
Abstract
Thymidylate synthase (TS) [TYMS; OMIM reference number (188, 350)] is normally considered to be a cytoplasmic enzyme. However, a few reports have suggested it may also be present in the nucleus. To explore this in more detail, we used a highly specific polyclonal antibody to TS and a combination of techniques, including immunocytochemistry, confocal microscopy, cell fractionation, and Western blotting. We developed cell line HeLa-55, a HeLa derivative that grossly overexpresses TS. Although the vast majority of TS was in the cytoplasm, some TS also was seen in the nucleus. TS in parental HeLa cells and in normal human fibroblasts was seen exclusively in the cytoplasm. HeLa-55 cells exposed to 5-fluorodeoxyuridine were fractionated and examined by Western blotting. Interestingly, both free TS and the ternary complex of TS were seen in the cytoplasmic fraction but only free TS was detected in the nuclear fraction. Amongst different cell lines examined, HCT-15 and normal fibroblasts showed no nuclear TS, HCC-2998 and SW-620 showed a small amount of nuclear TS, and HT-29, RKO, and HCT-116 showed a strong nuclear TS signal. Nuclear staining was clearly evident in some clinical colorectal specimens, both normal and malignant. This staining was definitively shown to be TS by competition with recombinant TS protein. A putative leucine-rich nuclear export sequence was identified but its function could not be confirmed. We conclude that small amounts of TS protein is present in the nucleus of some cell types but further work is needed to determine the significance of this observation.
T
Although TS is usually considered to be a cytoplasmic enzyme (Kucera and Paulus 1986; Johnston et al. 1991, 1995; Suzuki et al. 1998; Yamachika et al. 1998; Paradiso et al. 2000), there is some suggestion in the literature that TS may also be present in the nucleus of some mammalian cells (Johnston et al. 1991; Samsonoff et al. 1997). The strongest evidence for the presence of nuclear TS comes from the studies of Wong et al. (2001), who examined primary and metastatic colorectal carcinoma using monoclonal antibody TS106 (Johnston et al. 1991). Nuclear TS was identified immunohistochemically in adenocarcinoma cells of primary colorectal cancer specimens, but also in normal basal crypt colonocytes and germinal center cells. High TS nuclear expression was correlated with poorer response to 5-FU. The presence of nuclear TS in primary carcinomas was associated with high nuclear TS in metastatic lesions from the same patients.
Immunohistochemical studies are critically dependent upon the choice of antibodies used. Our laboratory developed and extensively characterized a polyclonal antibody to recombinant human TS (Haqqani et al. 1999). The present study extends earlier findings about the presence of TS in the nucleus. To ensure that the nuclear signal was indeed specific for TS, recombinant full-length TS protein (used originally as the antigen) was used as a competitor for immunohistochemistry, immunocytochemistry, and immunofluorescence. Our studies provided definitive evidence that TS is present in the nucleus of a number of colorectal cancer cell lines and tissues.
Materials and Methods
Antibodies
Sheep and rabbit (hTS7.4) anti-hTS antisera used in these studies were obtained from Rockland Immunochemicals (Gilbertsville, PA) or Roche Diagnostics (Laval, Canada). These polyclonal antibodies were raised against recombinant (his)6-rhTS (Haqqani et al. 1999). Horseradish peroxidase (HRP)-conjugated rabbit anti-sheep secondary antibody (cat. no. 613–4328) and goat anti-lactate dehydrogenase polyclonal antibody (cat. no. 100–1173) were also from Rockland Immunochemicals. CY3-conjugated donkey anti-sheep secondary antibody (cat. no. 716-165-147) and HRP-conjugated donkey anti-goat secondary antibody (cat. no. 705-036-147) were from Jackson ImmunoResearch Laboratories (West Grove, PA). HRP-conjugated anti-rabbit/anti-mouse secondary antibody (“Polymer”) was from DAKO (Carpinteria, CA).
