
Abstract
Mammalian spermatogenesis is a complex process involving regulatory interactions of many gene products. In this study, we found that dynein light chain-1 (DLC1), a component of the dynein motor complex, is highly expressed in mouse and rat testes. Immunohistochemically detectable levels of DLC1 are observed specifically in spermatids in steps 9–16 in distinct subcellular compartments: in steps 9–11, DLC1 is predominantly localized in the nucleus; in steps 12 and 13, it is found in both nucleus and cytoplasm; and in step 14–16, it is present exclusively in the cytoplasm. In addition, we found p21-activated kinase 1 (Pak1), a protein kinase that activates DLC1 by phosphorylating DLC1 at Serine 88, was also expressed during these stages of spermatogenesis. Pak1 was also expressed in Leydig cells, in preleptotene primary spermatocytes, and in round spermatids. The spermiogenic stage-specific expression of DLC1 suggests a role for DLC1 in chromatin condensation, spermatid shaping, and the final release of sperm from the spermatogenic epithelium. Further, Pak1 may also play a role in spermiogenesis by regulating DLC1 phosphorylation and, consequently, its function.
Keywords
Cytoplasmic dynein is a large, multisubunit molecular motor complex that translocates cargoes toward the minus ends of microtubules. The dynein complex was originally shown to regulate the movement of chromosomes, the assembly and orientation of mitotic spindles, and nuclear migration (Steuer et al. 1990; Vaisberg et al. 1993; Holzbaur and Vallee 1994; Beckwith et al. 1998; King 2000). Dynein light chain 1 (DLC1), a subunit of the dynein motor complex, is a highly conserved small protein composed of 89 amino acids. In addition to playing an essential role in the dynein motor complex, DLC1 also regulates other biological events by binding to various proteins and enzymes. For example, DLC1 binds to and inhibits neuronal nitric oxide synthase and thus is also known as a protein inhibitor of neuronal nitric oxide synthase (Jaffrey and Snyder 1996). DLC1 is also known to bind to other proteins, such as the proapoptotic Bcl-2 family member protein, Bim (Puthalakath et al. 1999); the Drosophila RNA localization protein, Swallow (Schnorrer et al. 2000); myosin V (Puthalakath et al. 2001); neuronal scaffolding protein, GKAP; and IκBα, an inhibitor of the NFκB transcription factor (Naisbitt et al. 2000). Despite the widespread expression of DLC1, its specific functions in different tissues and organs remains unknown, with the exception of its role in Drosophila. In Drosophila, partial loss of function of DLC1 leads to morphogenetic defects, female sterility, alterations in axonal guidance, and total loss of function, resulting in apoptosis and embryonic death (Dick et al. 1996; Phillis et al. 1996). In addition, we have recently shown that p21-activated kinase 1 (Pak1), a serine/threonine kinase, interacts with and phosphorylates DLC1 at Serine 88, and that Pak1-DLC1 interactions play a fundamental role in cell survival (Vadlamudi et al. 2004).
Protein kinases, the enzymes responsible for the phosphorylation of a wide variety of proteins, are the largest class of signaling molecules known to regulate growth, development, and neoplastic transformation (Kumar and Wang 2002). Pak1 was originally discovered as a cytoskeleton-associated protein kinase. Pak1 is involved in the maintenance of cell shape, cell migration, and cell survival (Kumar and Vadlamudi 2002). In addition, Pak1 has also been localized in the cell nucleus, where it appears to be involved in the phosphorylation of histones and transcription factors, such as estrogen receptor and Stat5a (Li et al. 2002; Wang et al. 2002, 2003). In the present report, we document that DLC1 and Pak1 are highly expressed in mouse and rat testes in a developmentally regulated, stage-specific manner, and that Pak1 regulation of the testis-specific isoform of cAMP-responsive element modulator (CREM®) might play an important role in the regulation of DLC1 expression in testes.
Materials and Methods
Animals and Tissue Preparation
Mice and rats used in the experiments were housed in the Institutional Animal Care Facility according to the National Institutes of Health and institutional guidelines for laboratory animals. Male mice of outbred genetic backgrounds (Black6xDBA) of 1, 5, 10, 15, 25, 30, and 90 days of age, and adult Lewis-Brown rats of 60 days of age were used for testis collection. For histological studies, testes were dissected and fixed in Bouin's solution for 2–12 hr depending on the size. After dehydration, the testes were embedded in Paraplast (Fisher Scientific Co.; Pittsburgh, PA) and further processed into 4-μm-thick sections. For Western blotting, the testes were directly homogenized in NP-40 lysis buffer. RNA was extracted with Trizol (Invitrogen; Carlsbad, CA) according to manufacturer's protocol.
RT-PCR and Northern and Western Blotting
For RT-PCR, we used the Access RT-PCR system (Promega Corp.; Madison, WI) and followed the protocols suggested by the manufacturer. The primers used were F-CTGTCTTCTGCTGCTTGAGCG and R-GTACACAACACAACAGATTCAAACGAG. For Northern blot analysis, 20 μg of total RNA was resolved on a 1% formaldehyde agarose gel, transferred to a nylon membrane, probed with appropriate probes, and exposed to a phosphoscreen. For immunoblot analysis, 40 μg of total protein lysate was resolved on a 15% SDS-polyacrylamide gel, transferred to a nitrocellulose membrane, and probed with appropriate antibodies.
Immunohistochemistry and Immunofluorescence Studies
Immunohistochemical staining was done with the indirect enzyme labeling method, as described previously (Wang and Zhao 1999). Briefly, the sections were dewaxed in xylene and rehydrated by exposure to descending concentrations of ethanol, immersed in 0.3% H2O2/methanol for 30 min, and washed with PBS. The sections were then sequentially incubated with 1% normal rabbit serum for 1 hr; mouse anti-DLC1 (1:200; Transduction Lab, Cat # 610,726), rabbit anti-CREM® (1:250, a gift from Dr. Paolo Sassone-Corsi, France), or rabbit anti-Pak1 (1:50; Santa Cruz Biotechnology, Santa Cruz, CA) for 2 hr; then with horseradish peroxidase-conjugated donkey anti-mouse or anti-rabbit IgG (1:100; Amersham Biosciences, Piscataway, NJ) for 1 hr, with PBS washes after each incubation. Finally, the antigen sites were visualized with diaminobenzidine/H2O2 solution, and the sections were counterstained with Mayer's hematoxylin for 30 sec. For specificity control, the primary antibody was replaced with normal mouse or normal rabbit serum. In the case of immunofluorescence studies, the second antibody was replaced with FITC-labeled anti-mouse IgG (1:200; Molecular Probes).
