
Abstract
Chagas disease is an incurable illness caused by the protozoan Trypanosoma cruzi. Cardiomyocytes represent important targets for the parasite infection and alterations in their physiology were reported. Because endocytosis is involved in different cellular events and guanosine triphosphatase (GTPase) Rab proteins play important roles in various aspects of the membrane traffic, our aim was to characterize the expression of Rab proteins in T. cruzi-infected cardiomyocytes, which displayed a downregulation of Rab7 and Rab11, whereas the expression of Rab5a was maintained in the infected cultures even after longer periods of parasite internalization, but early endosome antigen 1 was partially downregulated. The parasite infection also decreased the uptake of fluid phase ligands by the cardiac cultures. The regulation of GTPase proteins and effector molecules can contribute to the altered physiology of the host cells by modifying the normal incoming of nutrients as well as interfering with other important events related to the endocytic pathway.
Keywords
I
The endocytic pathway shuttles materials along specific but varied routes between the plasma membrane, endosomes, lysosomes, and Golgi (Gruenberg 2003). It is composed by complex and heterogeneous organelles consisting of an extensive network of tubulovesicular membranes extending from the cell surface to the peri-nuclear region (Mukherjee et al. 1997). Shortly after internalization, membrane-bound transport vesicles fuse with the early sorting endosomes, where the internalized ligands, membrane proteins, and lipids are sorted and either returned to the plasma membrane (directly or via recycling endosomes as fast and slow routes, respectively), or transported to the late endosomes and then to lysosomes for final degradation. Alternative pathways also include the transport of molecules to the trans-Golgi network (Gruenberg 2003).
Guanosine triphosphatase (GTPases) are small guanosine triphosphate (GTP)-binding proteins that participate in a variety of cellular events, including cell activation, proliferation, and differentiation (Etienne-Mannerville and Hall 2002). Regarding membrane traffic, the Rab GTPases and their divalent effector proteins are implicated in cargo selection, vesicle budding, vesicle motility, tethering, docking, fusion membranes, and functional organelle identity (Zerial and McBride 2001). Individual Rabs are localized in different organelles of the secretory and endocytic pathways defining compartment identity. However, a significant degree of colocalization occurs within the same organelle, for example while sorting endosomes are enriched in Rab5 and Rab4 domains, with a smaller ratio of Rab11-positive regions, the recycling endosomes are mostly composed of Rab11 and Rab4 domains (Miaczynska and Zerial 2002). On the other hand, late endosomes are characterized by the presence of Rab7 and Rab9, which control the transport between early and late endosomes and govern the formation of carriers destined to the Golgi, respectively (Rodman and Wandinger-Ness 2000).
Some pathogens can modulate the intracellular trafficking of the host cells to survive and replicate (Walburger et al. 2004). Trypanosoma cruzi is an obligatory intracellular protozoan that causes Chagas disease, a chronic, incurable illness, which is an important cause of morbidity and mortality, infecting up to 18 million people in South and Central America (WHO 2002). Cardiac muscle cells are considered important targets for T. cruzi infection, and alterations in the physiology of the cardiomyocytes have been largely reported (Soeiro et al. 1995; Calvet et al. 2004). Because (1) endocytosis takes place in many important cellular functions (Conner and Schmid 2003) and (2) the molecular characterization of the GTP-binding proteins represents a powerful tool for identifying and understanding the endocytic routes of eukaryotic cells, our aim was to characterize the expression of the GTPases Rab5a (and its effector protein early endosome antigen 1 [EEA1]), Rab11, and Rab7 in both uninfected and T. cruzi-infected cardiomyocytes and further analyzed if the infection affects the endocytosis of fluid phase ligands by the cardiomyocytes.
Materials and Methods
Reagents and Antibodies
Anti-EEA1, anti-Rab11, and anti-Rab7 polyclonal antibodies were a generous gift from Dr. Gareth Griffiths (EMBL; Heidelberg, Germany). Anti-Rab5a antibodies were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA). Anti-rabbit IgG rhodamine conjugated antibody was purchased from Jackson ImmunoResearch Laboratories Inc. (West Grove, PA). Anti-rabbit IgG FITC conjugated antibodies were purchased from Sigma Chemical Co. (St Louis, MO). Alkaline phosphatase-conjugated goat anti-rabbit IgG was purchased from Southern Biotech (Birmingham, AL). All other reagents were purchased from Sigma.
Cell Cultures
Primary cultures of embryonic cardiomyocytes (CM) were obtained from Swiss mice following the method previously described (Meirelles et al. 1986). Isolated CM were adhered on gelatin-coated culture plates and were sustained in DMEM supplemented with 5% FBS, 2.5 mM CaCl2, 1 mM
Parasites
Trypomastigote forms of T. cruzi Y strain (representative of the lineage II that circulate in the domestic biological cycle of T. cruzi) and Dm28c clone (representative of the lineage I mostly related to the sylvatic cycle) were used throughout the experiments (Devera et al. 2003). Cell culture-derived trypomastigotes (Dm28c) were isolated from the supernatant of Vero cell line, previously infected with metacyclic forms as described (Calvet et al. 2004). Bloodstream trypomastigotes (Y stock) were harvested by heart puncture from T. cruzi-infected Swiss mice at the parasitemia peak day (Meirelles et al. 1982).
Infection Assays
After 24 hr of platting, CM were infected with trypomastigotes forms of T. cruzi (Y or Dm28c stocks) for 2 and 24 hr at 37C, respectively, employing a parasite:host cell ratio of 10:1. After this host cell-parasite contact, the cultures were washed to remove unbound parasites and the infection was followed for different periods for 72 hr.