The specificity of the anti-hTS antibody was verified using recombinant (his)6-tagged hTS protein (rhTS) as a competitor. This protein was expressed in Escherichia coli, purified by affinity chromatography on a nickel column and then twice-purified by SDS-PAGE as previously described (Haqqani et al. 1999). After electroelution, the protein was precipitated with ethanol and dissolved as a stock solution at a concentration of 0.5 mg/ml in 0.2% SDS (sodium dodecyl sulfate). The detergent was required to maintain the denatured TS protein in solution. When used as a competitor for Western blot analysis, immunofluorescence, immunocytochemistry, or immunohistochemistry, the stock was diluted at least 10-fold in phosphate-buffered saline (PBS) such that the concentration of SDS was ×0.02%. When used as a competitor, essentially similar results were obtained using anti-TS antiserum preincubated with rhTS for 1 hr at room temperature or overnight at 4C. Control experiments were carried out in which the same antiserum was preincubated with S-buffer (PBS containing 0.02% SDS without rhTS).
Selective depletion of anti-TS antibody from antiserum was performed using affinity chromatography. Sheep anti-TS antiserum was diluted 1:100 and passed 20 times through a bed of CN Br-activated Sepharose 4B (Sigma-Aldrich Chemicals; St. Louis, MO) to which rhTS protein was coupled (Haqqani et al. 1999). As a control, diluted anti-serum was passed through a similar matrix containing immobilized bovine serum albumin (BSA) (OmniPur BSA, Fraction V; EM Science-MERCK kGaA, Darmstadt, Germany).
Cell lines
HeLa cells were obtained from ATCC (Rockville, MD). A series of 5-FUdR-resistant HeLa cells clones was isolated by stepwise selection in increasing concentrations of the drug up to 0.1 μM over several weeks. Clone HeLa-55 overexpressed TS protein most strongly and was shown to have a highly amplified TS gene (data not shown). The following colon cancer cell lines from the NCI anti-cancer screening panel of 60 cell lines were used: HCT-116; HCT-15, HCC-2998, HT-29, and SW-620; RKO is a colon cancer line originally derived by Brattain et al. (1984). Normal primary human fibroblasts were from neonatal foreskin. Cells were cultured as monolayers in Dulbecco's modified Eagle's medium (GIBCO-BRL; Burlington, Canada) supplemented with 10% fetal calf serum in 100 mm dishes in 5% CO2 at 37C. RKO cells were maintained in McCoy's 5A media plus 10% fetal calf serum (GIBCO-BRL).
Immunofluorescence
Cells were grown on 22 mm2 glass coverslips in 10 cm dishes. Log phase cultures were used in all experiments. Coverslips were removed and washed twice with PBS. All subsequent steps were carried out at room temperature. Cells were fixed for 10 min with acetone:methanol (1:1). Coverslips were blocked for 30 min with Universal Blocking solution (DAKO). All antibody dilutions were in Antibody Diluting buffer (DAKO). Coverslips were incubated for 1 hr with primary anti-TS antibody (1:500 dilution). They were washed 5 times (5 min per wash) with PBS. Cells were then incubated for 30 min with secondary anti-sheep IgG-CY3 antibody (1:1000 dilution) and washed again. The coverslips were mounted onto slides with Fluorescent Mounting medium (DAKO) containing 5 μg/ml Hoechst 33258 (Sigma-Aldrich). Stained cells were examined and photographed with a Zeiss Axiophot fluorescence microscope (Carl Zeiss Canada Ltd.; North York, Canada). Confocal images were obtained using a Zeiss inverted LSM 410 confocal laser scanning microscope. Z-series images were obtained through the depth of cells using a step size range of 1-2 μm.
Immunocytochemistry
Coverslips containing cells in log phase were removed and washed twice with PBS. All subsequent steps were carried out at room temperature. Cells were fixed for 10 min with acetone:methanol (1:1). Coverslips were blocked for 30 min with Casein Blocking Solution in PBS [2% bovine milk casein (Sigma-Aldrich), 2 mM cyclohexanediamine tetraacetate (Sigma-Aldrich), in PBS]. All antibody dilutions were in PBS. Coverslips were incubated for 1 hr with primary anti-TS antibody (1:500 dilution). They were washed 5 times (5 min per wash) with PBS. Cells were then incubated for 30 min with secondary rabbit anti-sheep-IgG-HRP antibody (1:1500 dilution) and washed again. Visualization was achieved with diaminobenzidine substrate. Coverslips were mounted onto slides with Permount Mounting medium (Fisher Scientific; Toronto, Canada). Stained cells were examined and photographed with a Zeiss Axiophot microscope (Carl Zeiss Canada).
