
Abstract
In kidney epithelial cells, a variety of physiological processes are dependent on the active recycling of membrane proteins between intracellular vesicles and the cell surface. Although clathrin-mediated endocytosis occurs in several renal cell types, endocytosis can also occur by non-clathrin-coated vesicles, including pinocytotic structures known as caveolae that contain a novel coat protein, caveolin. Exo- and endocytosis of a vacuolar H+-ATPase in intercalated cells also occurs via specialized “coated” vesicles that do not contain clathrin. The aim of this study was to localize caveolin in the kidney and, in addition, to determine whether it could be a component of the H+-ATPase recycling process. Using an antibody against the α- and β-isoforms of caveolin-1, our immunocytochemical data show a marked heterogeneity in the cellular expression of this isoform of caveolin in kidney. In contrast, caveolin-3 was not detectable in renal epithelial cells. Caveolin-1 was abundant in endothelial cells and smooth muscle cells and was present in the parietal cells of Bowman's capsule. Distal tubule cells, connecting tubule cells, and collecting duct principal cells exhibited marked punctate basolateral staining, corresponding to the presence of caveolae detected by electron microscopy, whereas all intercalated cells were negative in both cortex and medulla. These data indicate that although caveolin-1 may participate in basolateral events in some kidney epithelial cell types, it does not appear to be involved in the regulated recycling of H+-ATPase in intercalated cells. Therefore, these cells recycle H+-ATPase by a mechanism that involves neither clathrin nor caveolin-1.
R
However, endocytosis can also occur via other pathways, and considerable attention has been focused on caveolae as mediators of an alternative route of internalization that does not involve clathrin (Van Deurs et al. 1993). These pinocytotic invaginations of the plasma membrane are abundant in smooth muscle cells and endothelial cells but are also present in epithelial cells in many tissues (Lisanti et al. 1995; Parton 1996). Caveolae are decorated on their cytosolic surface with a protein called VIP21 or caveolin, a transmembrane cholesterol binding protein (Murata et al 1995; Li et al. 1996) that is homologous to a v-src tyrosine kinase substrate (Glenney 1992; Rothberg et al. 1992). Although the precise role of caveolae is uncertain, glycosyl phosphatidylinositol (GPI)-anchored membrane proteins, as well as many other proteins, were reported to concentrate in these structures (Anderson 1993; Chang et al. 1994; Lisanti et al. 1994). Some reports consider this to be an experimental artifact, however (Mayor and Maxfield 1995). Nevertheless, many membranes are decorated with caveolae, which may represent an alternative mechanism of internalization to clathrin-mediated endocytosis in some cell types. In addition, the association of GTP binding proteins with caveolae indicates a role for these structures in cellular signaling processes (Sargiacomo et al. 1993; Chun et al 1994; Lisanti et al. 1994,1995; Li et al 1995). Although the capacity of caveolae to detach from the plasma membrane and undergo endocytosis has been a matter of considerable debate, recent evidence has shown that caveolae can indeed function as endocytotic organelles (Kurzchalia and Parton 1996; Schnitzer et al. 1996).
We previously showed that vigorous endocytosis of the vacuolar H+-ATPase and fluid-phase markers occurs in kidney collecting duct intercalated cells via a clathrin-independent mechanism (Brown and Orci 1986; Brown et al. 1987a,b; Brown and Breton 1996). The vesicles that are involved in this process have an extensive cytoplasmic coat that contains cytoplasmically-oriented subunits of the vacuolar H+-ATPase (Brown et al. 1987a), but no detectable clathrin (Brown and Orci 1986; Brown et al. 1987b). Therefore, it is likely that endocytotic mechanisms other than that mediated by clathrin are present in these epithelial cells.
The aim of the present study was to examine the distribution of caveolin in kidney epithelial cells and to determine whether it is located in those cell types in which clathrin-independent endocytosis is prevalent. Our results show that caveolin has a distinctive basolateral pattern of distribution in several epithelial cell types along the urinary tubule but that it is not detectable in intercalated cells where H+-ATPase-coated vesicles are predominantly involved in endocytosis.
Materials and Methods
Antibodies
Two anti-caveolin monoclonal antibodies (MAbs) were used in this study. One recognizes only the α-isoform of caveolin-1 (MAb 2234), and the other recognizes both the α- and β-isoforms (MAb 2297). All of the positive results presented were obtained using the latter antibody, suggesting that the isoform detected in the kidney by immunocytochemistry was the β-isoform. Antibodies against caveolin-3, found in muscle cells (Tang et al. 1996), did not label renal epithelial cells (results not shown). The characterization of these antibodies has been previously described (Scherer et al. 1995). Other antibodies used were raised against the common C-terminus of the AE1 and AE2 Cl-/HCO3 - anion exchanger (provided by Dr. Seth Alper, Beth Israel Hospital, Boston), the C-terminus of the 56-kD “kidney” subunit of the vacuolar H+-ATPase (provided by Dr. Stephen Gluck, Washington University, St Louis), or the C-terminal 14 amino acids of the aquaporin 2 (AQP2) water channel (Sabolic et al. 1995). These antibodies have been extensively characterized and used in previous immunocytochemical studies (Alper et al. 1989; Nelson et al. 1992; Sabolic et al. 1995).