Indirect Immunofluorescence Assays
Uninfected and T. cruzi-infected CM were washed with PBS, pH 7.4, and fixed for 20 min at 4C in 4% paraformaldehyde in PBS. After permeabilization for 10 min with 0.25% Triton X-100 in PBS, the samples were incubated for 30 min in 4% BSA to block unspecific labeling. The cultures were then incubated for 60 min at room temperature with the appropriated primary antibodies (anti-Rab5a, -Rab7, -EEA1, and -Rab11) diluted 1:250 in PBS containing 0.25% Triton X-100 plus 4% BSA. After two washes with PBS containing 0.25% Triton X-100, the samples were incubated for 30 min at 37C with anti-rabbit IgG rhodamine-conjugated antibodies (1:800). To visualize the striated cells, the cardiac myocytes cultures were further incubated for 30 min with 5 μg/ml phalloidin-FITC, for staining the actin filaments. The cultures were incubated with 10 μg/ml 4,6-diamidino-2-phenylindole for DNA staining, which enables the visualization of the parasites and the heart cells nuclei, mounted with 2.5% 1.4-diazabicyclo-(2.2.2)-octane and examined using a Zeiss Axioplan 2 microscope equipped with epifluorescence (Zeiss Inc; Thornwood, NY) or confocal laser scanning microscope (Bx512; Olympus Latin America Inc., Farmanguinhos/FIOCRUZ).
Flow Cytometry Analysis
For evaluation of Rab5a, Rab7, Rab11, and EEA1 expression, 106 uninfected and T. cruzi-infected cardiomyocytes (24 hr of parasite:host cell interaction) were washed with PBS and fixed for 20 min at 4C with 2% paraformaldehyde in PBS. After washing, the samples were blocked for 30 min in DMEM containing 5% FBS and 5% sheep sera. After permeabilization for 10 min with 0.1% saponin, the cells were incubated for 30 min at room temperature with the primary antibodies (anti-Rab5a, -Rab7, -EEA1, and -Rab11) diluted 1:100 in PBS containing 0.1% saponin. After washes, the CM were incubated for 30 min at room temperature with the secondary antibodies coupled to fluorescein. The samples were finally washed twice in PBS containing 0.1% saponin and once in PBS before the flow cytometry analysis. Data acquisition and analysis were performed using a FACS-Calibur flow cytometer (Becton Dickinson; San Jose, CA) equipped with Cell Quest software (Joseph Trotter; Scripps Research Institute, San Diego, CA). A total of 10,000 events were acquired in the region previously established as that corresponding to the cardiomyocytes. Alterations in the fluorescence for each protein were quantified using an index of variation (IV) obtained by the equation (MT/MC) × 100 − 100, where MT is the median of fluorescence for T. cruzi-infected and MC corresponds to uninfected cardiomyocyte's fluorescence, respectively. Negative controls were performed by the omission of the primary antibodies or by incubating the samples with nonimmunized sera.
Western Blotting Analysis
Cell lysates (20 μg of protein) were separated by SDS-PAGE (15% polyacrylamide gels). Proteins were transferred to 0.45 μm nitrocellulose membrane (Gibco BRL; Gaithersburg, MD) and blocked overnight at room temperature with PBS containing 5% denatured milk plus 0.1% Tween 20. Blots were incubated for 1 hr at 37C with antibodies against the Rabs diluted 1:500 in the same buffer. After the antibody incubation, the membranes were rinsed and subsequently incubated for 30 min at room temperature with anti-rabbit IgG antibodies coupled to alkaline phosphatase (1:3000). After rinsing, the enzymatic activity was revealed with 0.17 mg/ml 5-bromo-4-chloro-3 indolyl phosphatase (BCIP; Sigma) and 0.34 mg/ml nitroblue tetrazolium (NBT; Sigma) in substrate buffer (0.1 M Tris, 100 mM NaCl, 50 mM MgCl2, pH 9.5).
Endocytic Assays
Uninfected and T. cruzi-infected cardiomyocytes (48 hr of infection with Y stock) were allowed to take up 5 mg/ml-1 fluorescein isothiocyanate dextran (FITC-Dx) for 3 min at 37C. Afterwards, the cultures were extensively washed at 4C and further chased for 60 min at 37C in FITC-Dx free culture medium. The samples were then either fixed for 20 min at 4C in 4% or 2% paraformaldehyde and processed for fluorescent microscopy or flow cytometry, respectively, as described previously.
Results
Structural and functional alterations have been reported in heart cells during T. cruzi infection, mostly from the upmodulation and downmodulation of the cardiac genes and proteins expression. To verify if Rab GTPases could be regulated during the parasite invasion of cardiac cells in vitro, the expression of Rab5a, Rab7, and Rab11 was evaluated in both uninfected and T. cruzi-infected cardiomyocytes by immunocytochemical approaches. The data obtained during the interaction of CM with trypomastigotes from both stocks (Y and Dm28c) were similar independently from the GTPases analyzed (data not shown).