Immunohistochemistry
Colorectal cancer specimens were obtained from the Ottawa Colorectal Cancer Bank; they had been fixed in an alcohol-based fixative, GenoFix (DNA Genotek; www.DNAgenotek.com). 3 μm sections were cut from the paraffin-embedded tissue blocks, then deparaffinized and rehydrated using standard methods. For antigen retrieval, slides were heated for 5 min in a pressure cooker using 10 mM citrate buffer at pH 6.0. Endogenous peroxidase was inactivated with 0.3% H2O2 in methanol. Slides were blocked with 20% normal goat serum. Sections were incubated overnight at 4C with rabbit (hTS7.4) anti-hTS antiserum at a 1:100 dilution. Sections were then incubated with Dakocytomation Envision+ rabbit reagent K4002 (Dakocytomation; Mississauga, Canada) for 30 min. Dakocytomation diaminobenzidine substrate K3467 was used for color development. Sections were counterstained using hematoxylin. Positive controls included human tonsillar tissue. Negative controls included the omission of the primary antibody. Positive and negative controls were run in parallel. Additional negative controls utilized recombinant (his)6-tagged hTS protein (rhTS) as a competitor.
Preparation of Whole Cell and Nuclear Extracts
Cultured cells were detached by light trypsin treatment, then washed twice and suspended in cold PBS. To prepare whole cell extracts, an equal volume of 2 × SDS sample buffer (100 mM MOPS, pH 6.8, 4% SDS, 20% glycerol, 2% b-mercaptoethanol) was added. Extracts were boiled for 10 min and then sonicated briefly to reduce viscosity. Protein was quantified using fluorescamine (Sigma-Aldrich) (Udenfriend et al. 1972), using BSA as a standard. For isolation of nuclei, cells were washed twice in PBS and suspended in nuclear extraction buffer (10 mM Tris-HCl, pH 7.5, 10 mM KCl, 1 mM CaCl2, 1 mM MgCl2, 0.1 mM spermidine, 2 μg/ml aprotinin and 1 mM PMSF) at ∼1 × 106 cells/ml. After 20 min incubation at OC, the swollen cells were homogenized (50 strokes) with a tight-fitting glass Dounce homogenizer. Samples were centrifuged (800 × g) at 2-4C and the pellet was washed twice with nuclear extraction buffer containing 0.1% NP-40. Protein in the nuclear pellet was extracted by dispersion in SDS sample buffer, brief sonication and heating at 100C for 15 min.
Western Blot Analysis
Proteins in cell or nuclear extracts in SDS sample buffer were resolved by 12% discontinuous SDS-PAGE and then electrophoretically transferred to a PVDF membrane (Millipore; Nepean, Canada) at 15V for 18 hr in transfer buffer (3 mM Na2CO3, 10 mM NaHCO3, 10% methanol) (Haqqani et al. 1999). The membrane was stained with Ponceau S and then washed twice in 10 mM Tris-HCl, 150 mM NaCl, 0.1% Tween-20, pH 8 (TBST). All steps were carried out at room temperature. When using anti-TS as the primary antibody, the membrane was blocked for 1 hr with freshly prepared Casein Blocker solution (100 mM NaCl, 50 mM sodium acetate, pH 6.0, 1% bovine milk casein, 2 mM cyclohexanediamine tetraacetate). When anti-LDH primary antibody was used, the membrane was blocked with 5% horse serum in TBST. Sheep anti-TS antibody was diluted 1:5000 in Casein Blocking solution and goat anti-LDH primary antibody was diluted 1:1000 in TBST. In all cases, the primary antibody was incubated with the membrane for 1 hr. The membrane was washed 5 times in TBST and incubated for 1 hr with a secondary antibody as follows: rabbit anti-sheep-IgG-HRP was used at 1:3000 dilution in Casein Blocking solution; DAKO Polymer anti-mouse-IgG-HRP was used at 1:100 dilution in TBST; donkey anti-goat-IgG-HRP was used at 1:25000 dilution in TBST. The membrane was washed 5 times with TBST, and HRP was detected using the LumiGLO Chemiluminescent Substrate Kit (Kirkegaard and Perry Lab; Gaithersburg, Maryland).