Western Blotting
To confirm specificity of the anti-caveolin antibodies, Western blots of kidney outer medulla were performed as previously described (Sabolic et al. 1995). Intestine and epididymis were examined as control tissues that contain abundant smooth muscle cells. After separation by SDS-PAGE, proteins were transferred to Immobilon for immunoblotting. Antibodies were applied to Immobilon strips at a concentration of 1:1000. Recognized bands were visualized by enhanced chemiluminescence (ECL) after application of secondary goat anti-mouse antibodies coupled to horseradish peroxidase.
Animals
Male Sprague–Dawley rats were anesthetized and kidneys were fixed in vivo with PLP buffer containing 2% paraformaldehyde, 10 mM sodium periodate, 70 mM lysine (PLP), and 5% sucrose as previously described (McLean and Nakane 1974; Breton et al. 1995).
For preparation of 4-μm sections, tissues were cryoprotected in 30% sucrose before sectioning with a Reichert Frigocut microtome using disposable knives. For 1-μm sections, tissues were immersed in 2.3 M sucrose for at least 1 hr before freezing in liquid nitrogen and sectioning on glass knives in a Reichert FC4D ultracryomicrotome as previously described (Breton et al. 1995).
Immunostaining
Tissue sections were immunostained as previously described (Breton et al. 1995). An initial treatment with 1% sodium dodecyl sulfate (SDS) for 5 min was essential to reveal caveolin antigenicity (Brown et al. 1996). After blocking with PBS/1% BSA, primary antibody (1:50) was applied for 2 hr at room temperature. Secondary anti-mouse antibodies (diluted 1:150) coupled to CY3 (Jackson Immunologicals; West Grove, PA) were applied for 60 min. Sections were mounted in Vectashield anti-fading solution (Vector Labs; Burlingame, CA) diluted 2:1 in 0.1 M Tris-HCl, pH 8.0.
Some sections were double stained to confirm the identity of the cells that were positive or negative for caveolin in the collecting duct. After application of the anti-caveolin antibody followed by secondary antibody coupled to CY3, rabbit polyclonal antibodies against either the AQP2 water channel (a principal cell marker, affinity-purified, diluted 1:4), AE1 (a marker of A-intercalated cells, diluted 1:1600), or the 56-kD subunit of the H+-ATPase (a marker of A- and B-intercalated cells, diluted 1:100) were applied for 2 hr, followed by a goat anti-rabbit IgG coupled to FITC, diluted 1:60 (Jackson Immunologicals).
Sections were photographed on a Nikon FXA microscope and some color images were captured using an Optronics 3-bit CCD color camera (Optronics Engineering; Goleta, CA), and IP Lab Spectrum (Scanalytics; Vienna, VA) acquisition and analysis software running on a Power PC 8500. Separate images for CY3 and FITC staining were obtained from double stained specimens, and the individual images were color-separated into their RGB components. The red (for CY3) and green (for FITC) components were merged and the composite images were imported as TIFF files into Adobe Photoshop 3.04 for size reduction and printing on a Tektronix Phaser 440 dye-sublimation color printer.
Endosome Labeling by FITC–Dextran Uptake
Some rats were anesthetized as above and injected with 1 ml of a 25 mg/ml solution of FITC–dextran (10,000 kD; Molecular Probes, Eugene, OR) into the jugular vein, as previously described (Lencer et al. 1990). Kidneys were fixed by perfusion-fixation with PLP as above, 15 min after injection of the tracer. Cryostat sections of tissue in which endosomes had been labeled with FITC–dextran in this way were double stained with either anti-caveolin or anti-H+-ATPase antibodies, followed by secondary antibodies coupled to CY3 as above. Images were captured digitally and merged using the IP Lab Spectrum program, as described above.
Electron Microscopy
Conventional Electron Microscopy
Caveolae (pinocytotic invaginations) and other coated membrane vesicles were detected in renal epithelial cells by conventional electron microscopy in rat kidneys fixed with 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer.
Immunogold Electron Microscopy
PLP-fixed kidneys were used for immunogold electron microscopy on ultrathin frozen sections as previously described (Sabolic et al. 1995). Ultrathin frozen sections cut on a Reichert Ultracut E microtome with an FC4D cryoattachment were picked up on nickel grids and first treated for 1 min with a drop of 0.1% SDS before washing, blocking in 1% BSA for 10 min, and incubation with anti-caveolin antibody (1:50) for 1 hr. After washing in PBS, sections were labeled directly using goat anti-mouse IgG coupled to 8-nm gold particles (Amersham; Poole, UK).
Results
Western Blotting
The anti-caveolin α/β antibody MAb 2297 detected in the kidney a single band at 21 kD, corresponding to the β-isoform, whereas in the intestine a single band at a slightly higher molecular weight, probably representing the α-isoform, was observed. Both α- and β-isoforms are present in the epididymis (Figure 1). The caveolin band in the inner stripe of the outer medulla was fainter than in intestine or epididymis, in which caveolin-containing smooth muscle cells are very abundant. These results demonstrate that, in the kidney, only a single band at the expected molecular weight of the β-isoform of caveolin is recognized (Scherer et al. 1995).