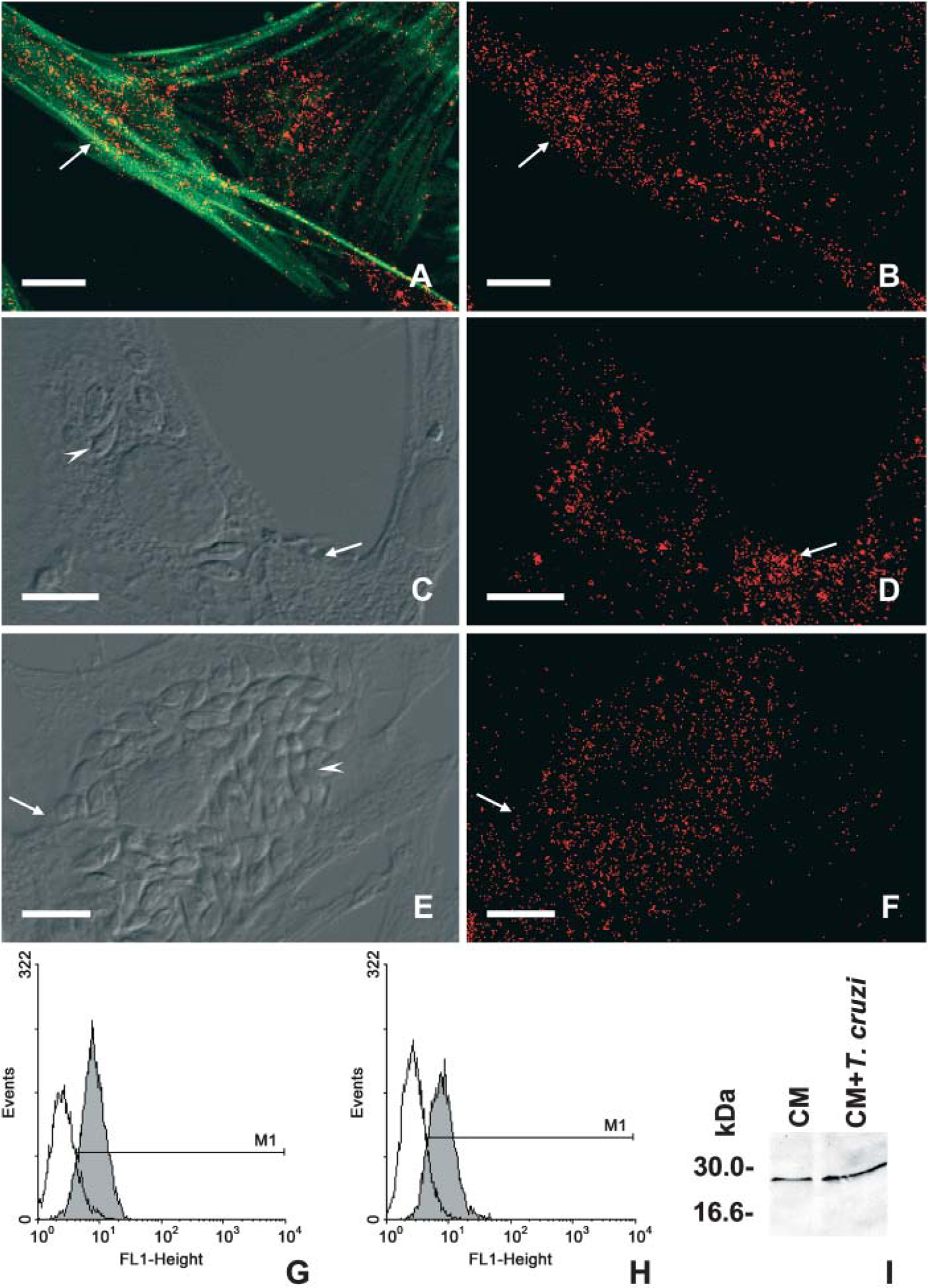
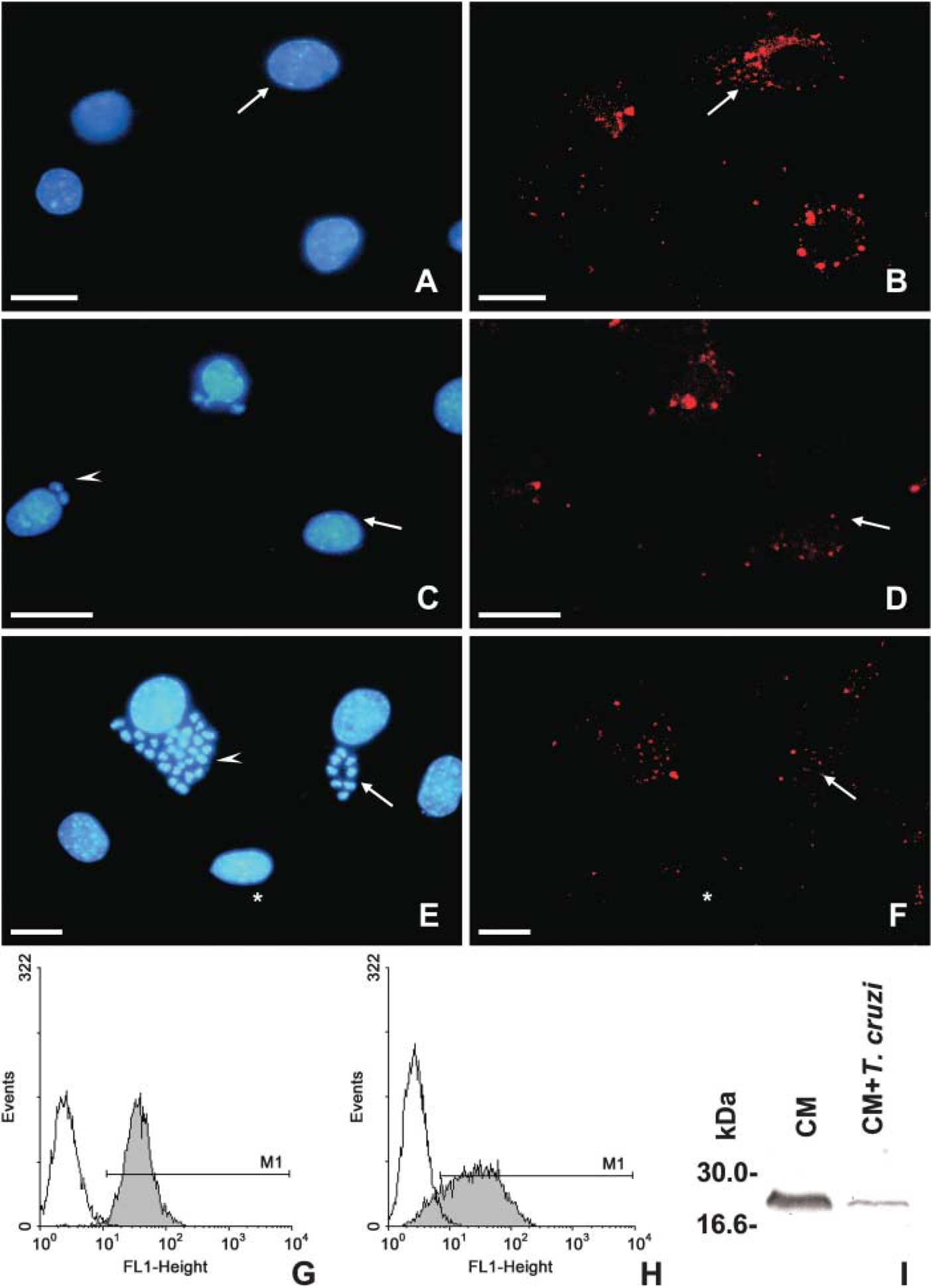
Rab5a, an early endosome-specific GTP-binding protein, displayed a punctate labeling at the cell periphery and throughout the whole cytoplasm of the uninfected CM as predicted for early compartments of mammalian cells (Figures 1A and 1B). Confocal laser scanning microscopy analysis revealed a similar pattern of Rab5a labeling in both uninfected (Figure 1B) and in T. cruzi-infected host cells after 24 (Figure 1D) and 72 hr of interaction (Figure 1F), which display ∼31 ± 10 and 40 ± 5% of infection, respectively. Next, flow cytometry analysis was performed to quantitatively assay the expression of Rab5a in both uninfected and T. cruzi-infected cardiomyocytes. Our data did not show differences regarding Rab5a expression in cardiac cells after 24 hr of parasite infection (Figure 1H) as compared with the uninfected cultures (Figure 1G). Additionally, Western blotting analysis revealed a band of ∼29 kDa in both uninfected and T. cruzi-infected cells, and showed that Rab5a expression was sustained in the infected cardiomyocyte cultures even after 72 hr of parasite invasion (Figure 1I). Because EEA1 is a key factor controlling vesicle fusion, being an effector molecule of Rab5, we next evaluated EAA1 expression during the infection of cardiomyocytes by T. cruzi. Our immunocytochemistry studies showed that EEA1 labeling was present throughout the cytoplasm of the uninfected cardiomyocytes, including the perinuclear region (Figures 2A and 2B). Infected cardiac