Plasmid DNA Constructs
Green Fluorescent Protein-nuclear export signal (GFP-NES): pEGFP-C1 plasmid was obtained from BD Biosciences Clontech (Mississauga, Canada). Two 46-mer oligonucleotides, hTSNESF (AATTCTATTTACCTGAATCACATCGAGCCACTGAAAATTCAGCTTG) and hTSNESR (GATCCAAGCTGAATTTTCAGTGGCTCGATGTGATTCAGGTAAATAG), encoding the sense and antisense sequences of a putative TS nuclear export signal (NES), respectively, were synthesized. The oligonucleotides were annealed and the resulting fragment consisted of overhanging sequences compatible to EcoR1 and BamH1 sites. The annealed fragment was ligated to EcoR1/BamH1 digested pEGFP-C1. The resulting plasmid, GFP-NES contained the putative NES of TS fused in frame to the carboxyl end of GFP; its sequence was confirmed by sequencing.
H6TS313: PQE-hTS (Haqqani et al. 1999) encodes human TS with the first 5 amino acids deleted and 16 additional amino acids including a 6× histidine tag inserted at the N terminus. This His6-TS (1 kb) fragment was isolated from PQE-hTS using EcoR1/Xba1 and ligated to EcoR1/Xba1 digested pCR3 (Invitrogen; Burlington, Canada).
H6TS248: pcDNA3/HA-hTS29 was isolated as a spontaneously occurring single base pair deletion in the codon for amino acid 248 of human TS (Cowling and Birnboim 1998), which resulted in changes to amino acids 248-259 and a premature stop codon at amino acid 260. An 870 bp fragment containing this frameshift mutation was isolated from pcDNA3/HA-hTS29 using Pst1/Xba1. H6TS-PCR3 was digested with Pst1/Xba1 and the 5167 bp fragment was ligated to the 870 bp fragment of pcDNA3/HA-hTS29.
H6TS186: The nucleic acid fragment corresponding to amino acids 186–313 in H6TS-PCR3 was excised with BglII/Xba1. The remainder of the construct was re-ligated following blunting of the ends with mung bean nuclease.
fAeqTS-pCDNA3 was synthesized to contain full-length TS cDNA. The coding region was obtained from pWHTS-1 (Davisson et al. 1989) and the flanking 5'-UTR containing triple GC-rich repeats (Horie et al. 1995) was synthesized by PCR amplification of genomic DNA. The correctness of the construct was confirmed by sequencing.
Transfection
HeLa and RKO cells were seeded at 1.25 × 105 cells in a 3 cm dish on glass coverslips. The following day, cells were transfected with 2 μg of purified plasmid DNA and 3 μl of Fugene 6 Transfection Reagent (Roche Diagonostics Corp.; Indianapolis, IN), according to the manufacturer's protocol. Forty-eight hours posttransfection, coverslips were removed and immunofluorescence staining was performed as described above.
