
Abstract
The comparative structure and expression of salivary components and vesicular transport proteins in the canine major salivary glands were investigated. Histochemical analysis revealed that the morphology of the five major salivary glands—parotid, submandibular, polystomatic sublingual, monostomatic sublingual, and zygomatic glands—was greatly diverse. Immunoblot analysis revealed that expression levels of α-amylase and antimicrobial proteins, such as lysozyme, lactoperoxidase, and lactoferrin, differed among the different glands. Similarly, Rab proteins (Rab3d, Rab11a, Rab11b, Rab27a, and Rab27b) and soluble N-ethylmaleimide-sensitive fusion protein attachment protein receptor (SNARE) proteins VAMP4, VAMP8, syntaxin-2, syntaxin-3, syntaxin-4, and syntaxin-6 were expressed at various levels in individual glands. mmunohistochemistry of Rab3d, Rab11b, Rab27b, VAMP4, VAMP8, syntaxin-4, and syntaxin-6 revealed their predominant expression in serous acinar cells, demilunes, and ductal cells. The VAMP4/syntaxin-6 SNARE complex, which is thought to be involved in the maturation of secretory granules in the Golgi field, was found more predominantly in the monostomatic sublingual gland than in the parotid gland. These results suggest that protein expression profiles in canine salivary glands differ among individual glands and reflect the properties of their specialized functions.
Introduction
The salivary glands generate and secrete saliva and are known to have an important role in digestion. Secretions from salivary glands wet the surface of the mouth and food, and play a role in facilitating mastication, swallowing, and vocalization. Salivary glands have also been implicated in the maintenance of the oral environment by secreting antimicrobial substances.1–3 In mammals, the structure and function of salivary glands are richly diverse. There are three major salivary glands in humans—parotid, submandibular, and sublingual—of which the submandibular glands account for 70%. 4 In carnivores, there are five major salivary glands—parotid, submandibular, polystomatic sublingual, monostomatic sublingual, and zygomatic. The polystomatic sublingual gland is located on the lateral side of the tongue and consists of multiple small glandular lobules that are integrated by connective tissues with multiple minor sublingual ducts. The monostomatic sublingual gland consists of a single lump with a major sublingual duct and is located on the cranial side of the submandibular gland. The zygomatic gland is a large, spherical, carnivore-specific gland originally derived from the dorsal buccal glands, which are minor salivary glands. The major salivary glands consist of an acinus and a well-developed duct system. An acinus is a cluster of secretory cells (serous, mucous, or a combination of the two) that connects to an intercalated duct to send saliva to a continuous striated duct. The arrangement and composition of the two types of acinar cells and ductal organization are diverse in each salivary gland of the same animal as well as within animal species, and even salivary glands of the same name.
The Rab family of proteins is composed of more than 60 members in mammals and regulates a specific step in intracellular vesicle trafficking.5,6 Rab proteins exhibit ubiquitous or tissue-specific expression, 7 and in exocrine glands such as the exocrine pancreas, Rab3d, Rab27a, and Rab27b have been shown to be localized on acinar zymogen granules and play a role in regulating exocytosis.8–11 In the salivary glands, the expression of several Rab proteins has been found not only in acinar cells but also in ductal cells. For example, Rab3d, Rab4, Rab26, Rab27b, and Rab33a are described in rat parotid acinar cells,12–19 whereas Rab11a and Rab11b are expressed in a human submandibular ductal cell line. 20
Soluble N-ethylmaleimide-sensitive fusion protein attachment protein receptor (SNARE) proteins function to mediate vesicle fusion by assembling synaptobrevin/vesicle-associated membrane protein 2 (VAMP2)/synaptobrevin-2 (v-SNARE; localized in vesicle membranes) and syntaxin-1 and SNAP-25 (t-SNARE; localized in target membranes) to form a trans-SNARE complex. 21 Mammalian SNARE proteins comprise three conserved families—VAMPs, syntaxins, and SNAP-25 homologues. In the original SNARE hypothesis describing the release of a neurotransmitter from the synaptic vesicles of neurons, v-SNARE forms a heterodimeric core complex with t-SNARE on the plasma membrane, and their interaction mediates membrane fusion to release the neurotransmitter. 22 In contrast to the small synaptic vesicles released by neurons, endocrine and exocrine cells release larger secretory granules that contain bioactive hormones or zymogens. Knowledge about the expression of SNARE proteins and the molecular mechanisms responsible for the regulated exocytosis in exocrine cells has been integrated. VAMP2 is expressed at the apical region around the acinar lumen of the exocrine pancreas 23 and the rat parotid gland, 24 and interacts with syntaxin-3 in the stimulated rabbit lacrimal gland 25 or with syntaxin-3, syntaxin-4, and SNAP-23 in the rat parotid gland. 26 In rat parotid acinar cells, intracellular localization of SNARE proteins has been demonstrated; syntaxin-2 and syntaxin-3 are localized in the apical plasma membrane, whereas syntaxin-4 is localized in the basolateral plasma membrane, and SNAP-23 is localized in the apical and intracellular plasma membranes. 27 In the human submandibular gland, SNAP-23, syntaxin-2, syntaxin-4, and VAMP2 were demonstrated to be localized at the luminal plasma membranes, and syntaxin-2 is colocalized with lactoferrin and α-amylase on the same secretory vesicles. 28 Furthermore, VAMP8/endobrevin plays an important role in regulating secretions of the pancreatic, lacrimal, and parotid acinar cells by forming a complex with syntaxin-4 and SNAP-23 in mice.29,30 It was also demonstrated that the interaction of SNAP-23 with VAMP2 was enhanced in response to an isoproterenol stimulus, but the interaction of SNAP-23 with VAMP8 or syntaxin-4 was scarcely augmented in the rat parotid gland. 26
Expression profiles of salivary secretory proteins and vesicular transport proteins in carnivorous salivary glands have not been investigated. Although Rab and SNARE proteins have been shown to be involved in regulating exocytosis in salivary glands, most of the findings are relatively limited to the tissue from the rodent parotid gland and the human submandibular gland, or epithelial cell lines of parotid or submandibular gland origin. So far, there has been no report on the expression of Rab and SNARE proteins in carnivorous salivary tissues. In this study, we attempted to elucidate the expression profiles of Rab, SNARE, and secretory proteins in the major salivary glands of the dog (Canis lupus familiaris). We further analyzed v- and t-SNARE protein complexes that were expressed in the canine tissue. Our results showed that expression levels of these proteins were not consistent in individual salivary tissues and that there was a difference in the expression pattern between different species, such as dog and rat.
Materials and Methods
Animals
Two male Beagle dogs (C. lupus familiaris) aged 9 to 10 months were purchased from Kitayama Labes Co., Ltd. (Nagano, Japan), and four Wistar male rats aged 8 weeks were purchased from CLEA Japan, Inc. (Tokyo, Japan). Following euthanasia by intravenous injection of 50 mg/kg body weight (bw) sodium pentobarbital and 2 mmol/kg/bw potassium chloride solution, the dogs were processed for sample preparation. The rats were deeply anesthetized by intraperitoneal injection of 0.1 mg/g/bw sodium pentobarbital and then processed for sample preparation. All animal experiments were performed in accordance with the Guidelines for the Care and Use of Laboratory Animals at the Institute of Experimental Animal Science, College of Bioresource Sciences, Nihon University, and the Guidelines for Animal Care, the Regulation of Infectious Agents, and Experimental Committee, Gunma University, Japan.
Primary Antibodies and Tissue Preparation
Antibodies against secretory proteins, the Rab family, and SNARE proteins used in this study are listed in Table 1. The canine major salivary glands (parotid, submandibular, polystomatic sublingual, monostomatic sublingual, and zygomatic) and rat parotid gland were fixed in Bouin’s fixative for general histological and immunohistochemical staining, or frozen on dry ice and stored at −80C until used for immunoblotting and coimmunoprecipitation.
Antibodies Used for the Analysis of Canine Salivary Glands.