cultures showed a decrease in the expression of EEA1 in both nonparasitized and parasitized host cells after 24 (Figure 2C and 2D) and 72 hr (Figures 2E and 2F) of parasite internalization. The quantitative analysis by flow cytometry assays showed that after 24 hr of parasite uptake, ∼20% (M2) of the CM culture presented a remarkable reduction in the fluorescent labeling, reaching an IV value of 86% (Figure 2H) as compared with the uninfected cultures (Figure 2G).
We next investigated the expression of Rab11, a GTPase involved in the mammalian recycling pathway. In uninfected cardiomyocytes, the analysis of recycling endosomes detected by anti-Rab11 antibodies showed a variable labeling in large vesicles and web elements distributed throughout the cell cytoplasm, besides a perinuclear staining (Figures 3A and 3B). After 24 hr of parasite invasion, the infected CM cultures displayed a decrease in Rab11 staining, which was independent from the presence of intracellular parasites (Figures 3C and 3D). After 72 hr of parasite internalization, the reduction in Rab11 labeling was even more accentuated in the infected cultures, whose cells only displayed a faint labeling (Figures 3E and 3F). However, labeled intracellular parasites could be noted (Figure 3F, arrow). Flow cytometry analysis confirmed that, after 24 hr of T. cruzi invasion, the cardiac Rab11 labeling displayed a striking decreased in the median of fluorescent reaching 39% of reduction in the index of variation (Figure 3H), as compared with uninfected cultures (Figure 3G).
