN promoter has been shown to consist of both a functional RAS-activated enhancer (Denhardt et al. 2003) and an AP-1 response element (Nakamachi et al. 2007). Both the initiator and tumor promotion effect of UVB can induce the activation of RAS and AP-1, which can result in triggering OPN expression and secretion into the microenvironment of the initiated and normal keratinocytes. OPN, on binding to appropriated cell surface receptors, could in turn enhance the survival of UVB-induced initiated keratinocytes and consequently facilitate the development of AK and SCC.
UVB may also indirectly stimulate OPN expression through mutated p53, which is commonly found in >90% of cutaneous SCC and 50% of AK (Brash et al. 1996). Generally, elevated expression of OPN has been shown to be associated with p53 mutation (Pan et al. 2003; Graessmann et al. 2006) and p53-null mesenchymal stem cells (Tataria et al. 2006). However, OPN has also been reported to be transcriptionally regulated by TP53 in embryonic fibroblasts (Morimoto et al. 2002); thus, additional studies as to whether OPN is suppressed or stimulated by p53 in epithelial cells will need to be clarified.
Another indirect route of stimulating OPN is through UVB-induced production of vitamin D3, which can be converted to the active form 1α,25-dihydroxyvitamin D3 (calcitriol) in the skin We have previously shown that calcitriol induces the synthesis and secretion of OPN in murine epidermal-like cells (Chang and Prince 1991; Chang et al. 1994). The vitamin D response element in the rat and mouse OPN promoter