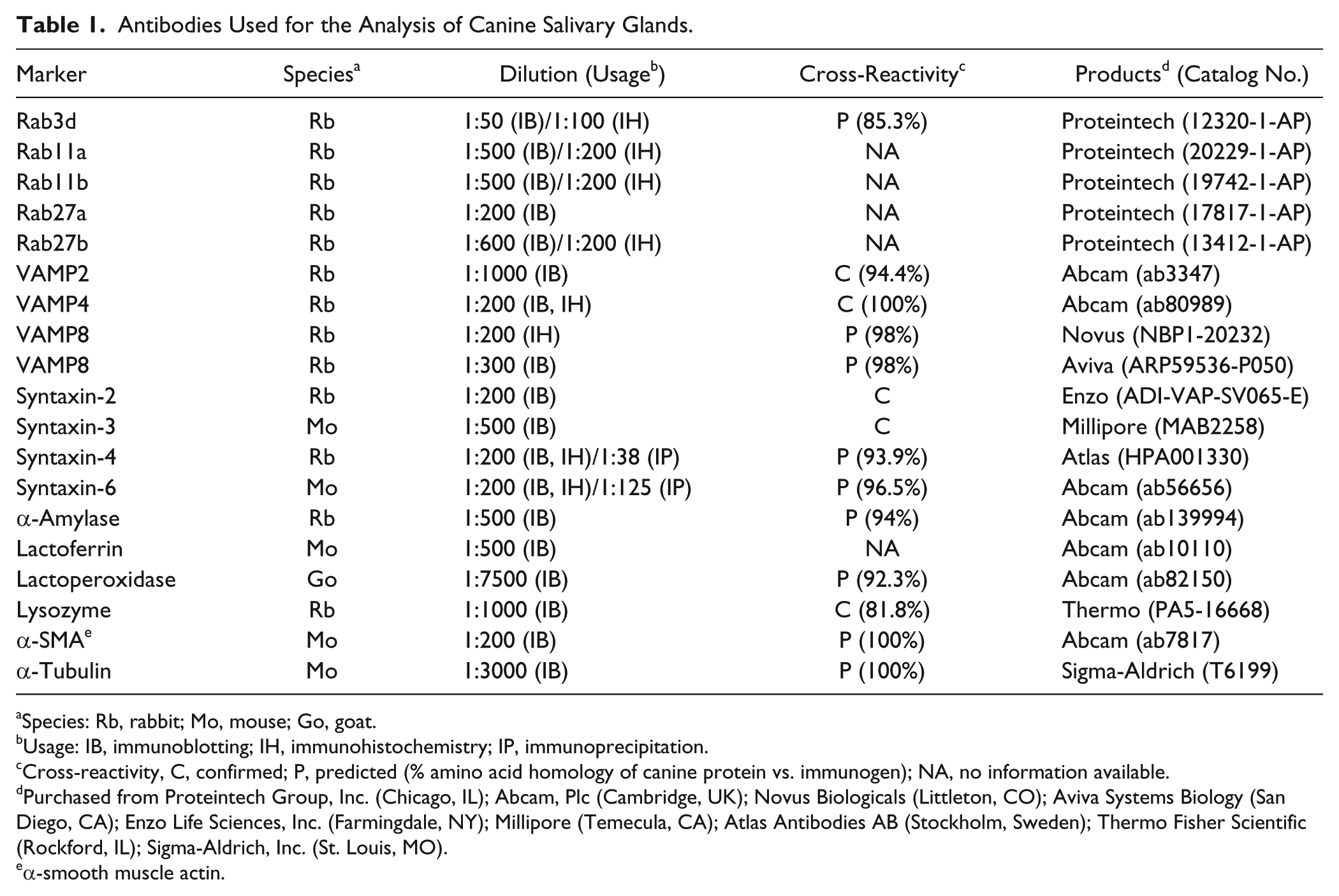
Species: Rb, rabbit; Mo, mouse; Go, goat.
Usage: IB, immunoblotting; IH, immunohistochemistry; IP, immunoprecipitation.
Cross-reactivity, C, confirmed; P, predicted (% amino acid homology of canine protein vs. immunogen); NA, no information available.
Purchased from Proteintech Group, Inc. (Chicago, IL); Abcam, Plc (Cambridge, UK); Novus Biologicals (Littleton, CO); Aviva Systems Biology (San Diego, CA); Enzo Life Sciences, Inc. (Farmingdale, NY); Millipore (Temecula, CA); Atlas Antibodies AB (Stockholm, Sweden); Thermo Fisher Scientific (Rockford, IL); Sigma-Aldrich, Inc. (St. Louis, MO).
α-smooth muscle actin.
General Histology
Fixed tissue blocks were dehydrated and embedded in paraffin. Tissue sections 5 µm–thick were mounted on silane-coated glass slides, and sections were deparaffinized in xylene and then rehydrated in a series of graded ethanol and water solutions. H&E stain was applied to observe the gross histological features of the tissues. Mallory–Azan (Azan) and alcian blue (pH 2.5)–periodic acid Schiff (AB-PAS) methods were used to segregate serous and mucous acinar cells. 31
Immunoblotting
Tissue samples were prepared for sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and immunoblotting was performed as described previously. 32 Rat tissue was used as a positive control. The protein concentrations of the tissue extracts were determined using a Coomassie Brilliant Blue color solution (Nacalai Tesque; Kyoto, Japan). Each protein extract was normalized for total protein content (25 or 50 µg for loading on polyacrylamide gel) and was used for SDS-PAGE and immunoblotting. Immobilon-P transfer membranes (Merck Millipore; Darmstadt, Germany) were used for immunoblot antigens with molecular mass >20,000 Da, and Immobilon-PSQ transfer membranes (Merck Millipore) were used for immunoblot antigens with molecular mass <20,000 Da. To analyze Rab and SNARE proteins, primary antibodies against Rab3d, Rab11a, Rab11b, Rab27a, Rab27b, VAMP1, VAMP2, VAMP3, VAMP4, VAMP7, VAMP8, syntaxin-1, syntaxin-2, syntaxin-3, syntaxin-4, and syntaxin-6 were used. For the analysis of secretory proteins, primary antibodies against α-amylase, lactoferrin, lactoperoxidase, and lysozyme were used. Antibodies against α-smooth muscle actin (α-SMA) were used to detect ductal and myoepithelial cell markers. Anti-α-tubulin antibody was used as the internal control. The peroxidase-conjugated secondary antibodies (diluted 1:5000) used were as follows: anti-rabbit IgG (catalog no. 111-035-144), anti-mouse IgG (115-035-146), and anti-goat IgG (705-035-147) (Jackson ImmunoResearch Laboratory; West Grove, PA).
Immunohistochemistry
For immunohistochemistry of Rab and SNARE proteins, we used paraffin-embedded sections. Antibodies (anti-Rab3d, -Rab11b, -Rab27b, -VAMP4, -VAMP8, -syntaxin-4, and -syntaxin-6), which are workable in paraffin-embedded sections, were chosen to analyze the intracellular localizations and polarities of target proteins, and their expressions were confirmed by immunoblotting. Deparaffinized sections were processed using microwave antigen retrieval as described previously. 33 The sections were treated with Dako REAL Peroxidase-Blocking Solution (Dako; Glostrup, Denmark) and blocked with 5% normal goat serum (Vector Laboratories; Burlingame, CA) in PBS before the antibodies were added. After incubating with the primary antibodies (diluted with PBS containing 0.01% Triton X-100) overnight at 4C, sections were incubated with secondary antibodies for 2 hr at room temperature using the horseradish peroxidase (HRP)–labeled polymer technique with the EnVision Dual Link System–HRP Kit (Dako) and then visualized using the SK-4100 peroxidase substrate kit (Vector Laboratories). As a negative control, sections were incubated with normal rabbit or mouse IgG (Santa Cruz Biotechnology, Inc.; Santa Cruz, CA; sc-2027 or sc-2025, 1 µg/ml) instead of with primary antibodies. Alternatively, sections were incubated with a mixture of diluted antibodies and absorbing immunogen peptides (2 µg/ml) for Rab3d, Rab27b, VAMP4, VAMP8, and syntaxin-4. Images were acquired using a BX51 microscope equipped with a DP72 CCD camera (Olympus; Tokyo, Japan) and digitized with DP2-BSW software (Olympus).
Immunoprecipitation
Coimmunoprecipitation analysis of the interaction between v- and t-SNARE proteins was conducted as described previously. 34 Salivary glands were homogenized in 20 mM Tris (pH 7.5) containing 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 2.5 mM MgCl2, 1% Triton X-100, and 1× protease inhibitor cocktail (cOmplete mini; Roche Diagnostics, Mannheim, Germany). The homogenates were kept on ice for 30 min and then centrifuged at 20,000 × g for 20 min. The collected supernatants were centrifuged again to remove fat. The protein concentrations of the collected supernatants were determined, and the soluble protein extracts (2 mg total protein/ml) were precleared by incubation with 100 µl of 50% slurry of protein G-Sepharose 4 Fast Flow (17061801; GE Healthcare UK Ltd., Buckinghamshire, England) for 3 hr at 4C under gentle rotation, and then centrifuged 7000 × g for 3 min. A small aliquot (50 µl) of the precleared lysate was removed and mixed with an equal volume of 2× SDS sample buffer to prepare the original total protein samples. Tissue lysates were incubated with 8 µg of antibodies against syntaxins (HPA001330 rabbit anti-syntaxin-4, Atlas Antibodies, Bromma, Sweden; and ab56656 mouse anti-syntaxin-6, Abcam, Plc) or 8 µg normal sc-2027 rabbit or sc-2025 mouse IgGs (Santa Cruz Biotechnology, Inc.) overnight at 4C. The immune complexes were captured by the adding of 50 µl of 50% slurry of protein G-Sepharose 4 Fast Flow for 2 to 3 hr under gentle rotation at 4C. The immuneprecipitates were washed four times with homo-genization buffer. One-fourth of the precipitated immunocomplexes were separated using SDS-PAGE (15% gel), and the interaction among SNARE proteins was analyzed using immunoblotting with ab80989 rabbit antibodies against VAMP4 (Abcam, Plc) or ARP59536-P050 rabbit antibodies against VAMP8 (Aviva Systems Biology).