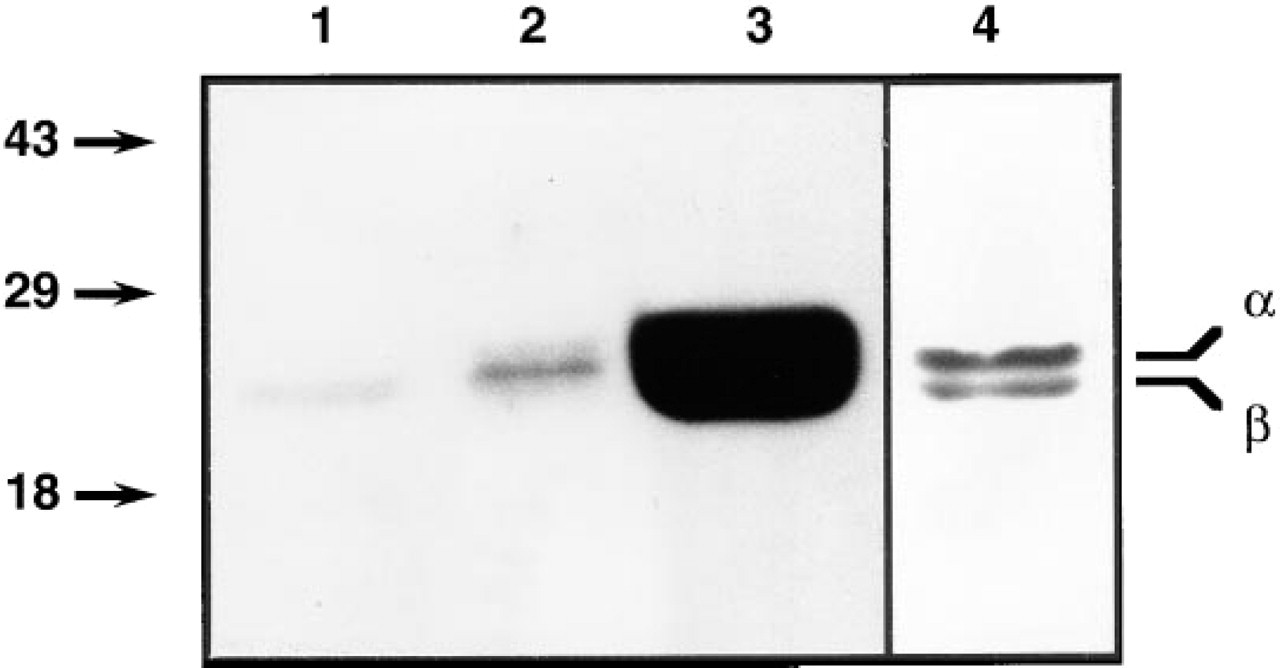
Western blot of kidney inner stripe (Lane 1), intestine (Lane 2), and epididymis (Lanes 3 and 4) using a monoclonal anti-caveolin-1 antibody, MAb 2297, that recognizes the α (24-kD)- and β (21-kD)-isoforms of the protein. Only the 21-kD isoform is detected in the kidney. Lane 4 is from the same transfer as Lane 3, but the time of exposure of the ECL was greatly reduced to distinguish the two distinct bands present in the epididymis.
Immunofluorescence Microscopy
Only the antibody against both α- and β-caveolin gave positive immunostaining in the kidney. In contrast, α-cave-olin-1 and caveolin-3 antibodies did not label renal epithelial cells (not shown). These results indicate that the isoform localized in the kidney is probably β-caveolin-1.
Cortex
Caveolin was most abundant at the basolateral pole of distal convoluted tubules and connecting tubules, where its pattern of staining suggested an association with the highly infolded basolateral plasma membrane of these cells (Figures 2 and 3A-3C). The apical pole was unstained. In the connecting segment, basolateral staining was present in some cells, whereas others were unstained. In the collecting duct, a mosaic pattern of basolateral staining was also seen (Figure 3D). Identification of the negative cells as intercalated cells was confirmed by double staining (see below). In addition to tubule staining, the parietal epithelium of Bowman's capsule was positive and some cells in the glomerulus were labeled (Figures 2A and 2B). Peritubule capillaries were unstained, but both smooth muscle cells and endothelial cells were brightly positive in small vessels, including afferent and efferent glomerular arterioles (Figure 2B). Proximal tubule cells from all segments (S1, S2, S3) showed no detectable staining (Figures 2A and 3A), and cortical thick ascending limbs of Henle were negative.