Plasmids
Pak1 expression plasmids have been described previously (Adam et al. 1998). pSV-CREM was provided by Dr. Sassone-Corsi. DLC1 promoter luciferase constructs were made by amplifying the upstream fragments of DLC1 from the BAC clone and put into the pGL3-basic vector at the site of KpnI/XhoI.
Cell Culture, Transfection, and Luciferase Assay
COS-7 cells were maintained in DMEM/F12 medium supplemented with 5% fetal calf serum. Cells were plated in six-well culture plates, and transfection was performed using the Fugene (Roche Diagnostics; Basel, Switzerland) method. Luciferase assay was performed according to the manufacturer's instructions, as described previously (Mazumdar et al. 2001), and the results were standardized against the β-gal luciferase activity shown by the internal control. Each experimental group included three triplicate plates.
Results
DLC1 Highly Expressed in Mouse Testes
Because the testis is a highly specialized organ in which cells are constantly dividing and differentiating, which involves chromatin condensation and the reshaping of germ cells, we examined the expression of DLC1, a critical component of the dynein motor complex, using Northern and Western blot analysis of multiorgan blots. This showed DLC1 was highly expressed in the testis (Figures 1A and 1B).
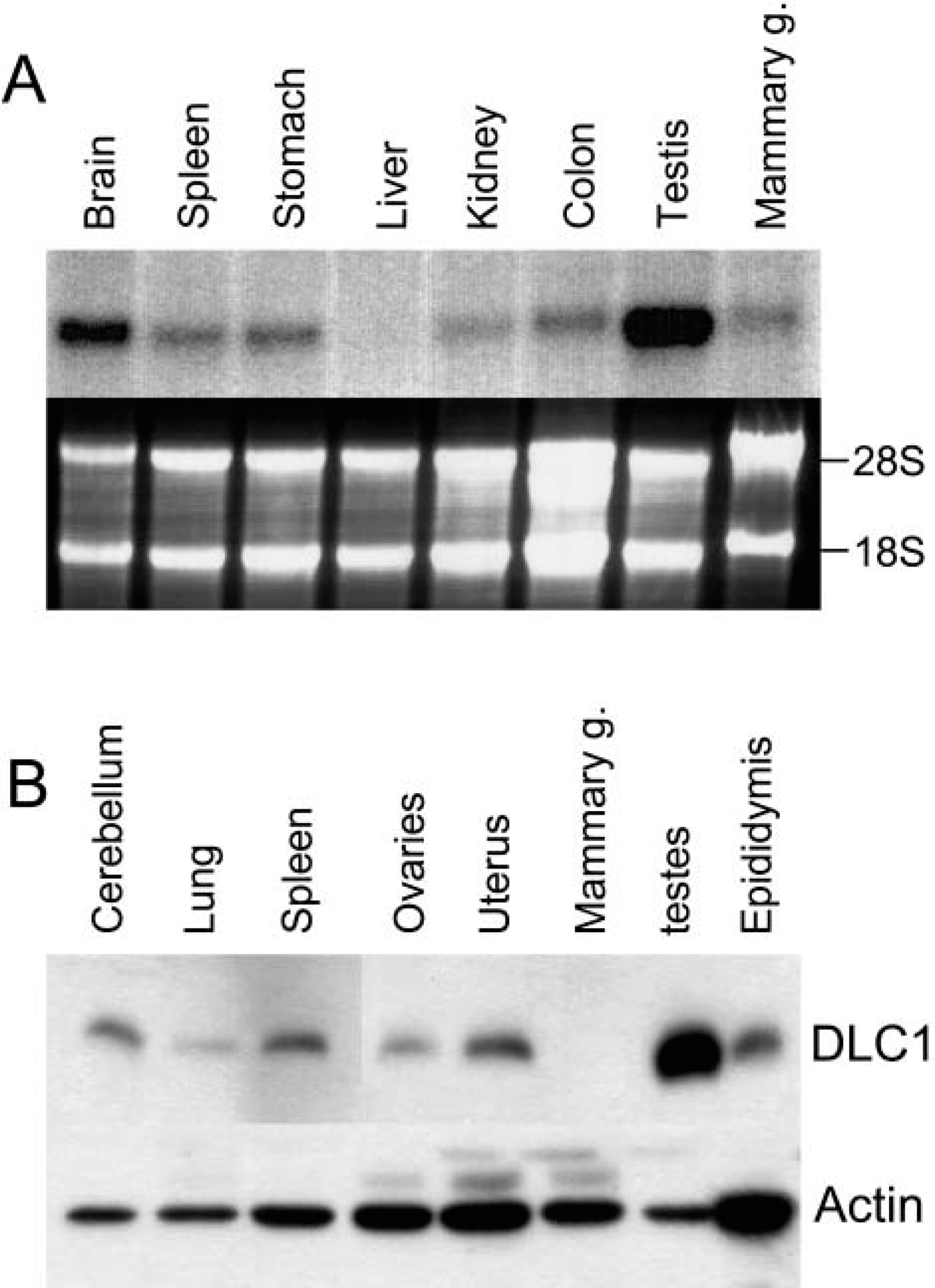
Expression of dynein light chain-1 (DLC1) in a variety of mouse organs.