Results
General Histology of Canine Salivary Glands
We first examined the structure of the secretory acini and ducts of canine salivary glands (Fig. 1) using general histological methods. H&E and Azan staining identified two types of secretory cells: serous and mucous acinar cells. In the control rat parotid gland, all acinar cells were serous cells (Fig. 2A–C). Similarly, in the canine parotid gland, nearly all acinar cells were serous cells, although the staining results were slightly different from those in the rat parotid gland (Fig. 2D–F). Mucous acinar cells were dominant in the submandibular and zygomatic glands, and the structure of serous demilunes was prominent (Fig. 2G and H, and insets in P and Q). The proportions of serous or mucous acini were different between the polystomatic sublingual and monostomatic sublingual glands. In the polystomatic sublingual gland, mucous acini predominated (Fig. 2J and K), whereas in the monostomatic sublingual gland, serous acini predominated (Fig. 2M and N). AB-PAS staining was strong in the mucous acinar cells (deep bluish magenta) and serous cells (magenta or light bluish magenta; Fig. 2F, I, L, and R). Coexistence of the mucous and serous acini constituting the clusters was observed in the polystomatic sublingual and zygomatic glands (Fig. 2J–L and P–R). Striated ducts were obvious in all glands, whereas intercalated ducts were well-developed in the parotid and submandibular glands (Fig. 2E, G, and H).
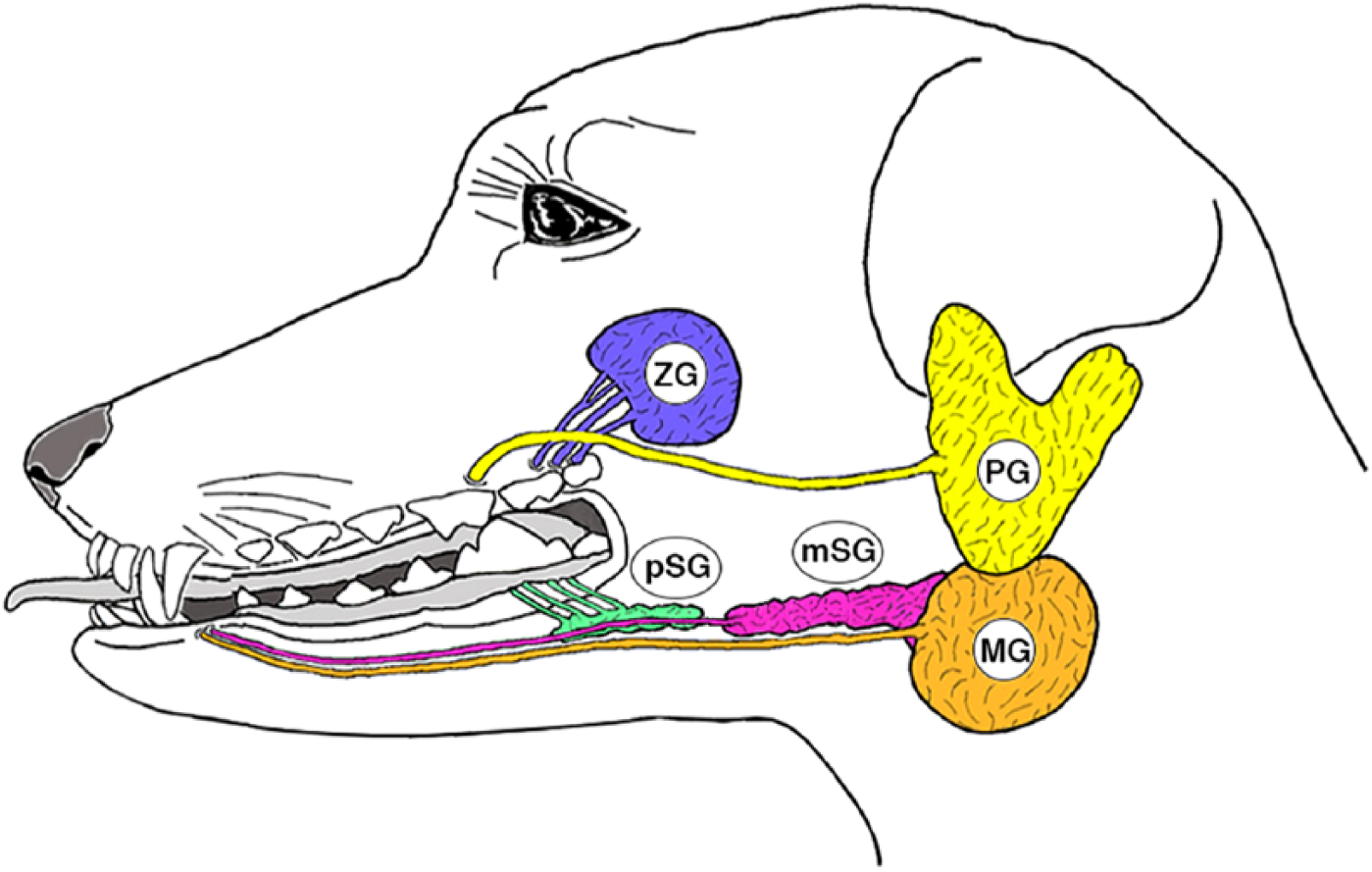
Canine major salivary glands. Abbreviations: PG, parotid gland; MG, submandibular gland; pSG, polystomatic sublingual gland; mSG, monostomatic sublingual gland; ZG, zygomatic gland.
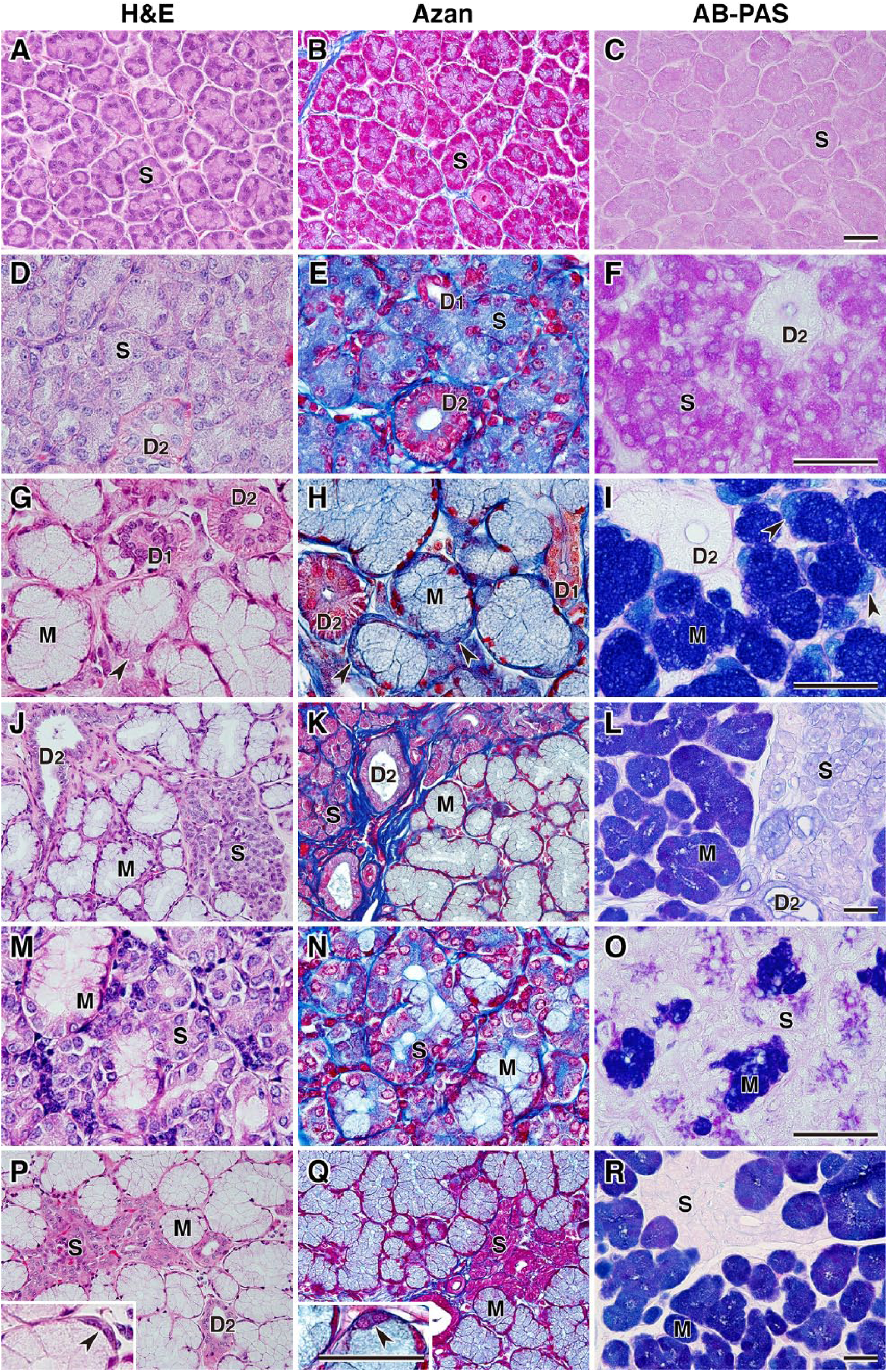
General histology of canine salivary glands and rat parotid gland. The rat parotid gland (A–C) and five canine salivary tissues—parotid (D–F), submandibular (G–I), polystomatic sublingual (J–L), monostomatic sublingual (M–O), and zygomatic glands (P–R)—were stained with H&E, Azan, and AB-PAS. Arrowheads in panels G–I indicate demilunes in the submandibular gland and the insets (in P and Q) indicate demilunes in the zygomatic gland. Scale, 50 µm. Abbreviations: AB-PAS, alcian blue (pH 2.5)–periodic acid Schiff; S, serous acinus; D1, intercalated duct; D2, striated duct; M, mucous acinus.