Analysis by fluorescent microscopy
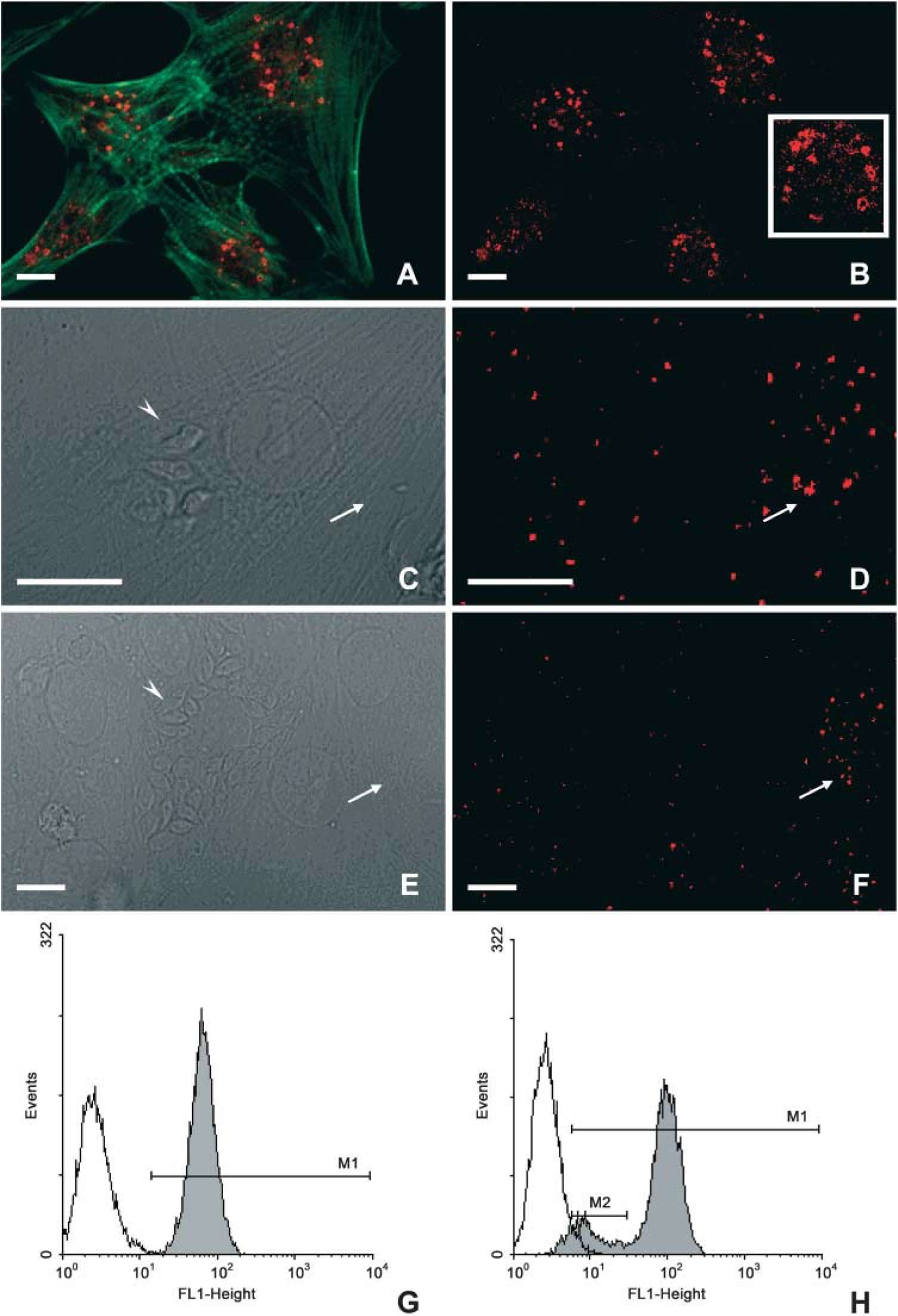
When we analyzed the expression of Rab7, a GTPase specific for late compartments, we noted that uninfected CM displayed a bulk labeling near the center of the CM, in close proximity to the cell's nuclei (Figures 4A and 4B). After 24–72 hr of T. cruzi infection, a meaningful decrease in Rab7 expression in the infected cultures was noticed (Figures 4C, 4D, 4E, and 4F), even in nonparasitized host cells (Figures 4E and 4F, asterisks). Intracellular parasites were strongly labeled (Figures 4D and 4F, arrow). In all fluorescent approaches, the control samples performed only with secondary antibodies alone or with nonimmune sera resulted in total absence of the fluorescent staining (Figures 3I and 3J).
The quantitative analysis by flow cytometry confirmed the decrease of Rab7 expression in infected cultures (Figure 4H) as compared with the uninfected ones (Figure 4G), reaching an IV of 30%. Western blotting analysis further confirmed the reduced GTPase expression as early as 24 hr of T. cruzi internalization (Figure 4I).