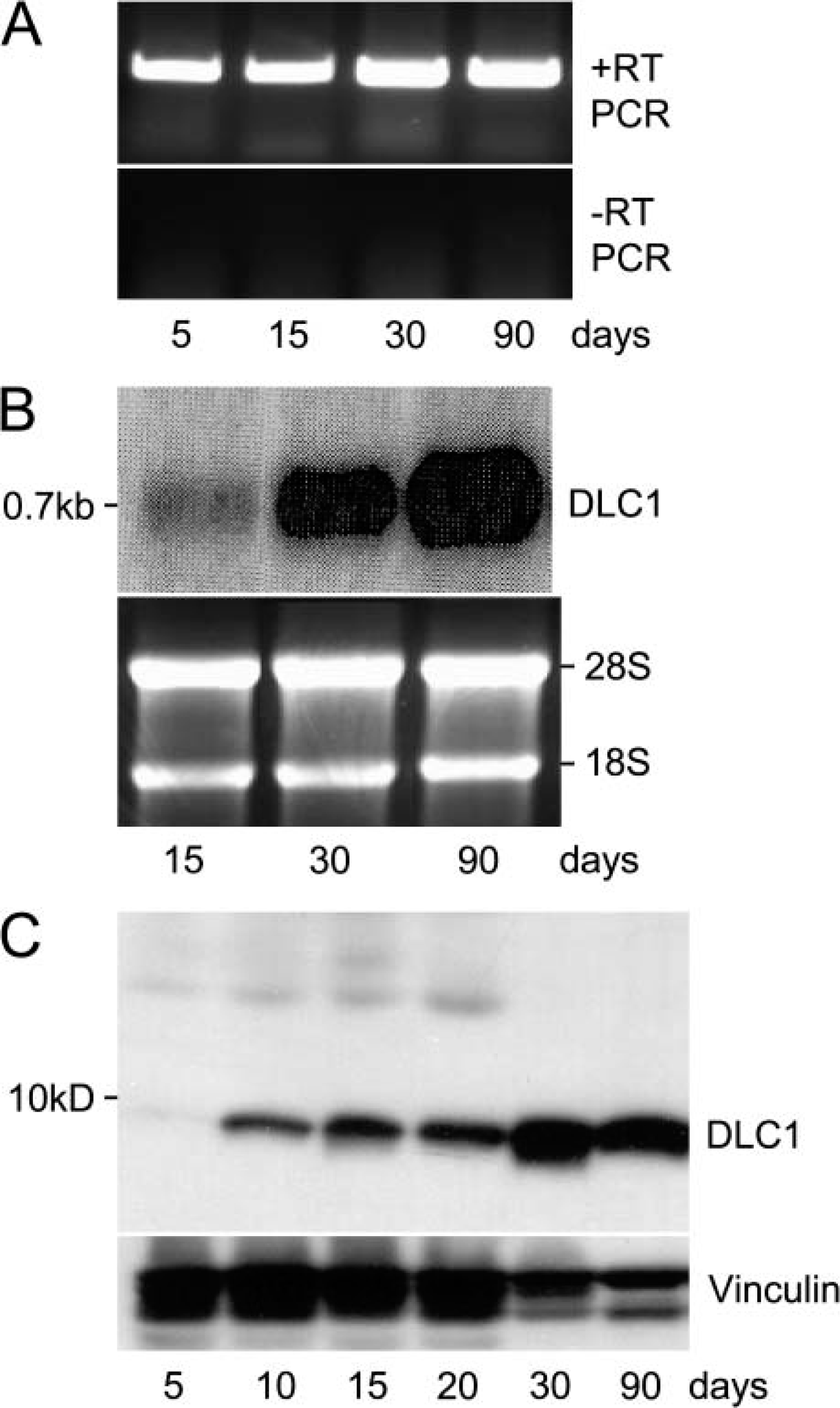
Expression of dynein light chain-1 (DLC1) during the development of testes.
Developmental Regulation of DLC1 in Testes
We next determined the expression profile of DLC1 during the postnatal development of murine testes. We performed RT-PCR analysis and found that DLC1 mRNA was expressed during all developmental stages (Figure 2A). However, Northern blotting revealed a substantial increase in DLC1 expression beginning on the 30th postnatal day, and the expression was even further elevated in 3-month-old adult mice (Figure 2B). Western blot analysis also revealed a corresponding increase in the DLC1 protein beginning on the 30th postnatal day (Figure 2C). In addition, there was an earlier, but smaller, increase in the levels of DLC1 protein in the murine testes between 5 and 10 days of age (Figure 2C).
In general, the biggest difference in murine testes between the 5th and 10th day is in the ratio of somatic cells to germ cells. On the 5th day, the germ cells, which are gonocytes transforming into type A spermatogonia at this stage of development, are few in number, whereas the somatic cells, mostly Sertoli cells, are actively proliferating and are dominant. By the 10th day, germ cells have undergone active proliferation, increased in numbers relative to the somatic cells, and have progressed to the leptotene spermatocyte stage (Bellve et al. 1997). When mice are 30 days old, their testes begin active spermiogenesis. In this context, we speculate that the increase in the level of DLC1 first noticed on the 10th day signifies a higher level of DLC1 expression in mitotically dividing germ cells than in somatic cells, whereas the DLC1 increase at the 30th day is related to spermiogenesis.