Expression of Secretory Proteins in Canine Salivary Glands
We examined the expression of secretory proteins by immunoblotting. Equal amounts of protein extracts (25 µg) from different salivary glands were analyzed using antibodies against α-amylase, lactoferrin, lactoperoxidase, and lysozyme (Fig. 3). In the rat parotid gland, presumed glycosylated (~67 kDa) and unglycosylated (~52 kDa) forms of α-amylase were observed, while the unglycosylated form was dominantly detected in the canine salivary glands and pancreas in both species. Expression levels of antimicrobial substances were different in the canine salivary glands. Lactoferrin was expressed in all salivary glands, but its level in the submandibular gland was very low. Although two bands with molecular masses of ~70 and 100 kDa lactoperoxidase were detected, either expression in the rat and canine parotid glands, or both expressions in the canine sublingual glands were observed. Expression levels of lactoperoxidase in the zygomatic gland were very low, while no expression could be detected in the submandibular gland. The expression profile of lactoperoxidase was consistent with the finding that two bands are detected in human saliva and whey. 35 Expression levels of lactoperoxidase in the rat parotid gland appeared higher than those in the canine salivary glands, presuming that the primary antibody reacts equally with both the rat and dog antigens. Lysozyme was strongly expressed in canine parotid and zygomatic glands. Lysozyme expression in the rat parotid gland has been reported immunohistochemically 36 ; however, the expression level of lysozyme could not be compared by immunoblotting because the primary antibody used in this study reacts with canine protein but not with rat protein. The expression level of α-SMA, a marker for myoepithelial cells, 37 was relatively strong in the canine tissues, suggesting the probable association between secretory function and muscular contraction.
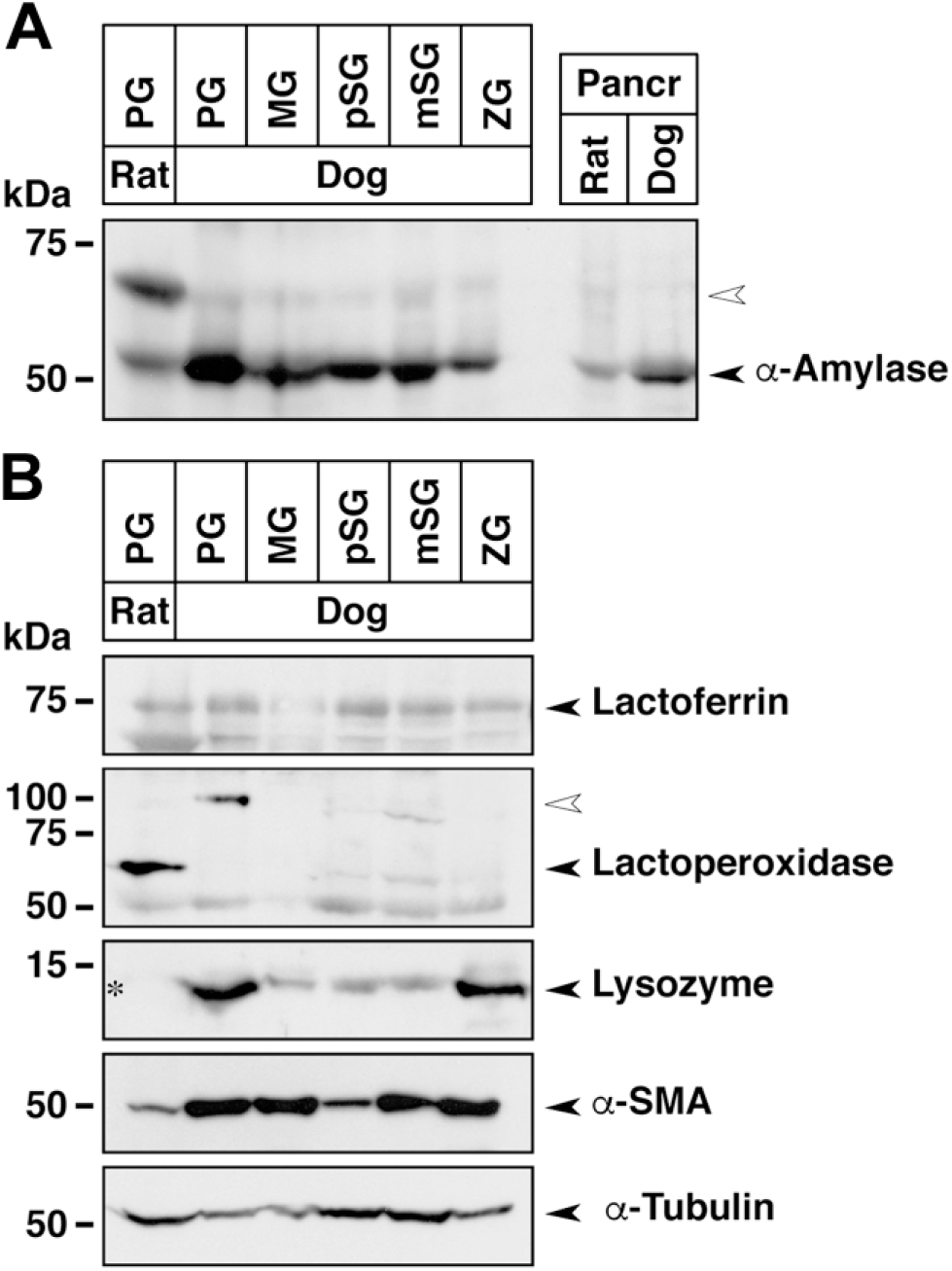
Expression of amylase, antibacterial substances, and structural marker proteins in canine salivary glands. Equal amounts of protein extracted from the indicated salivary glands (25 µg) and pancreas (10 µg) of rat and dog were used for SDS-PAGE, followed by immunoblotting. (A) Expression of α-amylase in salivary glands and pancreas. The open arrowhead in the large-sized bands indicates the presumed glycosylated form. (B) Expression of antibacterial substances (lactoferrin, lactoperoxidase, and lysozyme) and marker proteins. The open arrowhead in the lactoperoxidase panel indicates the presumed glycosylated form. Asterisk indicates no cross-reactivity of antilysozyme antibody against rat protein. The bottom two panels show the control α-SMA and α-tubulin immunoblots. Abbreviations: SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis; α-SMA, α-smooth muscle actin; PG, parotid gland; MG, submandibular gland; pSG, polystomatic sublingual gland; mSG, monostomatic sublingual gland; ZG, zygomatic gland; kDa, kilodalton.
Expression of Rab Proteins in Canine Salivary Glands
We examined the expression of Rab proteins in canine salivary glands using immunoblotting and immunohistochemistry. Protein extracts (25 µg) from rat and canine tissues were analyzed by immunoblotting using antibodies against Rab3d, Rab11a, Rab11b, Rab27a, and Rab27b (Fig. 4A). Among the four isoforms of Rab3, only Rab3d was detected in the tissue preparations. The expression levels of the Rab11 and Rab27 subfamilies varied in the canine salivary glands.