Analysis by fluorescent microscopy
Analysis by fluorescent microscopy
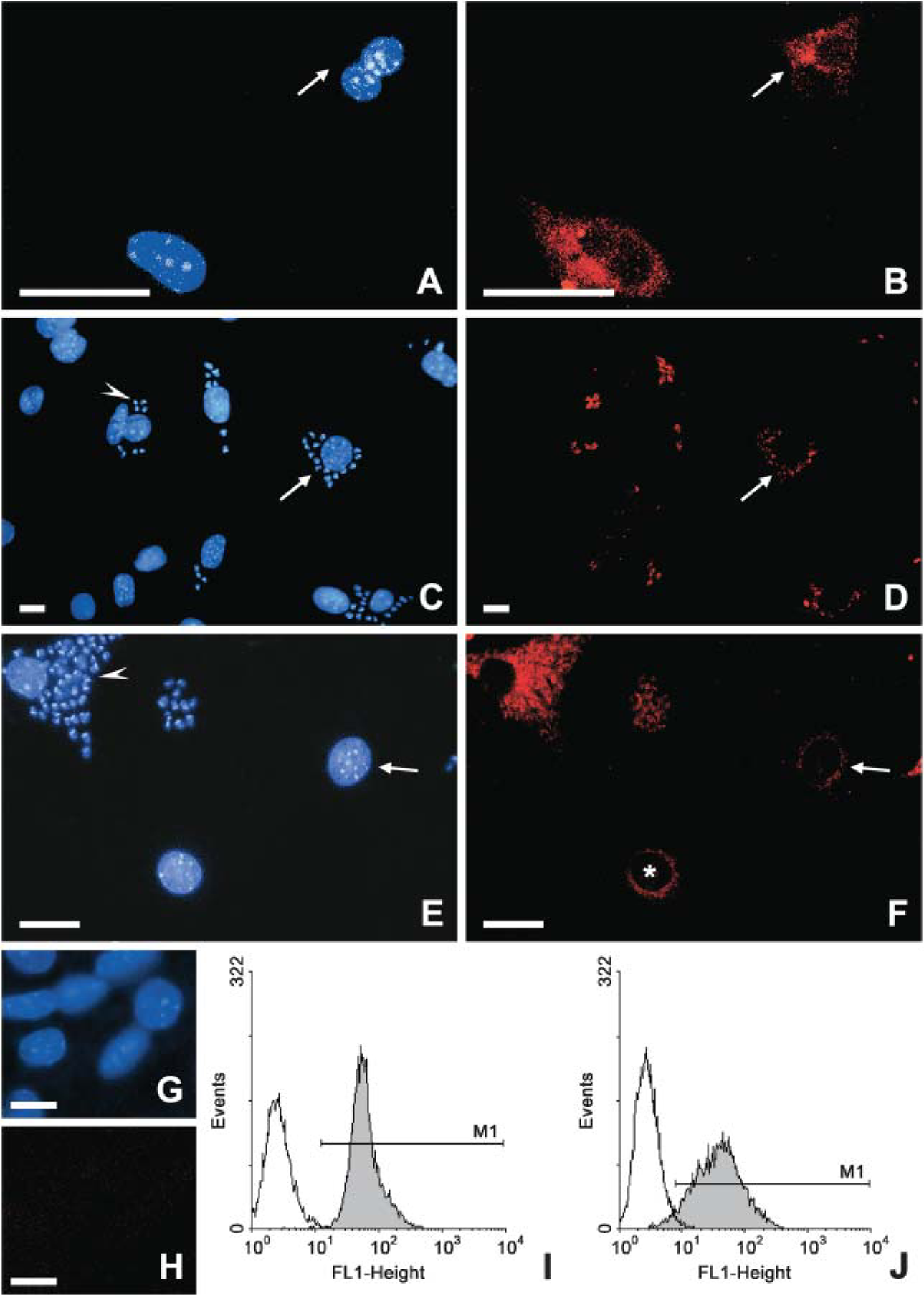
To address whether the noted regulation in the Rabs expression could result in altered molecule acquisition by T. cruzi-infected cardiomyocytes, we finally assayed the internalization of fluorescent endocytic tracers by fluorescent microscopy (Figure 5). In these assays, CM were infected or not with trypomastigotes of T. cruzi and then incubated with FITC-Dx. We observed a decrease in the number and intensity of fluorescent endocytic compartments of the T. cruzi-infected cultures (Figure 5E) as compared with the uninfected cell cultures, which displayed a large number of compartments labeled by FITC-Dx mainly localized in the perinuclear regions, corresponding to the late endosomes (Figure 5B). The reduced incorporation of FITC-Dx was also observed on uninfected neighbors cells present in the infected cultures. Flow cytometry analysis confirmed that T. cruzi-infected cardiomyocytes incorporated less FITC-Dx (Figure 5F) as compared with uninfected cardiac cells (Figure 5C), reaching an IV value of 25%.
Collectively, our data indicate that during T. cruzi infection in vitro, the cardiomyocytes present important alterations in the expression of GTPases and their effector proteins, which can, at least in part, be responsible for their impairment in the uptake of endocytic tracers.
Discussion
Rabs are small GTPases of the Ras super family that orchestrate the sequence of fusion events besides acting as membrane domain organizers. They behave as molecular switches cycling between GTP-bound (active) and GDP-bound (inactive) state (Zerial and McBride 2001). Considering that sorting and recycling activities of the early endosomes are fundamental to the proper cellular function and growth (De Renzis et al. 2002), we initiated our study by characterizing the expression of Rab5a, an isoform of Rab5, both in uninfected and T. cruzi-infected cardiomyocytes, which are one of the target cells in Chagas disease.
Analysis by fluorescent microscopy
Rab5 is a GTPase present in sorting endosomes, directing their fusion and motility and being also implicated in vesicle formation (Clague and Urbe 2001). EEA1, its effector molecule, plays a role in tethering two compartments, which leads to their fusion (Simonsen et al. 1998). The expression and localization of Rab5a were presently performed through different methodology, such as scanning laser confocal microscopy, Western blotting, and flow cytometry. Both the qualitative and the quantitative analysis showed that the expression of Rab5a was quite unaltered in cardiomyocytes after T. cruzi invasion. However, although the levels of Rab5a were maintained, the analysis of EEA1 revealed a large impaired expression of this effector molecule, displaying as high as 86% of IV reduction in ∼20% of the infected culture population. The downregulation of EEA1 can contribute to an altered vesicle budding, transporting, tethering, and docking events in the early endosomes, and consequently diminish the normal incoming of nutrients and other endocytic ligands. In fact, our data showed an impairment of a fluid-phase ligand by cardiac cells during their interaction with T. cruzi. Our unpublished results performed by both flow cytometry and confocal laser scanning microscopy confirmed that the infection also impairs the receptor-mediated endocytosis of transferrin and horseradish peroxidase by the cardiac cells. The physiology significance of this altered endocytic process in T. cruzi-infected cardiomyocytes is now under investigation.