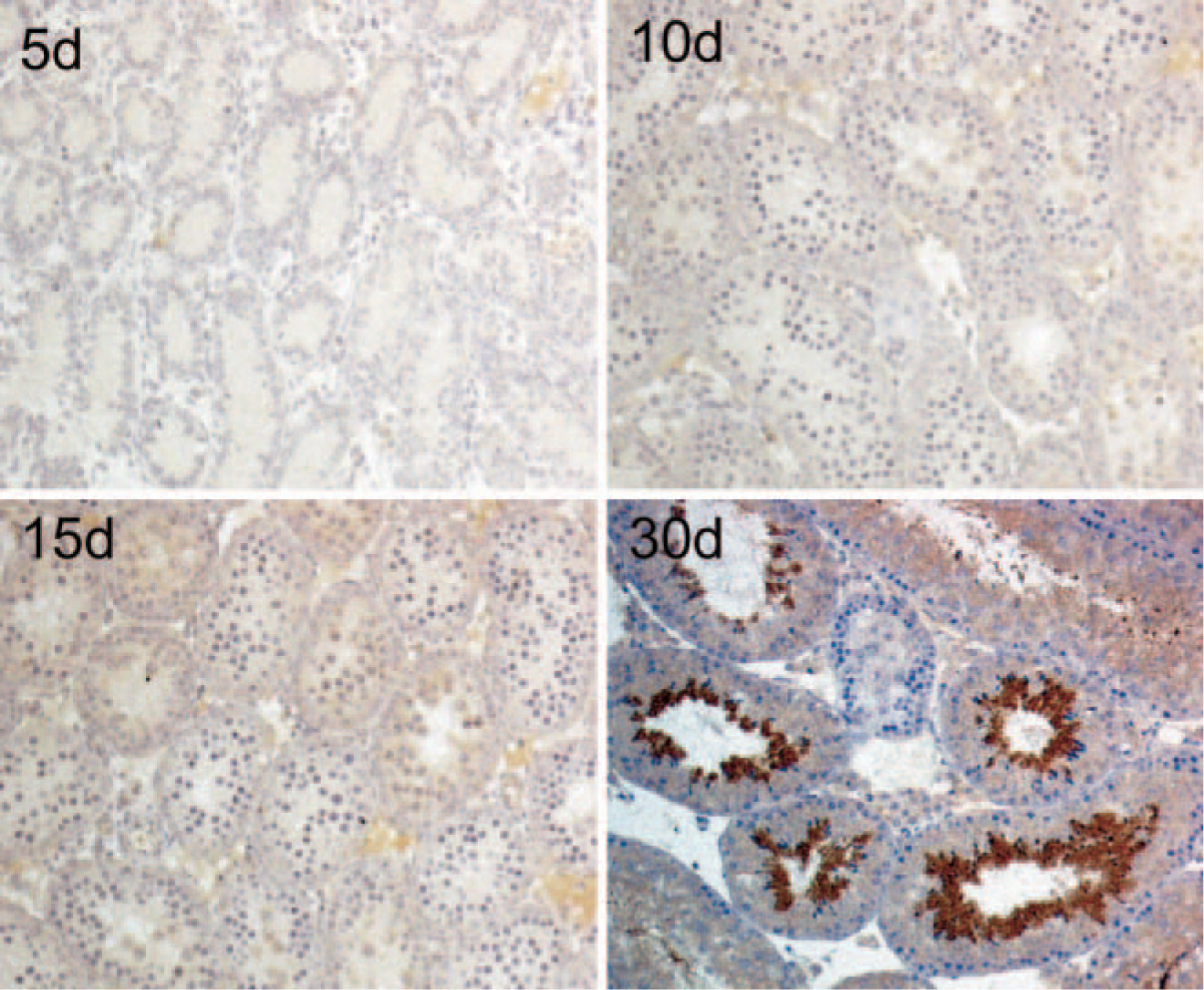
Immunohistochemical detection of dynein light chain-1 expression in different developmental stages of mouse testes. Positive staining is not seen until the 30th day, when active spermatogenesis begins. Samples were examined using Bouin's fixation, indirect horseradish peroxidase-labeled antibody staining, and hematoxylin counterstaining.
Stage-specific Expression of DLC1
We next conducted immunohistochemical analysis to further elucidate the expression pattern of DLC1 in testes. Although Western blotting showed more DLC1 in testes than in other tissues before active spermiogenesis, this was not confirmed by immunohistochemistry. This was probably because of the different detection limits of the two methods. Nevertheless, we found a very strong DLC1 immunoreactivity in the luminal side of the multilayered seminiferous tubules soon after the onset of active spermiogenesis at the age of 30 days (Figure 3).
A careful examination of the staining pattern revealed a stage-specific pattern of DLC1 expression in adult mouse testes. In particular, we found that DLC1 staining first appeared in the nucleus of step 9-elongating spermatids, the stage in which chromatin remodeling begins. Staining became progressively more intense as the spermiogenesis proceeded (Figure 4), because we observed the most intense DLC1 staining in the nucleus in step 12. Interestingly, some spermatid nuclei were positively stained in step 13, whereas DLC1 was predominantly cytoplasmic in other spermatids. From step 14 through the full maturation of spermatids, DLC1 protein was abundant only in the cytoplasm (Figure 4). Interestingly, somatic cells, such as Sertoli cells and Leydig cells, did not exhibit any DLC1-specific immunostaining.
We stained adult rat testes to determine whether the observed stage-specific pattern of DLC1 expression in mouse testes also occurred in other animal species. Interestingly, rat testes also showed the similar pattern of DLC1 expression as seen in mouse testes (Figure 5), with nuclear staining beginning at step 11 spermatids and the transition from nuclear to cytoplasmic staining occurring at step 14.
Pak1 Expression in Mouse and Rat Testes
Recent data suggest that Pak1 regulates DLC1 function by phosphorylating Serine 88 (Vadlamudi et al. 2004). Because Pak1 is involved in regulating cell migration and maintaining cell shape in many situations, we next examined Pak1 expression in mouse testes and whether it correlated with DLC1 expression. Pak1 was not detectable in the testes before the 10th day (Figure 6A). Subsequently, we found a progressive increase in the levels of Pak1 from the 15th postnatal day to adulthood in mouse testes. Moreover, immunohistochemical staining also showed that Pak1 was widely expressed in both mouse and rat testes (Figure 6). In particular, Pak1 immunostaining was observed in pre-leptotene spermatocytes (Figure 6E), in both the nucleus and cytoplasm of later stage elongated spermatids of mouse testes (Figures 6B and 6C). High levels of Pak1 was also detected round spermatids of mouse testes in step 7 (Figure 6D). In rat testes, however, Pak1 was detected only in the cytoplasm, but not the nuclei of elongated spermatids. Pak1 expression was also detected in the Leydig cells of both mouse and rat testes, with the latter having especially high levels (Figures 6E and 6F). In general, the stage-specific expression of Pak1 corresponds to that of DLC1, with the exception of preleptotene spermatocytes and step 7 round spermatids.