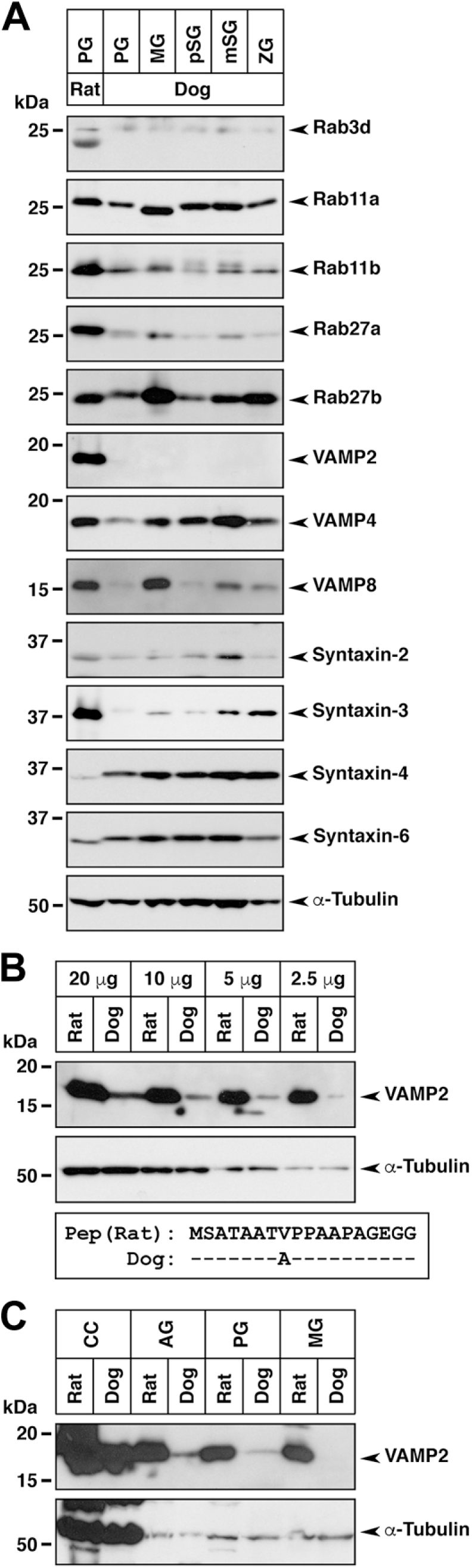
Expression of Rab and SNARE proteins in canine salivary glands. (A) Equal amounts of protein extract (25 µg) from the indicated salivary glands of the rat and dog were used for SDS-PAGE, followed by immunoblotting using antibodies against Rab, t-SNARE, and v-SNARE proteins. The bottom panel shows the control α-tubulin immunoblots. (B) Detection of VAMP2 in diluted brain (cerebral cortex) extract from the rat and dog. The bottom panel shows the peptide alignment of immunogen (Pep) from rat VAMP2 (amino acids 1–18) and corresponding dog VAMP2 amino acids. The identical amino acids are indicated by dashes. (C) Expression of VAMP2 in the cerebral cortex, adrenal glands, and salivary glands from the rat and dog (50 µg protein extract). Abbreviations: SNARE, soluble N-ethylmaleimide-sensitive fusion protein attachment protein receptor; SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis; VAMP, vesicle-associated membrane protein; PG, parotid gland; MG, submandibular gland; pSG, polystomatic sublingual gland; mSG, monostomatic sublingual gland; ZG, zygomatic gland; CC, cerebral cortex; AG, adrenal gland; kDa, kilodalton.
Immunohistochemical analysis of Rab3d, Rab11b, and Rab27b revealed that these Rab proteins were localized in the serous acinar cells or demilunes, ductal cells, and interacinar plasma cells (Fig. 5). The expression levels of Rab3d and Rab11b in the serous acinar cells of the polystomatic sublingual, monostomatic sublingual, and zygomatic glands were higher than those in the cells of the parotid and submandibular glands (Fig. 5A, B, D, E, G, H, J, K, M, N); however, similar expression levels of Rab27b in the serous acinar cells or demilunes in each salivary gland were observed (Fig. 5C, F, I, L, O). The sections in the negative control did not show significant immunoreactivity (data not shown).
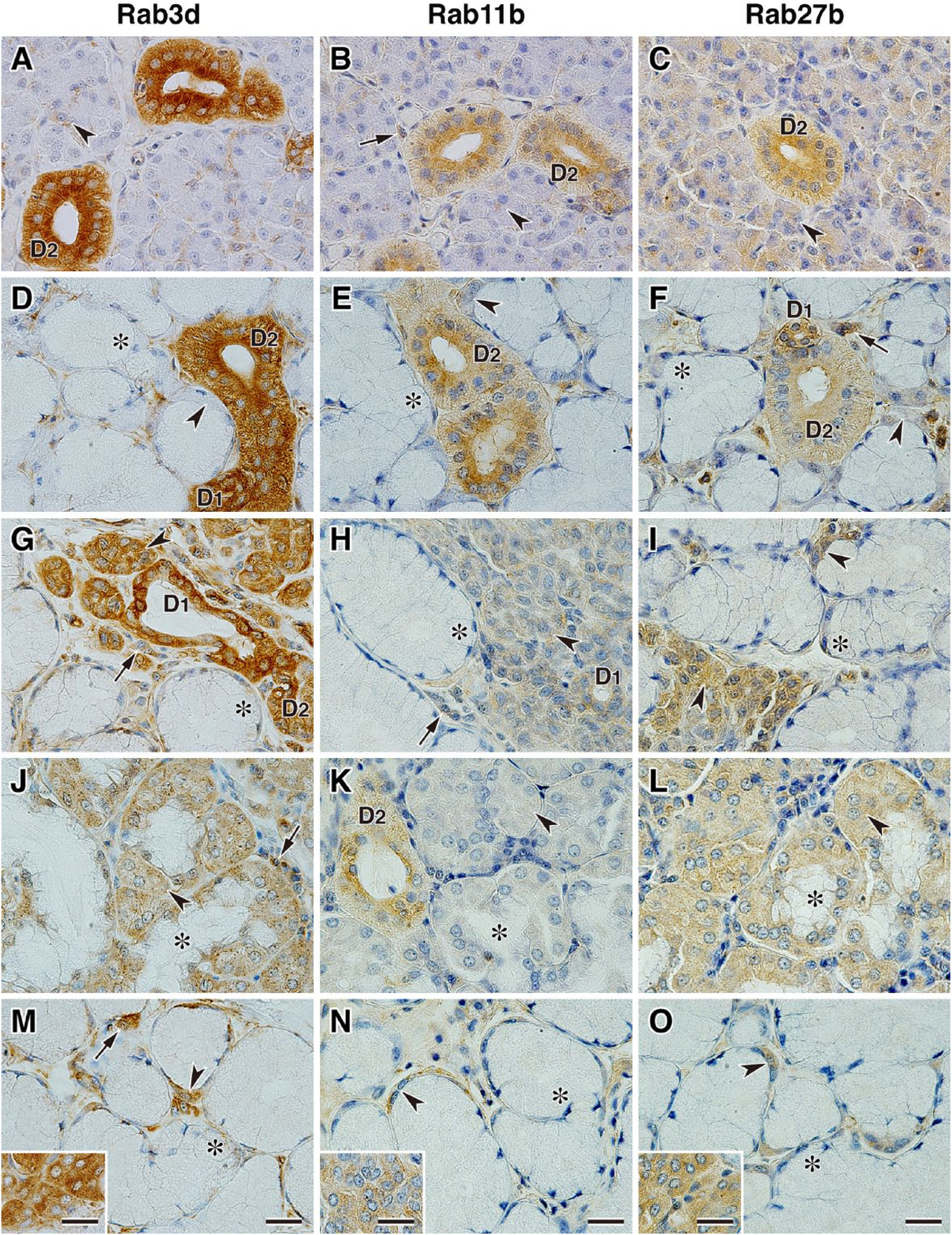
Localization of Rab3d, Rab11b, and Rab27b in canine salivary glands. Sections of parotid gland (A–C), submandibular gland (D–F), polystomatic sublingual gland (G–I), monostomatic sublingual gland (J–L), and zygomatic gland (M–O) immunostained with antibodies against Rab3d, Rab11b, and Rab27b. The inset panels in M, N, and O show the minor serous acinar cells coexisting with the major mucous acini in the zygomatic glands. Arrowheads indicate serous acinar cells or demilunes, and asterisks indicate mucous acinar cells. Arrows indicate interacinar plasma cells. Sections were counterstained with hematoxylin. Scale, 20 µm. Abbreviations: D1, intercalated duct; D2, striated duct.