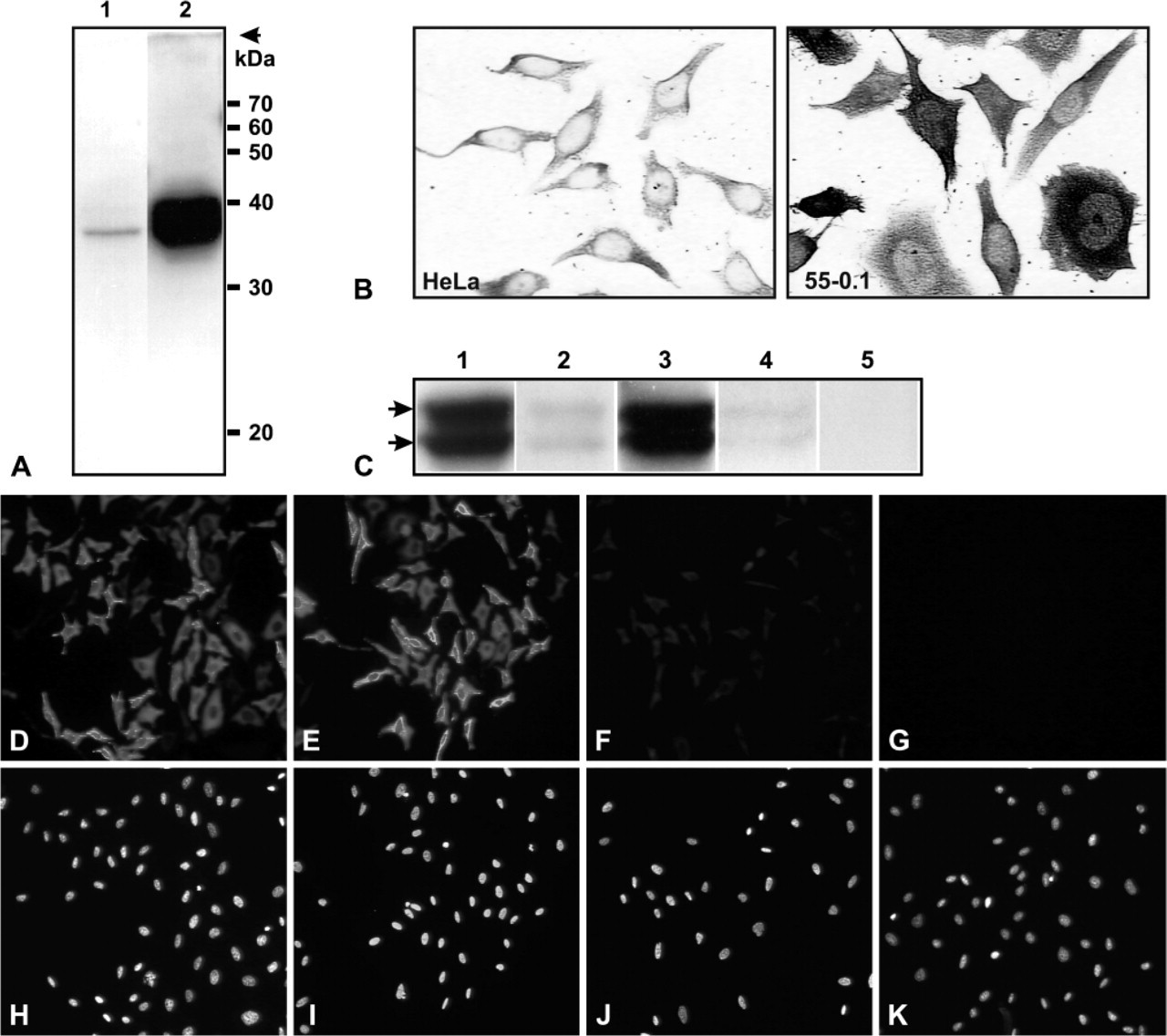
Specificity of the sheep anti-thymidylate synthase (TS) antibody studied by Western blot analysis, immunohistochemistry and immunofluorescence. (
Results
Specificity of Anti-hTS Antisera
The interpretation of results employing immunohistochemistry depends critically on the specificity of the antibodies used. Some previous immunohistochemical or immunocytochemical studies of TS have employed polyclonal anti-TS antibodies whose specificity was not fully documented. We chose to use multiple techniques to confirm the specificity of the primary anti-hTS polyclonal antibodies used in our experiments. Figure 1A (Lane 1) shows Western blot analysis of a HeLa cell extract probed with anti-hTS; the only detectable signal was a band of the expected molecular size for TS (35.7 kDa). Figure 1A (Lane 2) shows similar analysis of an extract of a 5-FUdR-resistant HeLa cell line, HeLa-55, growing in 5-FUdR. Two bands are present (Figure 1A, Lane 2), more clearly seen in Figure 1C. The lower band corresponds to free TS and the upper band to the previously described ternary complex containing TS-5-FdUMP-CH2-THF (Johnston et al. 1991). The high level of TS in HeLa-55 cells is due to amplification of the TS gene (Materials and Methods). No other bands were detected in extracts of either cell line. Figure 1B shows immunocytochemical analysis of the same preparation of HeLa and HeLa-55 cells as used in Figure 1A. There is a correspondence between the amount of TS detected in the two cell lines by the two methods. These results provide strong evidence that both the primary and the secondary antibodies used were specific for TS.