has been reported, and it is likely that the human OPN promoter contains the vitamin D response element, because human primary osteoblasts can be induced by calcitriol to produce OPN mRNA (Atkins et al. 2007). Collectively, these findings support the possibility of UVB-induced OPN expression, possibly indirectly through calcitriol in normal skin and/ or through both calcitriol and activated Ras, AP-1, and mutated p53 in AK and SCC.
In conclusion, our data suggest that OPN is associated with skin differentiation and/or is induced by sunlight, because foreskin not exposed to sunlight expressed less OPN. Additionally, OPN is associated with preneoplastic skin lesions, AK, and SCC, which have high potential to metastasize, but not with solid BCC.
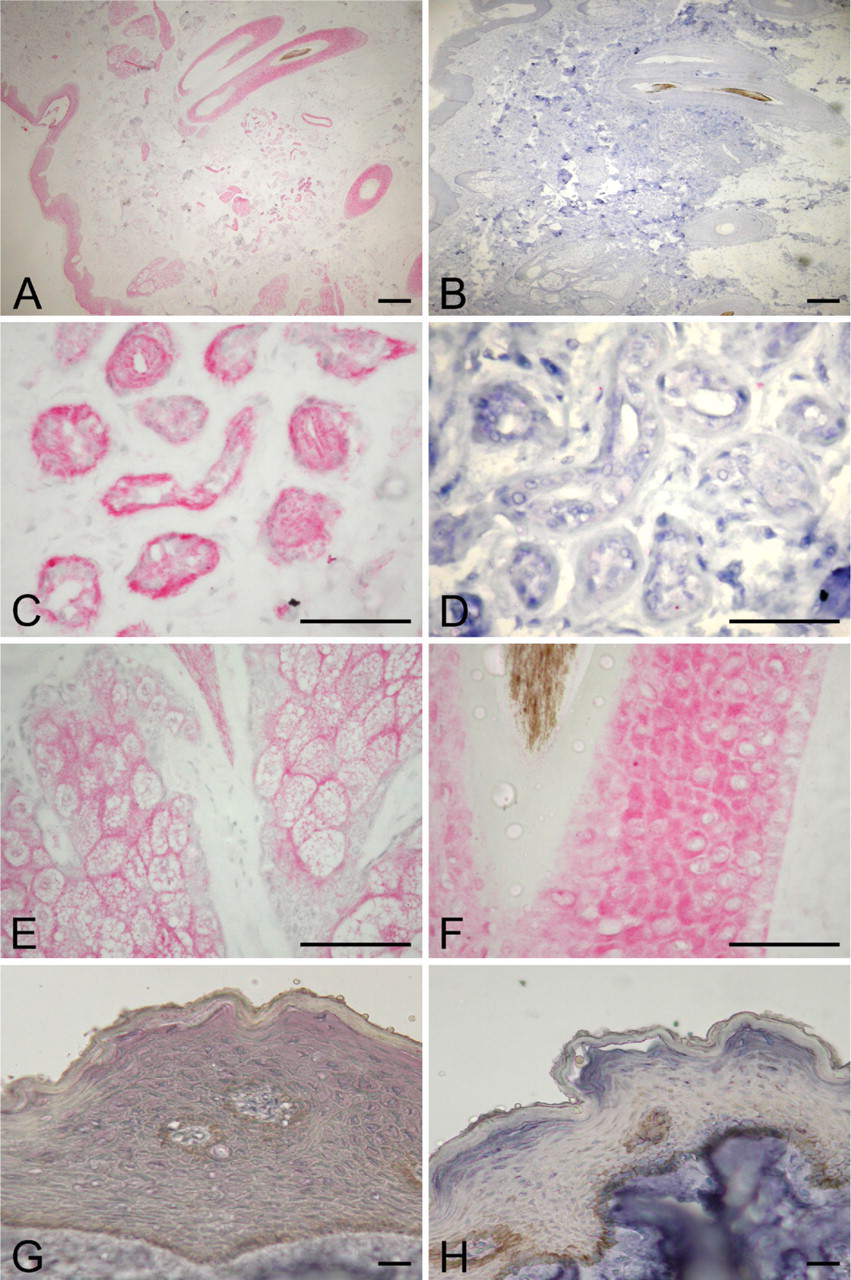
Immunohistochemical analyses of skin exposed and unexposed to sunlight for OPN. (
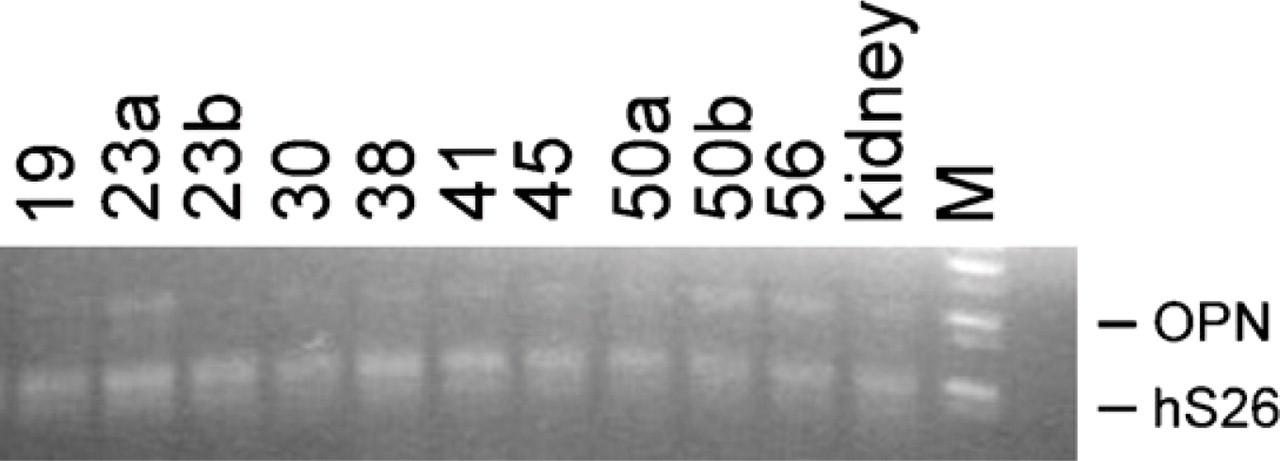
OPN mRNA expression in foreskin. Semi-quantitative analyses of OPN mRNA by RT-PCR. The foreskin samples are labeled by age. a and b designate specimens from different individuals. Kidney sample is used as a positive control. hS26, human ribosomal protein. M, DNA standard.
Footnotes
Acknowledgements
This work was supported by the Skin Cancer Foundation and the U.S. National Institutes of Health Grant R01 CA90920 (to P-LC), P30-AR and VA grants (to CAE), the UAB Comprehensive Cancer Center, and Grant 04MAY00089 from the Ontario Cancer Research Network (to AFC). A.F.C. is a Canada Research Chair in Oncology, supported by the Canada Research Chairs Program.