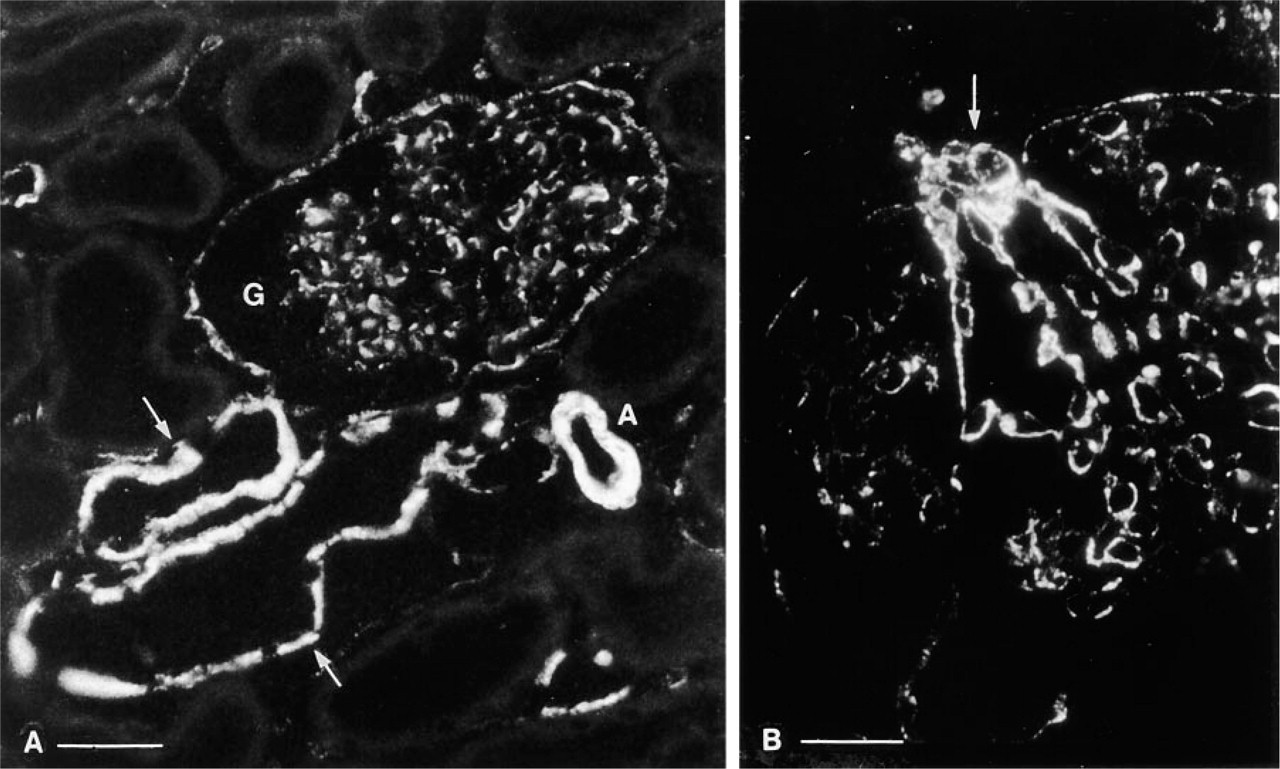
Caveolin immunostaining of rat kidney cortex. (
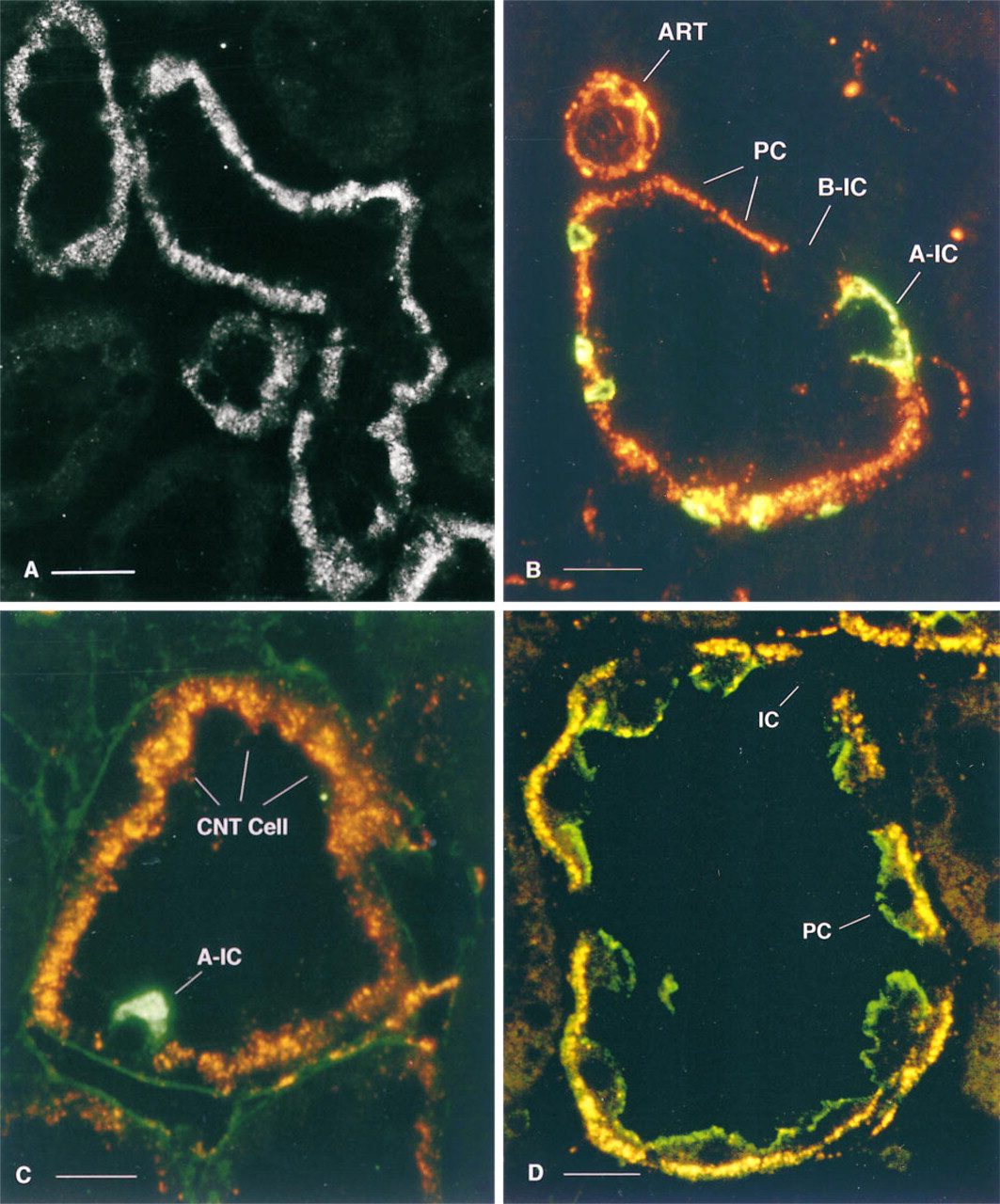
Double Staining
In connecting segments and collecting ducts, all cells positive for caveolin were negative for both AE1 (Figure 3B) and the H+-ATPase 56-kD subunit (Figure 3C), but were apically stained with anti-AQP2 antibodies (Figure 3D). Caveolin-negative cells all contained the 56-kD H+-ATPase subunit (Figure 3C), identifying them as intercalated cells (Brown and Breton 1996). Among the caveolin-negative cells, some showed bright basolateral AE1 staining, whereas others were AE1-negative (Figure 3B), indicating that both A- (AE1-positive) and B (AE1-negative)-intercalated cells (Alper et al. 1989) were unstained with the anti-caveolin antibody.