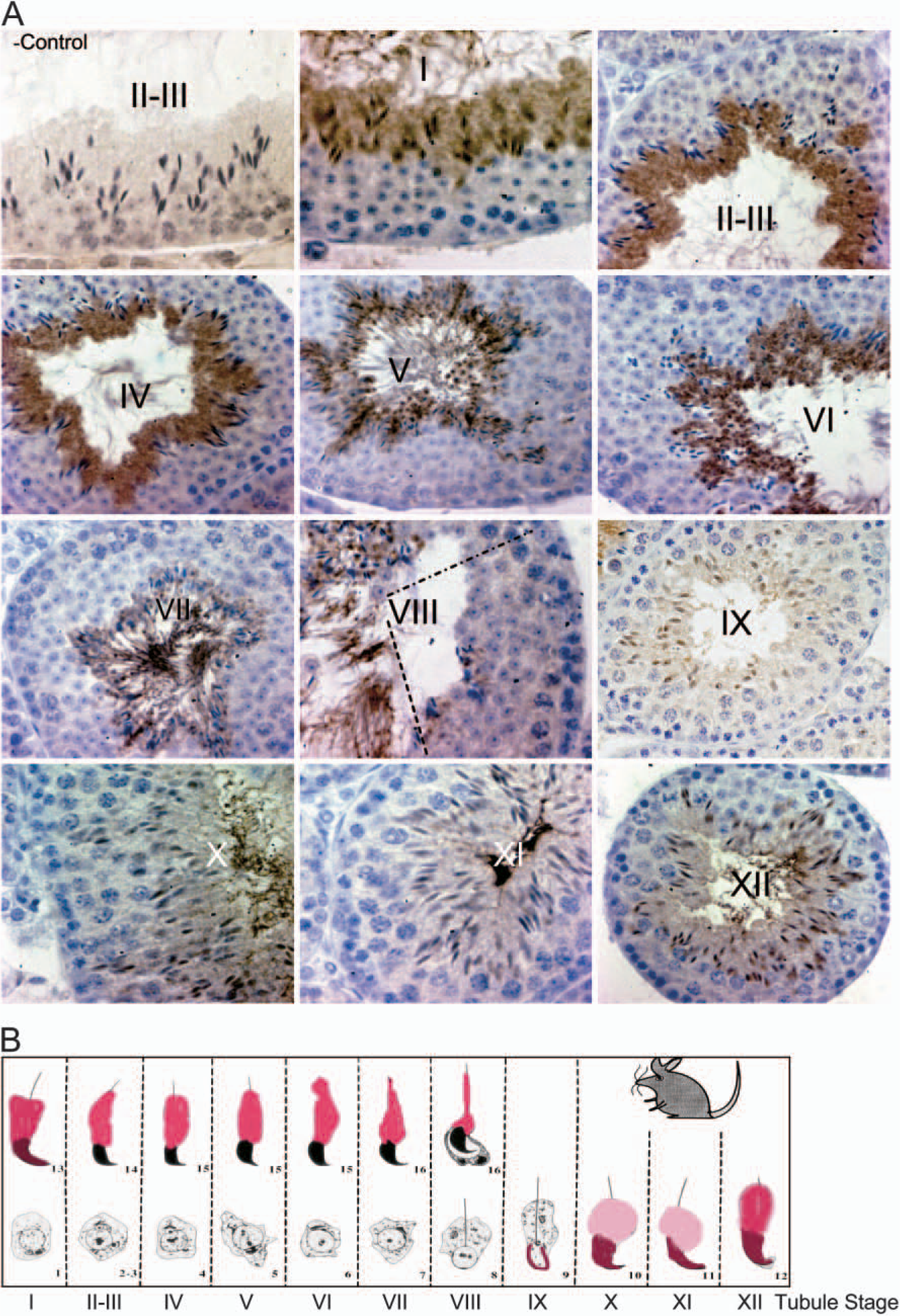
Differential expression of dynein light chain-1 (DLC1) during spermatogenesis.
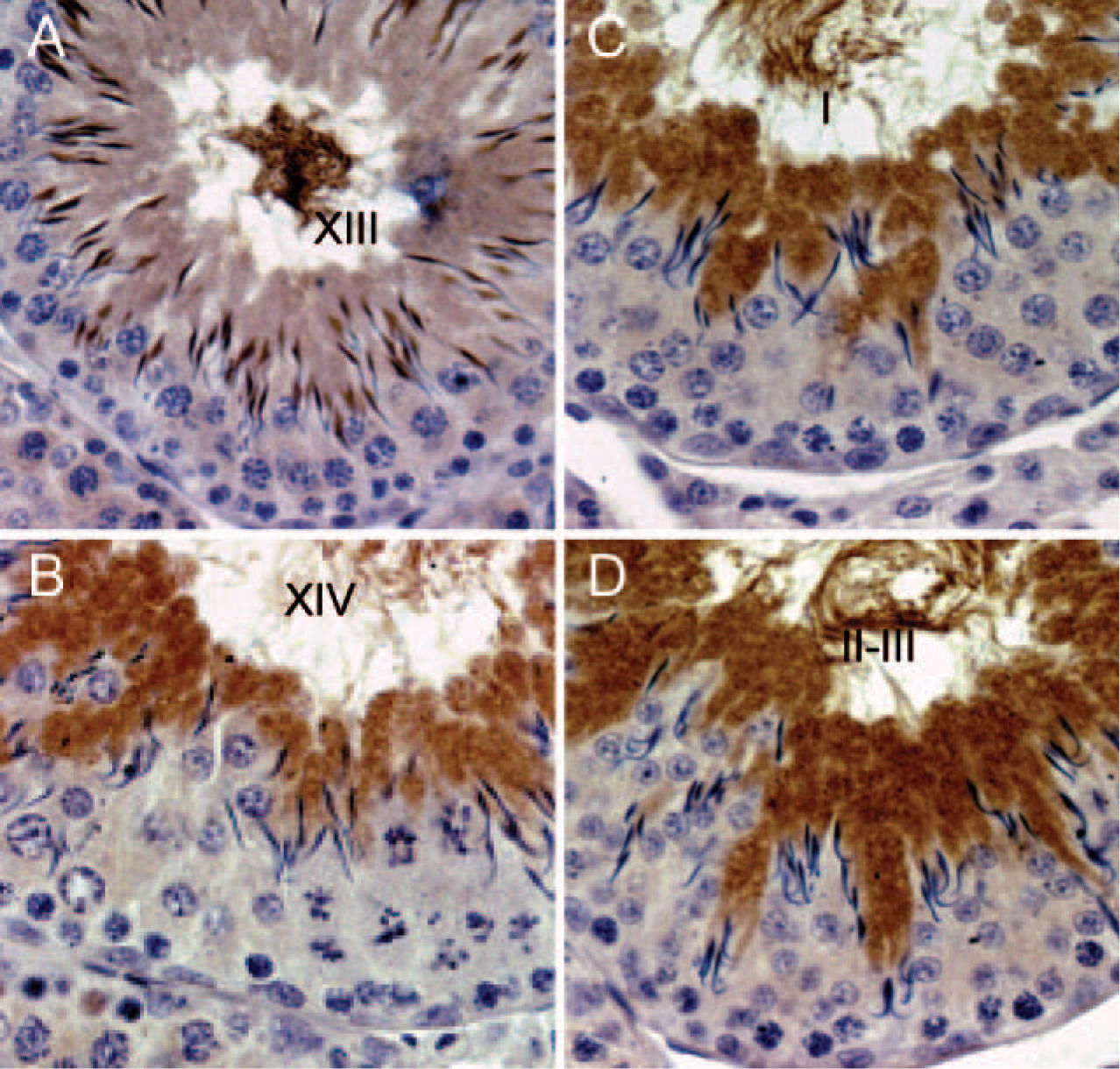
Expression of dynein light chain-1 (DLC1) in rat testes follows the same pattern as that of mouse testes.
Modulation of DLC1 Expression by CREM and Pak1
Having found a stage-specific expression of DLC1 in rodent testes, we next attempted to elucidate the potential mechanism responsible for the observed expression pattern. After step 8, transcription gradually decreases and is totally inactivated at step 11 (Kierszenbaum and Tres 1975). Therefore, the presence of high level DLC1 protein in elongated spermatids cannot be translated from the newly transcribed mRNA. Recent studies suggest that CREM® controls the transcription of a set of genes in the round spermatids, and these transcripts are later translated to protein in the elongated spermatids where they perform their functions (Kistler et al. 1994; Sun et al. 1995; Sassone-Corsi 2000). These findings suggest that DLC1 expression in the testes might also be under the control of CREM. Furthermore, Pak1 signaling has been shown to regulate the expression of a plethora of genes by phosphorylating and transactivating other transcription factors, such as estrogen receptor a and Stat5a (Wang et al. 2002, 2003). Together, these observations led us to hypothesize that Pak1 also regulates DLC1 expression by interacting with other transcription factors.