Expression of SNARE Proteins in Canine Salivary Glands
We examined the expression of v-SNAREs (VAMP1, VAMP2, VAMP3, VAMP4, VAMP7, and VAMP8) and t-SNAREs (syntaxin-1, syntaxin-2, syntaxin-3, syntaxin-4, and syntaxin-6) by immunoblotting. Significant signals for VAMP2, VAMP4, VAMP8, syntaxin-2, syntaxin-3, syntaxin-4, and syntaxin-6 were detected (Fig. 4A). In the rat parotid gland, the expression of VAMP2, VAMP4, and VAMP8 was detected, while the expression of VAMP2 in the canine salivary glands could not be detected. Previous reports demonstrated that VAMP2 immunoreactivity was detected not only in the rat parotid gland24,26 but also in the human submandibular gland. 28 The canine protein cross-reactivity with the anti-VAMP2 antibody (ab3347) used in our study was ensured for canine tissues, notwithstanding that there is one amino acid difference in the middle portion of VAMP2 immunogenic peptides (18 amino acids); therefore, we checked the cross-reactivity with canine brain, which is known as a VAMP2 high-expressing tissue, and detected a significant signal (Fig. 4B). We then examined VAMP2 expressions in several tissues (parotid and submandibular glands, as well as the brain and adrenal glands) with an overloading condition, and eventually we barely observed the weak immunosignals in the canine salivary glands (Fig. 4C). Thus, it was apparent that the cross-reactivity between the anti-VAMP2 antibody and the canine protein was very low and the expression levels of VAMP2 in the salivary glands was much lower than that in the brain (~1/10 to 1/20). In addition, expression levels of VAMP8 were low in the canine parotid and polystomatic sublingual glands compared with those in other glands.
Immunohistochemical analysis of VAMP4 and VAMP8 revealed that these v-SNAREs were predominantly localized in serous acinar cells, demilunes, ductal cells, and interacinar plasma cells (Fig. 6); however, in the submandibular gland, expression levels of VAMP4 and VAMP8 in demilunes were very weak compared with those in the ductal cells (Fig. 6C and D). Furthermore, VAMP4 but not VAMP8 was expressed in myoepithelial cells (Fig. 6A, C, E, G, I). Immunohistochemical analysis of syntaxin-4 and syntaxin-6 revealed that these t-SNAREs were expressed predominantly in serous acinar cells or demilunes and in ductal cells and showed a punctate staining pattern (Fig. 7). The intracellular localization of syntaxin-4 in serous and ductal cells showed uniform distribution (Fig. 7A, C, E, G, I), whereas the localization of syntaxin-6 in ductal cells showed a strong reaction on the luminal side (Fig. 7B, D, F).
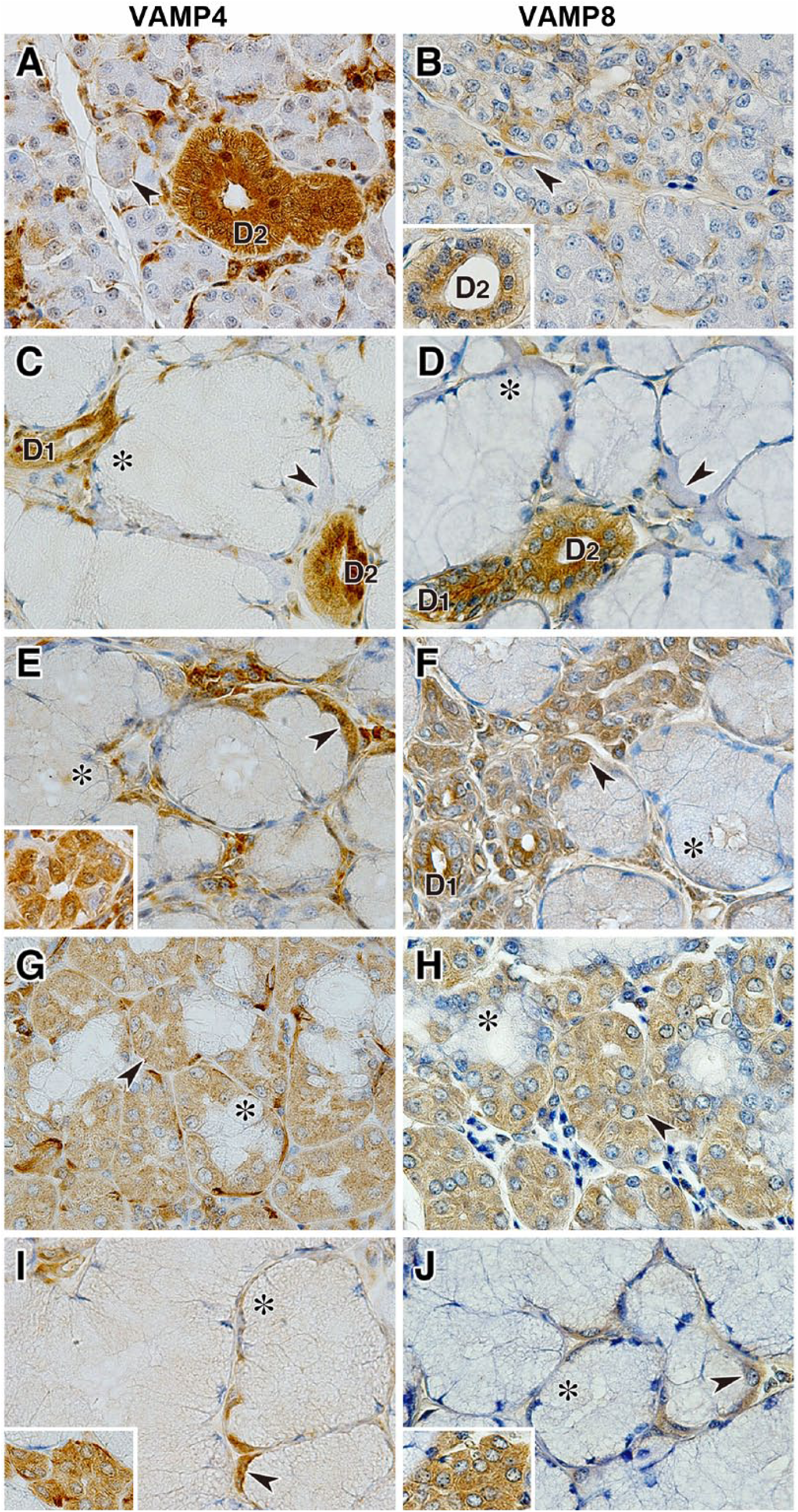
Localization of VAMP4 and VAMP8 in canine salivary glands. Sections of the parotid gland (A and B), submandibular gland (C and D), polystomatic sublingual gland (E and F), monostomatic sublingual gland (G and H), and zygomatic gland (I and J) immunostained with antibodies against VAMP4 and VAMP8. The inset panel in B shows the striated duct, and the inset panels in E, I, and J show the minor serous acinar cells coexisting with the major mucous acini in each gland. Arrowheads indicate serous acinar cells or demilunes, and asterisks indicate mucous acinar cells. The sections were counterstained with hematoxylin. Scale, 20 µm. Abbreviations: VAMP, vesicle-associated membrane protein; D1, intercalated duct; D2, striated duct.
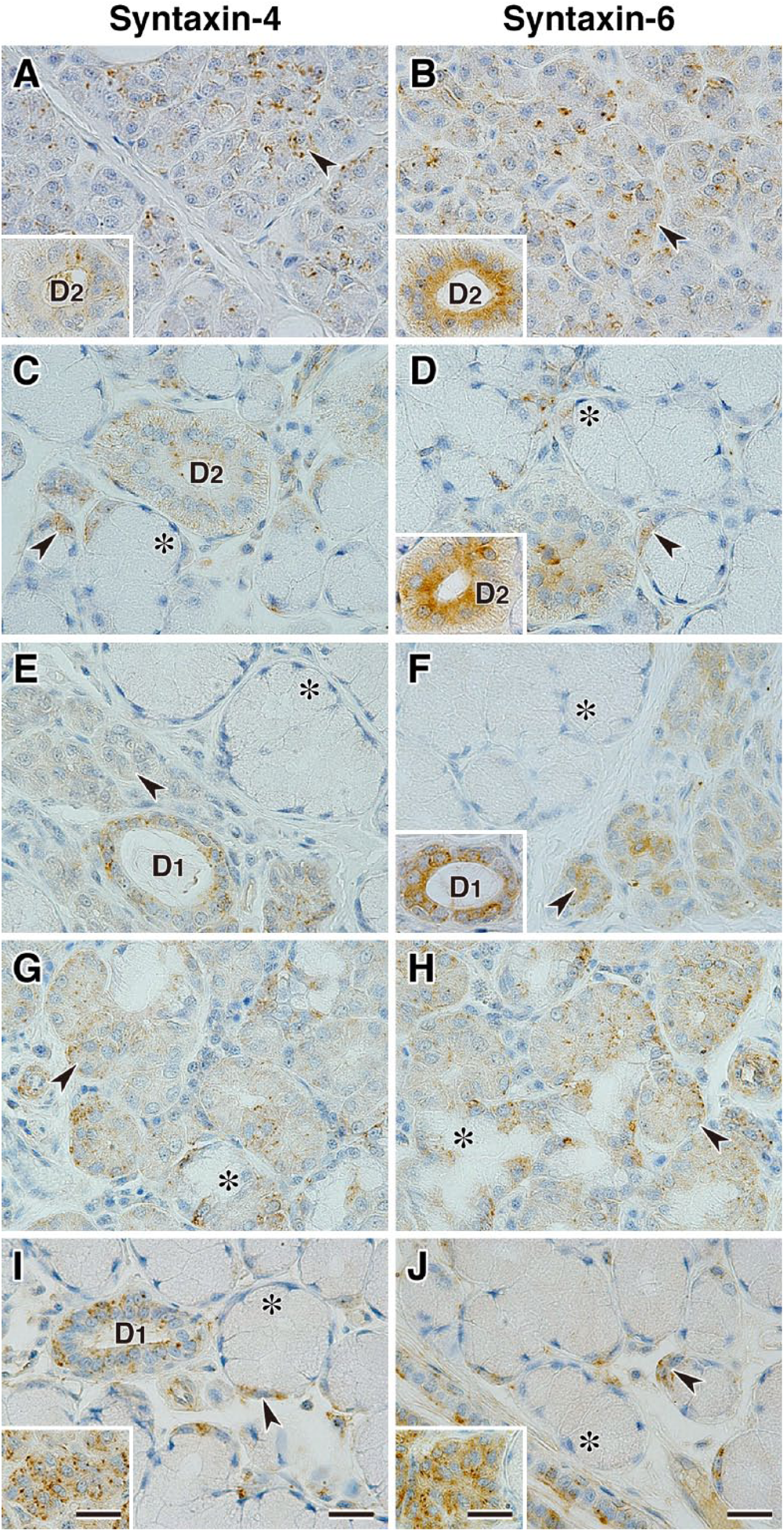
Localization of syntaxin-4 and syntaxin-6 in canine salivary glands. Sections of parotid gland (A and B), submandibular gland (C and D), polystomatic sublingual gland (E and F), monostomatic sublingual gland (G and H), and zygomatic gland (I and J) immunostained with antibodies against syntaxin-4 and syntaxin-6. The inset panels in A, B, and D show the striated duct, the inset panel in F shows the intercalated duct, and the inset panels in I and J show the minor serous acinar cells coexisting with the major mucous acini in each gland. Arrowheads indicate serous acinar cells or demilunes, and asterisks indicate mucous acinar cells. The sections were counterstained with hematoxylin. Scale, 20 µm. Abbreviations: D1, intercalated duct; D2, striated duct.