Outer Medulla: Outer Stripe and Inner Stripe
Collecting duct principal cells (AQP2-positive) showed an irregular, punctate basolateral staining for caveolin (Figures 4A and 4B, inner stripe). Intercalated cells, identified by an absence of AQP2 staining and by the presence of basolateral AE1 and apical H+-ATPase (not shown), did not contain detectable caveolin. S3 proximal tubule segments in the outer stripe (not shown) and thick ascending limbs were also unstained (Figures 4A and 4B). Endothelial cells of some vasa recta in the vascular bundles were brightly stained, whereas others were not fluorescent. Thin descending limbs of Henle inside the vascular bundles (descending thin limbs of short-looped nephrons) were stained (not shown).
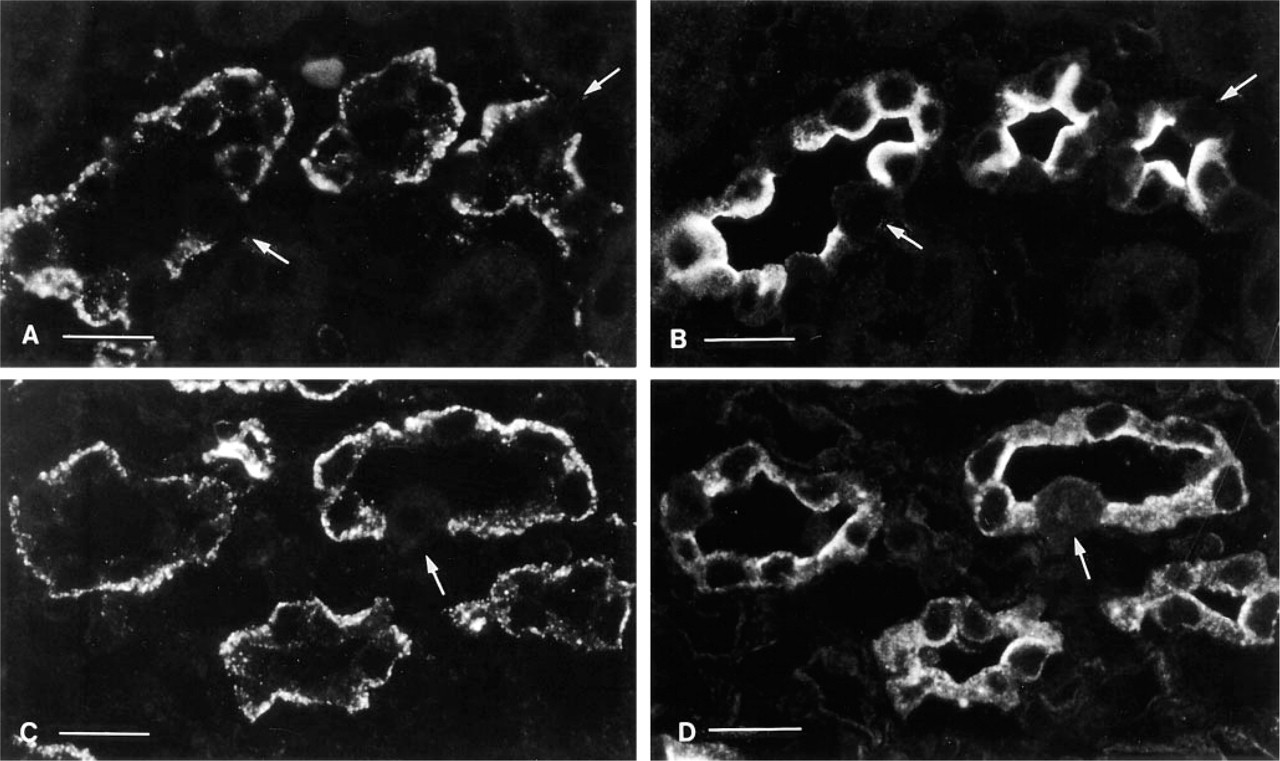
Composite plate showing the inner stripe of the outer medulla (
Inner Medulla/Papilla
Collecting duct principal cells, vasa recta, and some thin limbs of Henle were labeled. Intercalated cells comprise only 5–10% of the collecting duct epithelium in the initial portion of the inner medulla, and then disappear completely from collecting ducts in the middle and the tip of the papilla. In these regions, most of the epithelial cells (principal cells) were stained with both caveolin (Figure 4C) and AQP2 (Figure 4D). The papillary surface epithelium was unstained.