To test this hypothesis, we first evaluated the expression of CREM® in mouse testes. As previously reported (De Cesare et al. 1999), the expression of CREM in mouse testes was confined to the nucleus of round spermatids, especially in a high level at step 7 (Figure 7), during which high level of Pak1was also detected (Figure 6D). Next, we cloned the DLC1 promoter from BAC clones into the pGL3 luciferase vector. Sequence analysis done with TransFac software (Biobase; Braunschweig, Germany) revealed that there were two putative CREB binding sites on the DLC1 promoter (Figure 8A). We next examined the effect of CREM and Pak1 on DLC1 promoter activity using a COS-7 cell co-transfection assay. This showed a significant increase in DLC1 promoter activity when Pak1 and CREM were cotransfected, but neither Pak1 nor CREM alone affected DLC1 promoter activity (Figure 8B).
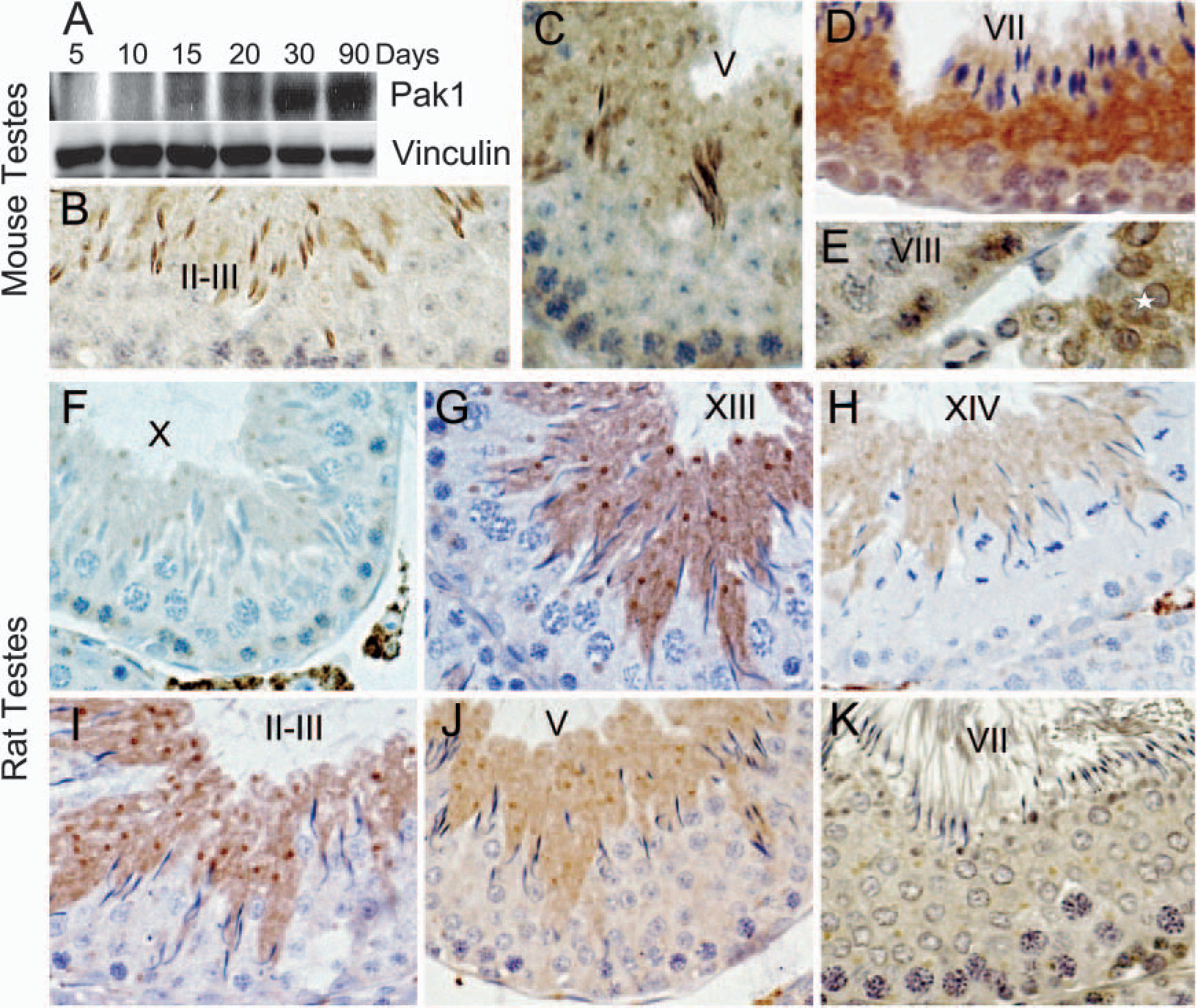
p21-activated kinase 1 (Pak1) expression in mouse and rat testes.
Discussion
We found that DLC1 was expressed in a developmentally regulated and stage-specific manner in mouse and rat testes. Immunohistochemically, DLC1 protein initially appeared in the nucleus of the early elongating spermatid and later was expressed in the cytoplasm. We further found that Pak1 was coexpressed with DLC1 and present in round spermatids, preleptotene spermatocytes, and Leydig cells. In addition, Pak1, in collaboration with CREM, regulated DLC1 promoter activity.
Given that the testis is a highly dynamic organ in which cells are constantly proliferating and differentiating, it is very likely that dynein complex components are involved in these processes. Indeed, two other components of the dynein light chain, Tctex1 and Tctex2, have also been documented to be