Competition with a specific antigen (usually a synthetic peptide) is another approach commonly used to establish the specificity of antibodies. Since the anti-TS polyclonal antibody was raised against recombinant hTS protein (rhTS) (i.e., not against a synthetic peptide), we prepared highly purified protein and tested this as a competitor. Competition was carried out by either preincubating rhTS with the anti-hTS antibody or using anti-hTS antibody passed through an rhTS-affinity column. Western blot analysis of HeLa-55 cell extracts probed with control and treated antibodies is shown in Figure 1C. Both control antibody and antibody passed through a control affinity column showed strong TS and TS-ternary complex bands (Figure 1C, Lanes 1 and 3). Essentially complete disappearance of the bands was seen using either soluble rhTS or immobilized rhTS as competitor (Lanes 2 and 4). Secondary antibody alone showed no signal (Lane 5). Immunofluorescence staining of HeLa-55 cells using the same antibodies is shown in Figures 1D-1K. An equivalently strong fluorescent signal was seen in these cells when standard buffer (D) or Competition Buffer (F) was used. Pretreatment of the primary antibody with soluble rhTS almost completely eliminated the signal (H). Similar disappearance of signal was seen when immobilized rhTS was used as competitor (data not shown). Fluorescent secondary antibody alone showed no signal (J). The same fields stained with Hoechst 33258 (Sigma-Aldrich) to indicate the position of nuclear DNA are shown in the bottom panels. Taken together, this combination of techniques provides compelling evidence that the primary and secondary antibodies available for this study were highly specific for TS.
Nuclear Localization of TS in an Overexpressing HeLa Cell Line
Cell line HeLa-55, which contains an amplified TS gene, strongly overexpresses TS protein (Figure 1). In addition to the strong staining of the cytoplasm of these cells, nuclear staining also appears to be present (Figure 1B). To confirm this observation, we used immunofluorescence confocal microscopy to study cellular localization (Figure 2). HeLa-55 cells were heterogeneous with respect to levels of TS overexpression. The filled arrowhead follows an example of TS in the nucleus of a strongly overexpressing cell through serial optical sections. Expression of TS was also seen in the nucleus of some low-expressing cells (open arrowhead). This figure provides clear evidence that TS is present in the nucleus of these TS-overexpressing HeLa cells.