FITC–Dextran Uptake
As previously reported, FITC–dextran injected into the jugular vein of rats is a fluid-phase marker of apical endocytosis in the kidney (Lencer et al. 1990). Intercalated cells showed a vigorous uptake of this marker (Brown and Breton 1996). Principal cells also internalized FITC–dextran, although much less than intercalated cells. Figures 5A-5C show an FITC-labeled inner medullary collecting duct double stained with anti-caveolin antibodies. Caveolin staining was restricted to the basolateral pole of principal cells (Figures 5A, red), and apical endocytotic vesicles in intercalated cells and principal cells contained FITC–dextran (Figure 5B, green). The merged image (Figure 5C), shows that caveolin and FITC–dextran are not co-localized. Figures 5D-5F show a similar inner medullary collecting duct, but double stained to localize H+-ATPase. The vacuolar H+-ATPase is concentrated at the apical pole of intercalated cells (Figure 5D, red), where FITC-labeled endosomes also accumulate (Figure 5E, green). The merged image confirms that the H+-ATPase is co-localized in the FITC-labeled endosomes, resulting in the appearance of yellow apical vesicles (Figure 5F).
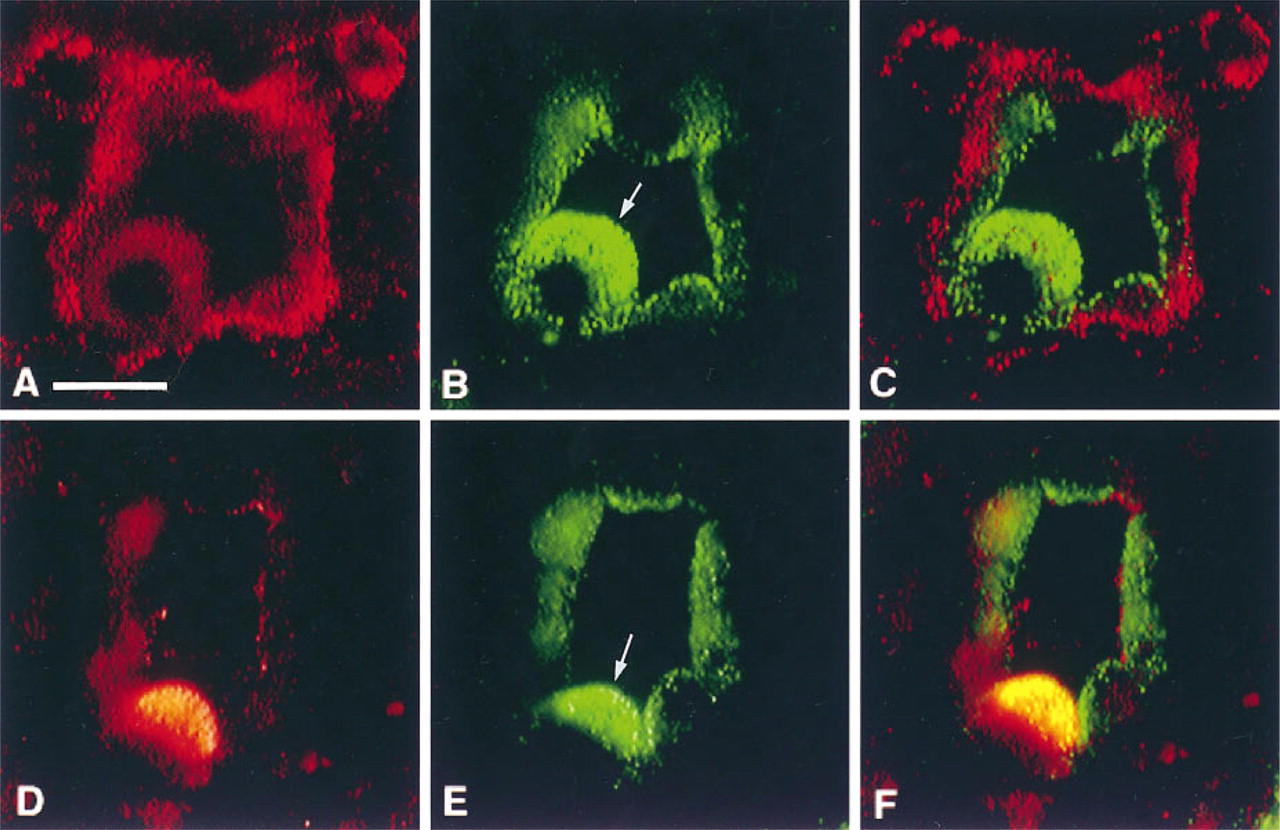
Comparison of caveolin and H+-ATPase antigenicity with the localization of FITC–dextran (green) in apical endosomes of intercalated cells (arrows) and adjacent principal cells in the inner medulla. The section shown in
Electron Microscopy
In many cell types, caveolae were associated with the basolateral plasma membrane. Figures 6A and 6B show the basal region of a connecting tubule epithelial cell (Figure 6A) and a collecting duct principal cell (Figure 6B). The basal plasma membrane infoldings of the connecting tubule cell penetrate much more deeply into the cytoplasm, and caveolae are found at different levels along these membrane infoldings. In contrast, the basal infoldings of the principal cell are much shallower but also contain caveolae. The marked difference in geometry of these infoldings is responsible for the different width of the immunofluorescent band of caveolin staining seen in these cell types (compare Figures 3C and 3D). Caveolae were not detected in the basolateral or apical plasma membranes of intercalated cells or proximal tubule epithelial cells, consistent with the absence of labeling with anti-caveolin antibodies. In contrast to epithelial cells, large numbers of caveolae were present in endothelial cells of the descending vasa recta (Figure 6C), arterioles, and the glomerular capillaries, as well as in smooth muscle cells and adipocytes.