expressed in testes and found to be a component of sperm flagella. Tctex1 and Tctex2 are believed to be involved in the non-Mendelian transmission of the t-haplotype in mice (Criswell and Asai 1998; Harrison et al. 1998). Though not fully understood, transmission ratio distortion (also called meiotic drive) is believed to be related to abnormal spermiogenesis (Harrison et al. 1998). The resultant abnormally developed sperm have little capacity to fertilize ova. Our study provided detailed documentation of DLC1 expression in the spermatids, which has several important functional implications and sheds some light on the mechanism that can go awry in spermiogenesis: first, DLC1 is not only a cytoplasmic protein, but it may also function in the nucleus, as has been recently shown in cultured cells using subcellular fractionation methods (Kaiser et al. 2003). In our study, we postulated that DLC1 plays an important motor function during early-stage chromatin condensation. Second, after initial chromatin condensation, the spermatid needs to streamline its shape and get rid of excess cytoplasm. All of these processes require active movement of the microtubule-associated cytoskeletons. In addition, DLC1 may also play a role in reshaping spermatid. Third, DLC1 may provide an important motor drive for the final release of mature sperm into the lumen. The coexpression of Pak1 and DLC1 in elongated spermatid stage suggests that DLC1 maybe activated through phosphorylation by Pak1.
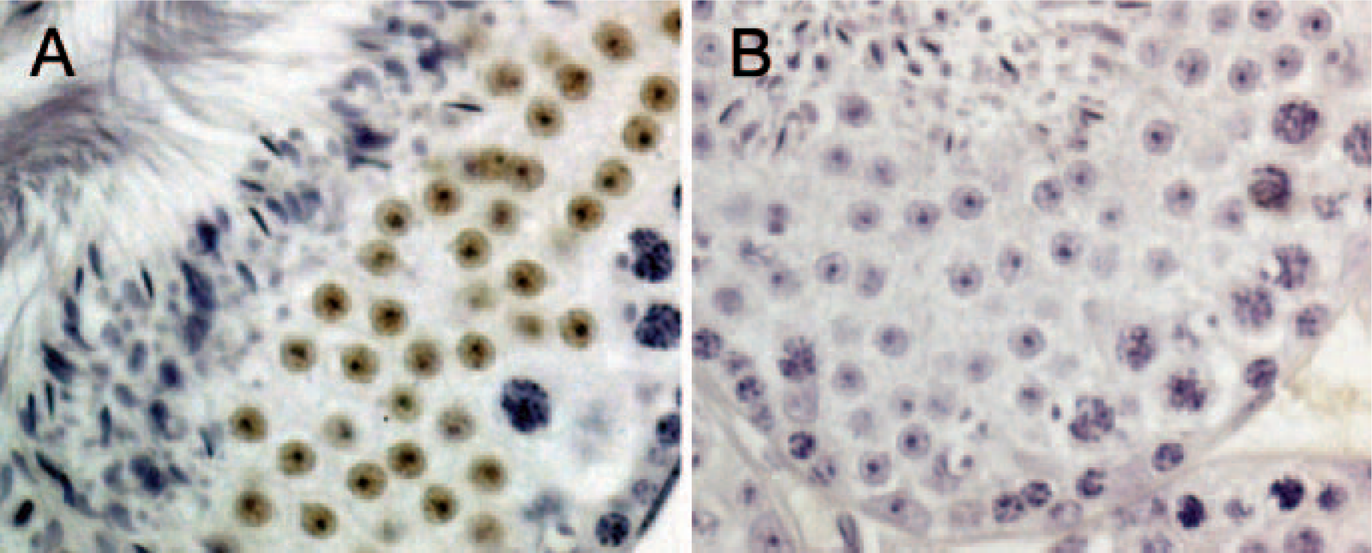
cAMP-responsive element modulator (CREM) is expressed in round spermatids of mouse testes.