Protein Interaction of v- and t-SNAREs in Canine Salivary Glands
We observed high expressions of v-SNAREs (VAMP4 and VAMP8) and t-SNAREs (syntaxin-4 and syntaxin-6) in the canine salivary glands after immunoblotting and immunohistochemical analyses; therefore, we examined the interaction of v- and t-SNAREs by coimmunoprecipitation. The immunoprecipitates using antibodies against syntaxin-4 and syntaxin-6 were analyzed by immunoblotting using antibodies against VAMP4 or VAMP8. As shown in Fig. 8, anti-syntaxin-4 and anti-syntaxin-6 antibodies clearly immunoprecipitated their own antigens. Briefly, a clear interaction of syntaxin-6 with VAMP4 was detected in the rat parotid gland and canine monostomatic sublingual gland, but was barely detectable in the canine parotid gland under these conditions (Fig. 8B). No significant interactions between syntaxin-4 and VAMP4 or VAMP8 (Fig. 8A) and between syntaxin-6 and VAMP8 (Fig. 8B) were detected. Thus, the amount of the interactive protein complex formed between syntaxin-6 and VAMP4 in canine salivary glands might depend on the basal expression level of VAMP4.
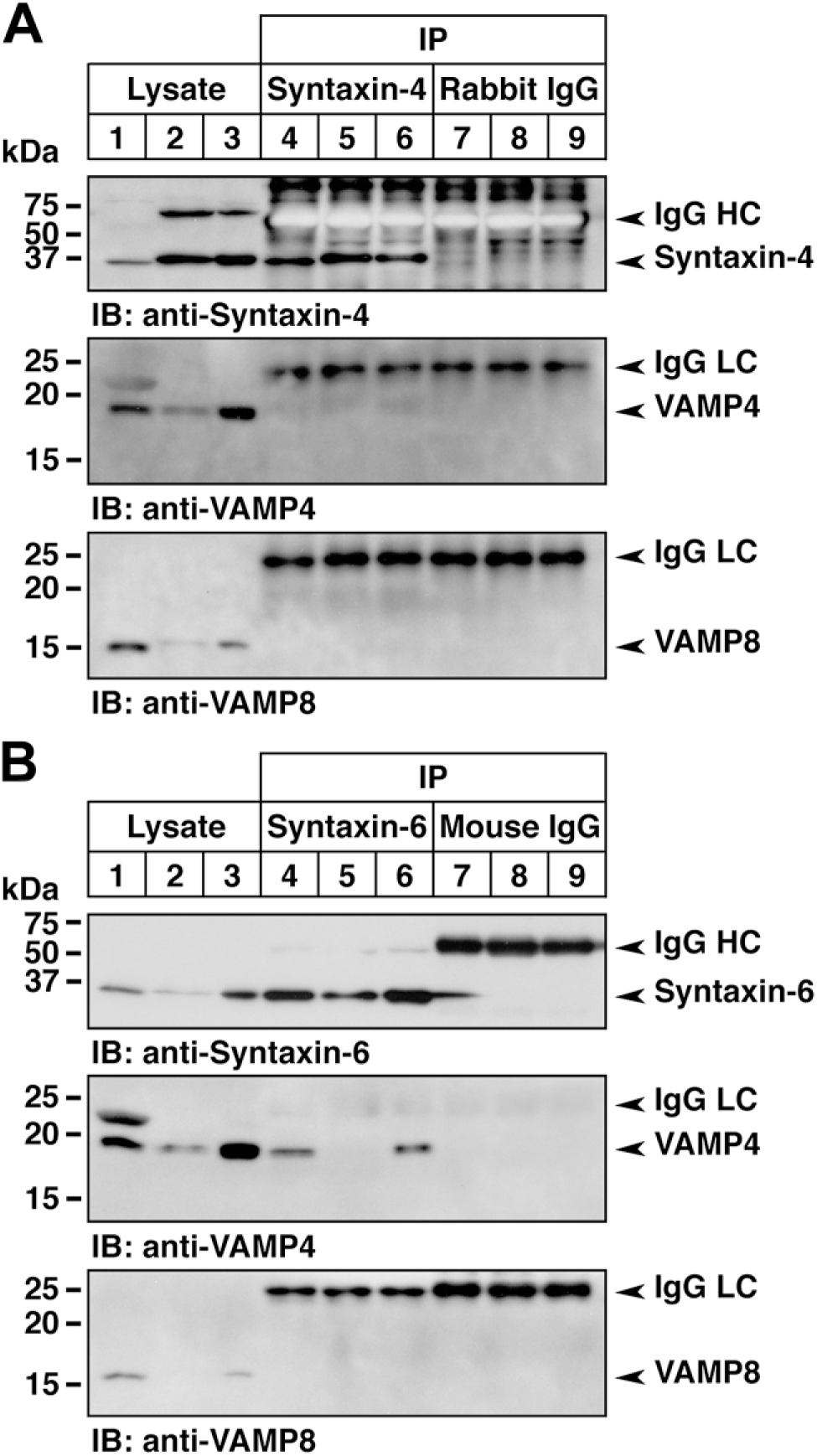
Interaction of v- and t-SNARE proteins in canine salivary glands. (A) Coimmunoprecipitation analysis of syntaxin-4 with VAMP4 or VAMP8. Tissue lysates from the rat (lanes 1, 4, and 7; parotid gland) and canine (lanes 2, 5, and 8, parotid gland; lanes 3, 6, and 9, monostomatic sublingual gland) salivary glands were immunoprecipitated using anti-syntaxin-4 antibody (lanes 4–6) and control normal rabbit IgG (lanes 7–9). The starting tissue lysates (25 µg, lanes 1–3) and immunoprecipitates were analyzed by immunoblotting with antibodies against syntaxin-4 (upper panel), VAMP4 (middle panel), and VAMP8 (lower panel). (B) Coimmunoprecipitation analysis of syntaxin-6 with VAMP4 or VAMP8. The same procedure in (A) was carried out using anti-syntaxin-6 antibody and control normal mouse IgG. Tissue lysates and immunoprecipitates were analyzed by immunoblotting with antibodies against syntaxin-6, VAMP4, and VAMP8. Abbreviation: VAMP, vesicle-associated membrane protein; HC, heavy chain; LC, light chain.
Discussion
The salivary glands produce and secrete several components, such as sugar digestive enzymes and antimicrobial substances. In mammals, their diversity in structure and function is rich, depending on feeding habits.38,39 In this study, we used the Beagle dog as a carnivore model and characterized its five major salivary glands. We first examined the expression of the secretory proteins in saliva and the vesicular transport proteins using immunoblot analysis, and then immunohistochemically examined Rab and SNARE proteins that were expressed predominantly in the salivary glands. We finally examined the in vivo properties of SNARE protein interactions by coimmunoprecipitation.