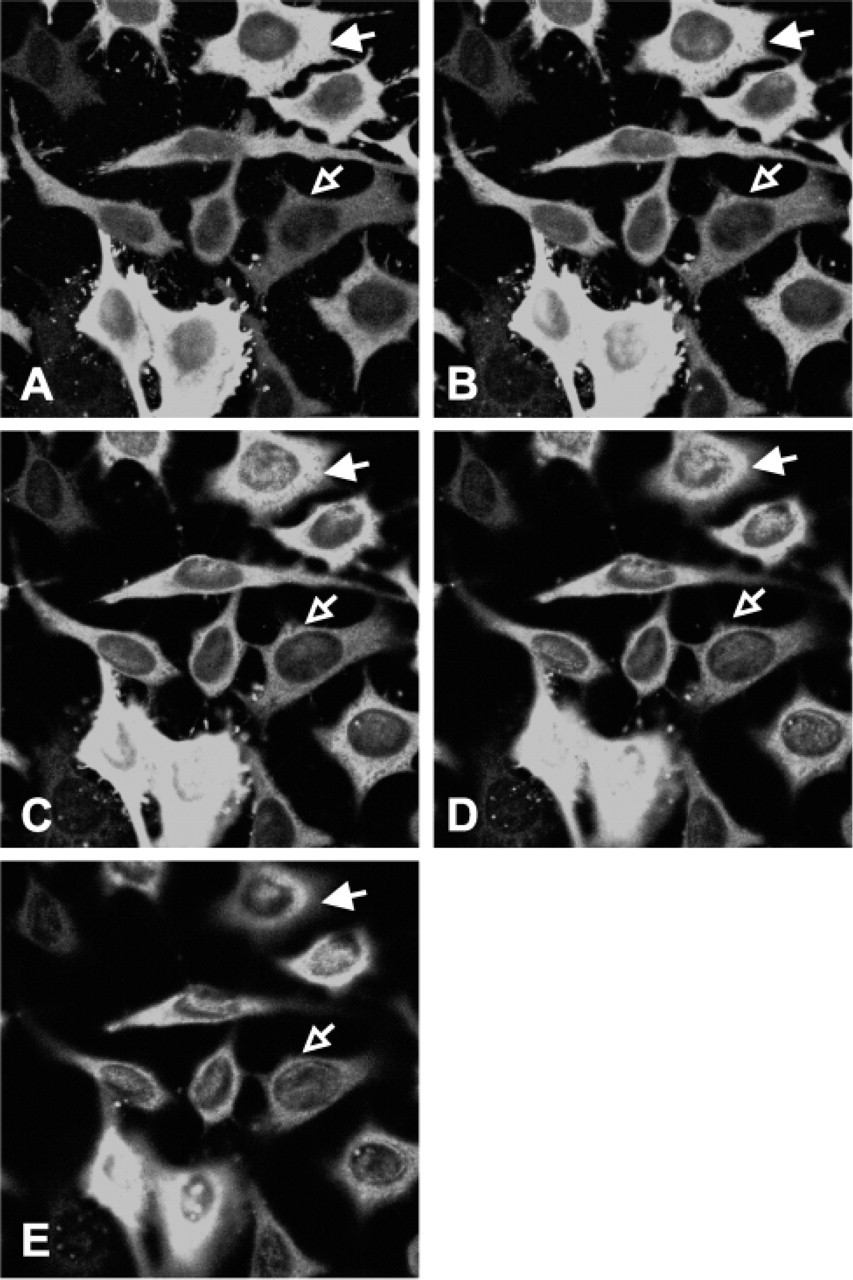
Confocal microscopy of HeLa-55 cells stained by immunofluorescence for thymidylate synthase (TS). Twomm optical sections were taken through cells from bottom to top, (
Nuclear Localization of TS in Other Cell Lines
It was previously reported that TS can be detected in the nucleus of a rat hepatoma cell line (Samsonoff et al. 1997) and in human colon cancer specimens (Wong et al. 2001). To extend these observations, we examined a number of human cell lines for nuclear TS expression. Cell lines included normal human fibroblasts and 6 colorectal cancer lines (HCT-15, HCC2998, SW620, RKO, HCT-116, and HT-29). Amongst the various cell lines examined, differences in both the intensity and localization of TS protein were seen (Figure 3). Normal fibroblasts and HCT-15 cells showed the lowest level of overall TS expression. There was no discernible nuclear expression in fibroblasts and only very low levels in HCT-15 cells. Some nuclear expression was seen in HCC2998 and SW620 cells. The strongest examples of nuclear expression were seen in RKO, HCT-116, and HT-29 cells. To support these observations about nuclear expression of TS, we performed immunofluorescence confocal microscopy on two of these cell lines (Figure 4). Serial optical sections of RKO cells (Figures 4A–4C) showed diffuse nuclear TS staining, whereas HeLa cells (Figures 4D–4F) showed little or no nuclear TS expression. These experiments provide further indication that TS is present in the nucleus of some cell lines.