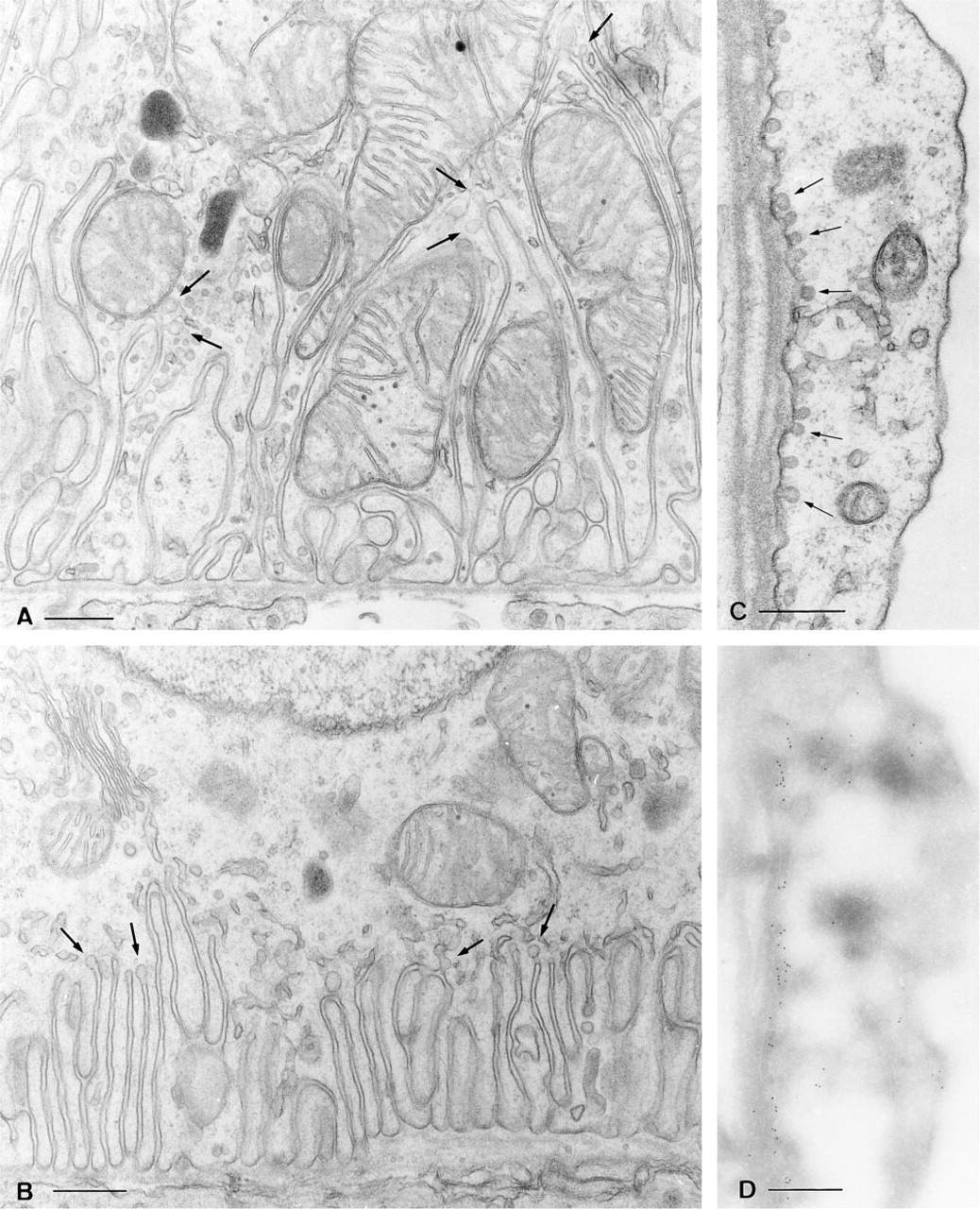
Electron microscopy showing the basolateral region of a connecting tubule (CNT) epithelial cell (
Morphologically recognizable caveolae were rarely seen in ultrathin frozen sections, probably because of the SDS treatment that was required to reveal caveolin antigenicity. Nevertheless, heavy immunogold labeling for caveolin was found in cells in which caveolae were abundant. As an example, an endothelial cell from a descending vasa recta is shown in Figure 6D. Many gold particles are located at the basolateral pole of this cell, precisely the site where caveolae are concentrated, as shown by conventional electron microscopy in Figure 6C. In epithelial cells, some small clusters of gold particles were found over the basal infoldings of connecting segment and collecting duct principal cells (not shown), but lack of membrane resolution made it impossible to attribute this staining to the presence of the single, isolated caveolae that occur in these cells.