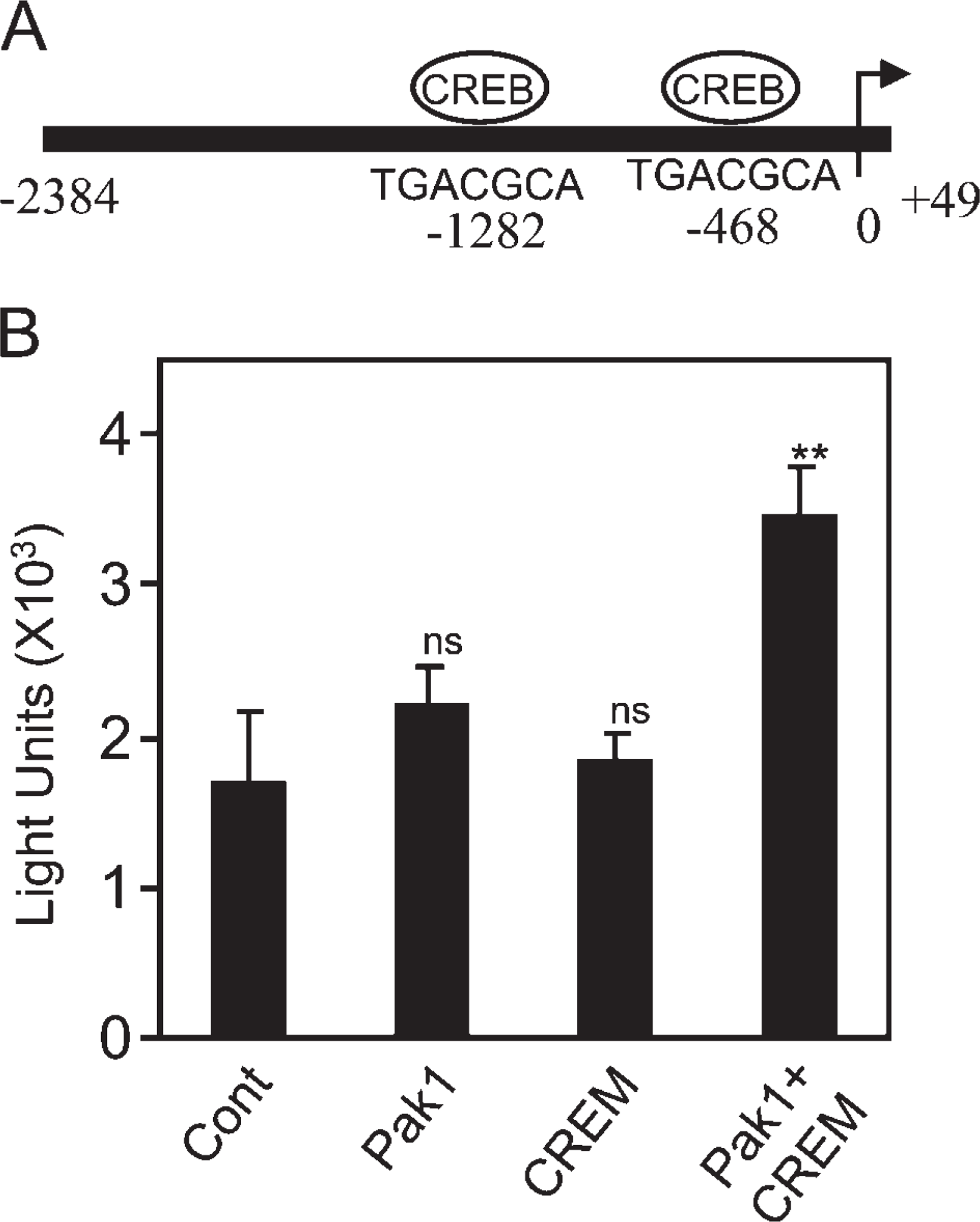
p21-activated kinase 1 (Pak1) and cAMP-responsive element modulator (CREM) in collaboration stimulate dynein light chain-1 (DLC1) promoter activity.
A time lag between transcription and translation is common in testes. It is reasonable to speculate that the transcription of these genes that encode proteins expressed in elongated spermatids must be inactivated in these spermatids, and, therefore, the coding mRNAs must be completed earlier. In addition to DLC1, protamines and transitional proteins are other good examples of tightly controlled protein expression (Hecht 1998). The pretranscribed mRNA remains free, without polysomal loading. The repression of translation of these mRNAs is likely to be related to RNA binding proteins that recognize the 3'-UTR region of the transcripts (Hecht 1998). The repression of earlier translation is important for spermiogenesis. In the case of protamine, when the 3'-UTR was modified and protamine protein was expressed in the round spermatids, the mouse became infertile because of the structural changes in the sperm chromatin (Braun et al. 1989). The time lapse between DLC1 mRNA transcription and protein expression also raises the possibility that DLC1 is important in chromatin condensation and the final morphogenesis of sperm.
In summary, we found that DLC1 is highly expressed in elongated spermatids, initially in the nucleus and subsequently in the cytoplasm. The stage-specific expression of DLC1 in elongated spermatids suggests that DLC1 may be involved in chromatin condensation, morphologic changes in nucleus, the shearing off of excess cytoplasm, and release of sperm into the lumen.
Footnotes
Acknowledgements
The study was supported by NIH Grants CA-90970 and CA-80066 (RK), and in-part by Cancer Center (CORE) Support Grant NIH-NCI CA-16672.
We thank Dr. Paolo Sassone-Corsi for kindly providing us with the CREM expression vector and antibody, and David Galloway for editing of the manuscript.