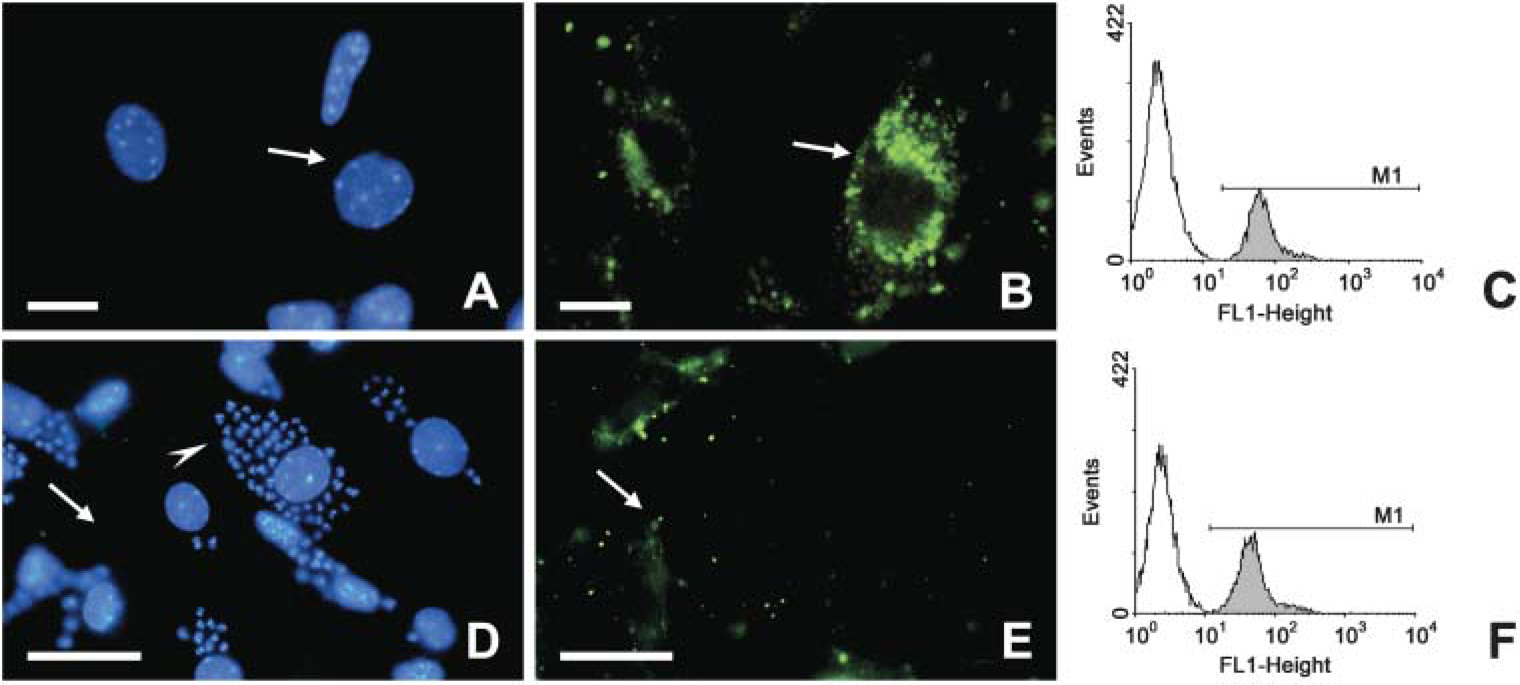
Analysis of the endocytosis of dextran-FITC in uninfected
Because other GTPase proteins could also be regulated during the parasite invasion, we further analyzed the expression of Rab11 and Rab7. In addition to EEA1 alteration, we found that both Rab11 and Rab7 expression were downregulated during T. cruzi infection, lowering ∼39% and 30% of the median fluorescence, respectively, as compared with the overall expression of the uninfected cardiac cultures.
The altered levels of Rab GTPase expression in cardiomyocytes during their infection by T. cruzi could contribute to the host impaired physiology, such as we noted during the defective endocytosis of fluid-phase ligands by the infected cardiac cells. In fact, alterations in the Rab proteins activities can be an important factor in the pathogenesis of diseases caused by intracellular pathogens (Seabra et al. 2002). Uptake of pathogens into phagocytic vacuoles or phagosomes is an essential component of the innate response being central for microbial killing and antigen presentation (Sacks and Sher 2002). The phagosomes mature by the acquisition of microbicidal properties, which involves sequential fusion events with compartments of the endocytic pathway (Vieira et al. 2003). Bacteria, fungi, and protozoan have the ability to modulate Rab trafficking and expression as part of strategies to subvert the host response (Hackstadt 2000). Although Coxiella brunetti survive in acidified phagosomes (Maurin et al. 1992), promastigotes of Leishmania donovani delay the association of late compartments and Rab7 acquisition to their phagosomes until their conversion to amastigotes (Scianimanico et al. 1999). Other examples are Legionella pneumophila and Toxoplasma gondii, which survive and multiply in compartments that do not mature into phagolysosomes (Hackstadt 2000). Live Salmonella also bypass the normal endocytic pathway by modulating the expression of different Rab proteins. In this case, their phagosomes retain Rab5 and Rab18 but are refractory to Rab7 and Rab9, avoiding the lysosomal fusion (Hashim et al. 2000). Similarly, Listeria monocytogenes delays phagosome maturation and subsequent degradation, prolonging its provisory survival inside the phagosomes and assuring the enhancement of the bacterial viability as a prelude to escape into the cytoplasm (Alvarez-Dominguez et al. 1997).