Discussion
Epithelial cells lining the urinary tubule carry out many specialized functions that, in several cases, involve the exo- and endocytotic recycling of membrane proteins (Brown and Stow 1996). Although clathrinmediated internalization of some of these proteins has been described (Brown et al. 1988; Strange et al. 1988; Maunsbach and Christensen 1992), the nature of the endocytotic pathway in other cases remains undefined. Notably, the vacuolar H+-ATPase is moved to and from the plasma membrane of proton-secreting collecting duct intercalated cells in “coated” vesicles that do not contain clathrin (Brown and Orci 1986; Brown et al. 1987b).
Our present results show that the coat protein caveolin is not detectable in intercalated cells, whereas caveolin is abundant in some other kidney cell types. The distal convoluted tubule had the brightest staining, which was, as in principal cells, uniquely basolateral. Interestingly, a Ca2+-ATPase has been located in caveolae in many cell types and is abundant in the basolateral membrane of the distal tubule (Fujimoto 1993). In addition, the presence of basolateral pinocytotic invaginations in connecting tubule cells has been previously described (Kaissling and Kriz 1992). Caveolae have not been identified on the apical membranes of kidney epithelial cells.
The role of caveolin in the basolateral membrane of principal cells is unknown, but a variety of proteins, including signal transduction proteins, have been found in caveolae (Parton 1996; Couet et al. 1997). These structures contain at least some of the molecules that are involved in the response to vasopressin, such as the stimulatory G-protein Gαs and adenylate cyclase (Wagner et al. 1972; Sargiacomo et al. 1993; Lisanti et al. 1994). Our previous results have also localized the heterotrimeric G-proteins Gαs and Gαi2 on the basolateral membrane domain of vasopressin-sensitive principal cells (Stow et al. 1991). The present results show that aquaporin 2 (AQP2), the vasopressin-sensitive water channel (Fushimi et al. 1993), is not preferentially located in caveolae.
It has been proposed that GPI-anchored membrane proteins are concentrated in caveolae in several cell types (Sargiacomo et al. 1993; Zurzolo et al. 1994). However, these proteins are usually targeted apically (Lisanti et al. 1990), whereas caveolin in kidney epithelial cells is predominantly basolateral. Furthermore, the isoform of caveolin detected in this study is not obligatorily associated with GPI-anchored proteins, because thick ascending limb cells that contain the GPI-linked proteins carbonic anhydrase Type IV and Tamm–Horsfall glycoprotein/uromodulin on their plasma membranes (Brown and Waneck 1992) did not show any detectable labeling with anti-caveolin antibodies. Nor was staining detectable in proximal tubules, which express several GPI-linked apical membrane proteins (Brown and Waneck 1992). Furthermore, it has recently been shown that caveolin is not necessary for sorting of GPI-anchored proteins to the apical membrane of transformed intestinal epithelial Caco-2 cells (Mirre et al. 1996).
The absence of caveolin and clathrin from H+-ATPase-recycling vesicles in intercalated cells suggests that these vesicles represent a distinct class of coated vesicle. Whether the H+-ATPase subunits that form this coat have roles in vesicle trafficking in addition to (or in parallel with) their function as ion-transporting proton pumps in intercalated cells and related H+-secreting cells (Brown and Breton 1996) is a matter for future investigation. Indeed, recent work has shown that one subunit of the H+-ATPase complex is homologous to a synaptophysin binding protein found in nerve synapses (Siebert et al. 1994), and a 50-kD coated vesicle α-adaptin subunit also co-purifies with the proton pump under some conditions (Liu et al. 1994). This hypothesis of subunit promiscuity is also supported by our own observation that endocytotic vesicles derived from clathrin-mediated endocytosis in principal cells lack several subunits of the H+-ATPase and do not acidify their lumen, yet they do contain the “kidney” isoform of the 56-kD subunit of the proton pump (Sabolic et al. 1992). Therefore, although this subunit is clearly important for proton pump function, it may have a more promiscuous role when associated with other proteins in non-acidic vesicles.
The presence of caveolin in defined membrane domains further supports the notion that different renal epithelial cells, and even different membrane domains in the same cell, have evolved specialized membrane trafficking pathways that are involved in the regulation and modulation of epithelial transport pathways in this complex tissue.
Footnotes
Acknowledgements
Supported by grants from the National Institutes of Health, DK 42956 (DB) and DK38452 (SB). ML was supported by an NIH FIRST award GM-50443, by a grant from the W. M. Keck Foundation to the Whitehead Fellows program, and by a grant from the Elsa U. Pardee Foundation. SB was partially supported by a grant from National Sciences and Engineering Research Council of Canada, by a Hoechst Marion Roussel fellowship from the National Kidney Foundation, and by a Claflin Distinguished Scholar Award from the Massachusetts General Hospital.