In canine salivary glands, the microscopic structure of the five major glands was prominently featured. There were various proportions and distributions of serous and mucous acinar cells. As is common in many mammalian species, the acinar cells in the canine parotid gland consist mostly of serous cells that contain PAS-positive substances. In the submandibular, polystomatic sublingual, and zygomatic glands, the proportion of AB-PAS double-positive mucous acini was high, and serous cells showed typical demilunes; however, in the polystomatic sublingual and zygomatic glands, medium- or small-sized serous acinar clusters coexisted in the predominant mucous acinar area. In contrast, the monostomatic sublingual gland was composed of predominantly serous and minor mucous acini. Furthermore, the staining patterns of serous acini in the parotid, polystomatic sublingual, monostomatic sublingual, and zygomatic glands using the Azan and AB-PAS methods were different. These results suggested that there are divergent properties in the expression of salivary components and vesicular transport proteins in the canine major salivary glands.
In this study, a comparative immunoblotting of secretory proteins revealed that α-amylase was expressed in the canine salivary glands. It has long been believed that salivary gland amylase is not expressed in carnivores. The report by Mocharla et al. 40 demonstrated that α-amylase gene expression was not detectable in the canine parotid gland by northern hybridization or reverse transcription–polymerase chain reaction (RT-PCR) analysis, despite slightly detected enzyme activity; therefore, we have an issue with α-amylase expression of canine salivary glands. One possible interpretation of no detection of the α-amylase RNA transcript is caused by mismatch of primers (four bases are different between the human and dog AMY gene in the P1 primer sequence) used in RT-PCR. Our immunoblotting using antisalivary α-amylase antibody detected two-sized bands (large, ~67 kDa; small, ~52 kDa) in the rat parotid gland and canine salivary glands. A small-sized band was also detected in both the rat and canine pancreas; therefore, it should be interpreted that α-amylase only slightly, but surely, expresses in canine salivary glands.
The expression and/or secretion of salivary components, which include antimicrobial substances, such as lysozyme, lactoperoxidase, lactoferrin, immunoglobulin A, and β-defensins, and glycoconjugates, such as mucin glycoproteins, as well as digestive enzymes such as α-amylase and lingual lipase,41,42 have been substantially demonstrated in the salivary glands of humans,3,43–48 rodents,36,49,50 and goats.51–53 Meanwhile, the expression of antimicrobial substances differed among glands, with high expressions of lysozyme in the parotid and zygomatic glands and lactoperoxidase in the parotid gland. Thus, our results emphasize that canine major salivary glands are functionally subdivided, such as for food digestion or the maintenance of the intraoral environment.
We confirmed the apparent expression of Rab proteins (Rab3d, Rab11a, Rab11b, Rab27a, and Rab27b) in canine salivary glands using immunoblot analysis. Interestingly, the comparative expression patterns of Rab11a versus Rab11b and Rab27a versus Rab27b differed between the dog and the control, suggesting a species-dependent difference in protein expression levels or salivary tissue architectures. In addition, immunohistochemical analysis of Rab3d, Rab11b, and Rab27b revealed that these proteins were predominantly expressed in serous acini/cells and ductal cells. This observation is consistent with the evidence showing that Rab3d, Rab27a, and Rab27b localize on the zymogen granules of rat parotid acinar cells and are involved in regulating amylase secretion.12,13,17,54,55 Rab11b is expressed in the ductal cells originating from the human submandibular gland and regulates acidosis-induced intracellular traffic of V-ATPase. 20 By analyzing genetically manipulated transgenic or knock-in mice, the expression of the Rab27a or Rab27b reporter gene was observed in the submandibular serous and mucous cells, which contain zymogen and mucigen granules.56,57 Furthermore, in human salivary glands, it was shown that Rab27b is expressed in serous acini and striated ducts. 58
We further confirmed the expression of SNARE proteins (VAMP4, VAMP8, syntaxin-2, syntaxin-3, syntaxin-4, and syntaxin-6) in canine salivary glands by immunoblotting. The immunohistochemistry of VAMP4, VAMP8, syntaxin-4, and syntaxin-6 revealed that these SNARE proteins were predominantly expressed in serous acinar cells, demilunes, and ductal cells. The immunoreactivity of Rab and SNARE proteins in mucous acinar cells appeared to be negative or very weakly positive. It is believed that the immunoreactivity of these proteins is most likely inhibited by intense glycoproteins, such as mucin in the mucous acinar cells identified by histochemical analysis.
It has been shown that syntaxin-6 is involved in clathrin-coated vesicle trafficking from the trans-Golgi network (TGN) to endosomes 59 and is localized in the limiting membrane of immature granules located in the Golgi region of rat pancreatic β-cells and parotid acinar cells, suggesting its role in secretory granule maturation in endocrine and exocrine cells. 60 It has also been shown that VAMP4 is localized in the Golgi stack, TGN membranes, clathrin-coated and noncoated vesicles, endosomes, and immature secretory granules, and it was found to colocalize with syntaxin-6 to form a protein complex in PC12 cells, suggesting that the interaction between VAMP4 and syntaxin-6 is involved in TGN-endosomal transport, including the formation of mature secretory granules. 61 This protein interaction is also implicated in post-Golgi early/recycling endosome-to-TGN retrograde transport. 62 More importantly, the formation of the VAMP4/syntaxin-6 complex is suggested to be linked to the degree of granule maturation and is involved in the transport of immature granules to the early endosomes in rat parotid acinar cells.63–65 It was recently reported that siRNA knockdown of VAMP4 and its cognate partners, syntaxin-6, syntaxin-16, and Vti1a, in HeLa cells led to a unique phenotype, showing a disrupting fragmentation of the Golgi ribbon structure in the juxtanuclear area. 66 Our coimmunoprecipitation results showed that VAMP4 sufficiently coprecipitated with syntaxin-6 principally in the rat parotid and canine monostomatic sublingual glands, but was barely detectable in the canine parotid gland. This result suggests that the degree of VAMP4/syntaxin-6 complex formation should be different depending on the salivary gland tissues and animal species.
It was reported that several SNARE protein complexes in the rat parotid gland are involved in the exocytosis of amylase granules.26,27,67 The syntaxin-4/SNAP-23 complex in the parotid gland is suggested to be compatible with the syntaxin-1/SNAP-25 complex in neural-endocrine tissues. 67 Furthermore, Imai et al. 27 reported interactions among syntaxin-2, syntaxin-3, and syntaxin-4 with SNAP-23 and VAMP8 using a yeast two-hybrid assay, and of syntaxin-3 with VAMP8 and syntaxin-4 with SNAP-23 in the apical plasma membrane preparation using immunoprecipitation. Interestingly, a stimulus-dependent increase in the transient interaction between SNAP-23 and VAMP2, but not VAMP8 or syntaxin-4, was found in the rat parotid gland treated with isoproterenol and/or cytochalasin D, and modified cAMP-mediated exocytosis. 26 In our study, the expression levels of syntaxin-2 and syntaxin-3 in the canine salivary glands were different among the tissues compared with those of syntaxin-4 and syntaxin-6. Using immunoprecipitation, we could not detect any significant interactions between syntaxin-4 and VAMP4 or syntaxin-4 and VAMP8 under our experimental conditions. Examination under additional experimental conditions might be necessary, and would be conducted by adjusting the sample preparations. The expression of VAMP2 was barely detected in our parotid tissue preparation, which did not include any enrichment for target vesicular or organellar membranes, as has been demonstrated in several reports.24,27,63,64 Furthermore, it is critical to be aware of the cross-reactivity between the canine tissues and the anti-VAMP2 antibody used in this study because these factors affect detection sensitivity. Because VAMP2 localizes on the secretory granule membrane and is involved in cAMP-induced amylase secretion through protein kinase A–dependent dissociation from its binding protein in the rat parotid gland,24,68 similar properties can most likely be explained in the canine salivary glands that express amylase protein.
In summary, we performed comparative histochemical and biochemical expression analyses of salivary components and vesicular transport proteins in the canine major salivary glands. This study is the first to describe the unique profiles of the canine salivary glands, which are characterized by divergent tissue structure, different expression levels of these proteins, and the species-specific differences in carnivorous animals.
Footnotes
Acknowledgements
We thank Dr. Chisato Kubota and Mrs. Mari Hosoi (Gunma University, Maebashi, Japan) for their technical support.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
HG designed, carried out the histological and biochemical experiments, data analysis, and drafted the manuscript. HO and RU performed immunohistochemistry for Rab and VAMP proteins. TY and AT carried out the canine tissue sampling and general histochemical analysis. ST and MH provided rat samples and contributed to the experimental design, data analysis, and reviewing manuscript. All authors have read and approved the final manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Grants-in-Aid from the Japan Society for the Promotion of Science (JSPS) #16K08078 (H.G.), #24390050 (S.T.), and #25440052 (M.H.). It was also supported in part by the joint research program of the Institute for Molecular and Cellular Regulation, Gunma University (no. 15012, H.G. and S.T., and no. 16010, M.H. and S.T.).