Pathogenic mycobacteria have evolved eukaryotic-like signal transduction mechanisms capable of modulating the host cell trafficking (Walburger et al. 2004). Some lipids moieties from mycobacterias have been related to the arrest of their phagosomal maturation (Vergne et al. 2004). Mycobacterium tuberculosis has evolved different strategies that modify its phagosome: first, a lipoarabinomannan phosphatidylinositol inhibits the acquisition of late endosomal/lysosomal molecules and second, a phosphatidylinositol mannoside mediates the fusion with early endosomal compartments (Chua et al. 2004). These mycobacterial lipid products act as trafficking toxins modulating the host cell membrane sorting and organelles biogenesis besides modulating the traffic of lysosomal constituents from the trans-Golgi network to the phagosome that harbors the mycobacterial (Chua et al. 2004). A protein kinase G actively secreted by pathogenic mycobacteria also inhibits phagolysosomal fusion and promotes bacterial survival within macrophages (Walburger et al. 2004).
In our study, the use of different methodologies allowed the accurate analysis of the endocytic compartments from uninfected and T. cruzi-infected cardiac cells. In situ data obtained by immunocytochemistry assays showed that the downmodulation of EEA1, Rab11, and Rab7 occurred in both nonparasitized as well as T. cruzi-parasitized host cells in the infected cultures, suggesting that soluble factors secreted or released from the parasite or from the infected cells could be involved in these alterations in a similar way that has been reported by Ochatt et al. (1997), which demonstrated that the cytosol of T. cruzi-infected macrophages impairs the fusion of endocytic compartments in in vitro assay.
The mechanisms and molecules implicated in the regulation of EEA1 and Rab proteins during the invasion of cardiomyocytes by T. cruzi are not known, but some hypothesis could be envisaged including (1) the participation of parasite (lipids or proteins) products, (2) the role of host cell (inflammatory mediators) molecules, or (3) a counterproductive consequence of activated cardiomyocytes exerting microbicidal activity or producing inflammatory mediators.
Indeed several alterations in the host cell's physiology caused by different molecules of the parasites have been reported. One of these molecules is the parasite trans-sialidase, a unique enzyme, which hydrolyses and transfers sialic acid residues from the host molecules to the T. cruzi epitopes (Frasch 2000). Besides playing a role during the parasite invasion (Schenkman and Eichinger 1993), it has been implicated in the neurite outgrowth and rescue of neuronal cell from apoptotic death caused by growth factor deprivation (Chuenkova and Pereira 2000), as well as triggering apoptosis in thymus, spleen, and lymph nodes during the acute infection (Mucci et al. 2002). Regarding altered endocytosis, a previous study suggested that T. cruzi display soluble factors that can inhibit the fusion of host endosomes (Ochatt et al. 1997).
Although molecules of the parasite are potential candidates for triggering the observed alterations in the cardiac GTPase expression and impairment of endocytosis, we cannot discard the possibility that mediators synthesized by their own cardiac infected cultures such as growth factors, cytokines, and chemokines could be responsible for the noted alterations. Actually, besides providing an ideal environment for parasite growth and spreading, cardiomyocytes are also considered active host cells because they can actively secrete inflammatory mediators and display trypanocidal activity mediated by nitric oxide (Machado et al. 2000). Literature data show that cytokines and chemokines can regulate Rabs expression (Stein et al. 2003). Stimulation of macrophages by IFN-γ, an inflammatory cytokine related to the intracellular resistance to microbes including T. cruzi (Fichera et al. 2004), results in the upregulation of Rab5a synthesis (Alvarez-Dominguez and Stahl 1998). Another interesting fact is the down-regulation of Rab11 by transforming growth factor β (Lindfors et al. 2001), which is a cytokine present in high levels during T. cruzi infection, being related to the genesis of fibrosis (Waghabi et al. 2002).
The differentiation of dendritic cells into antigen-presenting cells is accompanied by a dramatic and irreversible decrease in pinocytosis (Sallusto et al. 1995). Likewise, IFN-γ-and lipopolysaccharide-activated macrophages present diminished rates of fluid-phase endocytosis and phagocytosis besides presenting a delayed progression of incoming ligands to late endosomes and lysosomes (Tsang et al. 2000). As consequence of this slowed membrane trafficking, there is a prolonged exposure of the internalized molecules to the acidic nonlysosomal compartment, which may facilitate microbicidal chemistries or antigen processing (Tsang et al. 2000). So, another possibility is that the decreased pinocytosis of Dx-FITC by T. cruzi-infected cardiomyocytes resulted from the activation of the cardiac functions by their own synthesis of inflammatory mediators in vitro. Because cardiac cells have toxic microbicidal activity and are able to release inflammatory mediators (Machado et al. 2000), the slowing endocytosis could be a counterproductive consequence of the infected cardiomyocyte activation.
In summary, T. cruzi infection alters the expression of GTPase Rab proteins and impairs the endocytosis in mouse cardiomyocytes, which can contribute to the overall altered host cell physiology by modifying the normal incoming of nutrients and interfering with other important events related to the endocytic pathway.
Footnotes
Acknowledgements
Grants from Fundação Carlos Chagas Filho de Amparo a Pesquisa do Estado do Rio de Janeiro (FAPERJ), Conselho Nacional Desenvolvimento Científico e Tecnológico (CNPq), CAPES, and FIOCRUZ.
We are grateful to Dr. Andrea Henriques-Pons and Mirian Claudia Souza Pereira for critical suggestions (Instituto Oswaldo Cruz/FIOCRUZ). The authors thank Carlos Bizarro (Farmanguinhos/FIOCRUZ) for the confocal scanning microcopy analysis and Bruno Ávila (Instituto Oswaldo Cruz/FIOCRUZ) for technical support.