Western Blot Analysis of TS in Extracts of Isolated Nuclei
As an independent form of evidence to demonstrate the presence of TS in nuclei, we performed Western blot analysis of the nuclear fraction of HeLa, RKO, and HCT-116 cells. RKO and HCT-116 colorectal cell lines previously showed strong nuclear TS staining (Figure 3) and HeLa cells showed little or no nuclear staining (Figure 1B and Figure 4). Total cell extracts and nuclear extracts from an equivalent number of cells were analyzed. A strong TS band was detected in total cellular extracts of all three cell lines (Figure 5A, Lanes 1–3). A TS band was detected in nuclear extracts of RKO and HCT-116 cells (Figure 5A, Lanes 5 and 6), but not HeLa cells (Figure 5A, Lane 4). Lactate dehydrogenase, a cytoplasmic protein marker (Zaman et al. 1999), was used to confirm the absence of cytoplasmic contamination in nuclear extracts.
Nuclear Expression of Free- and Ternary Complex-bound TS
TS-overexpressing HeLa-55 cells grown continuously in the presence of 0.1 μM 5-FUdR were fractionated and a nuclear extract was probed for TS by Western blot analysis. The total cell extract (1/100th the number of cell equivalents) was compared with the nuclear extract (Figure 5B). From the relative intensities of the total cell and nuclear extracts, we estimate that ∼0.5% of the total free TS was present in the nuclear extract. The LDH control confirmed the virtual absence of cytoplasmic contamination in the nuclear fraction. Interestingly, almost none of the ternary complex-bound TS was found in the nuclear extract.
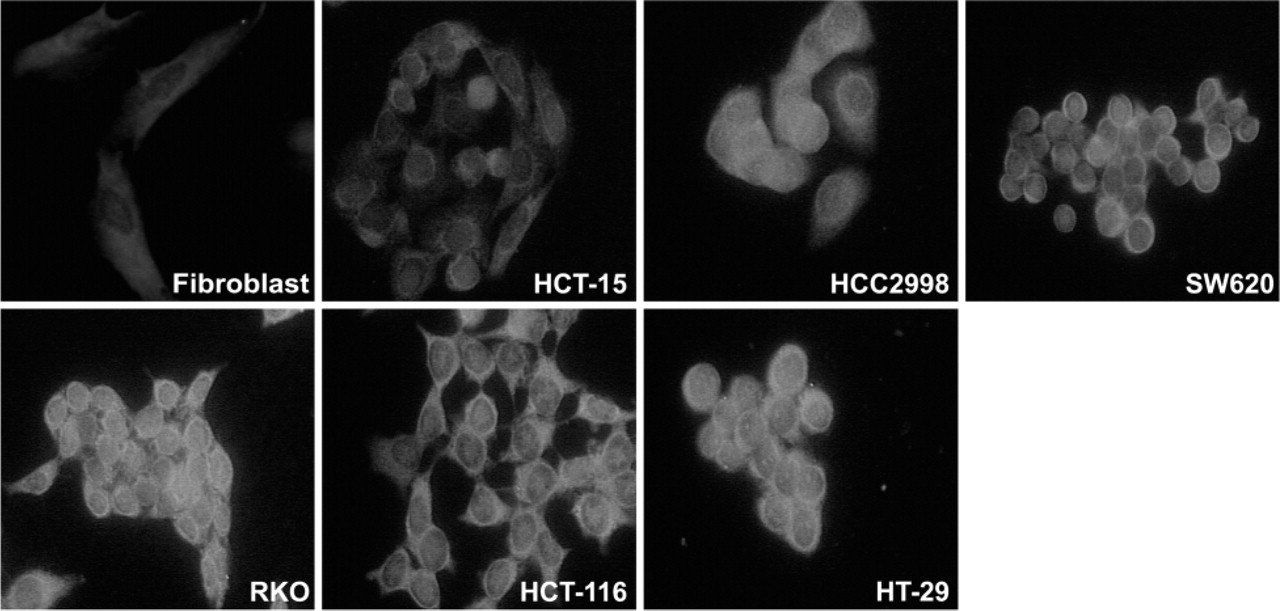
Thymidylate synthase localization in a variety of cell lines using immunofluorescence. Other details are in Materials and Methods.
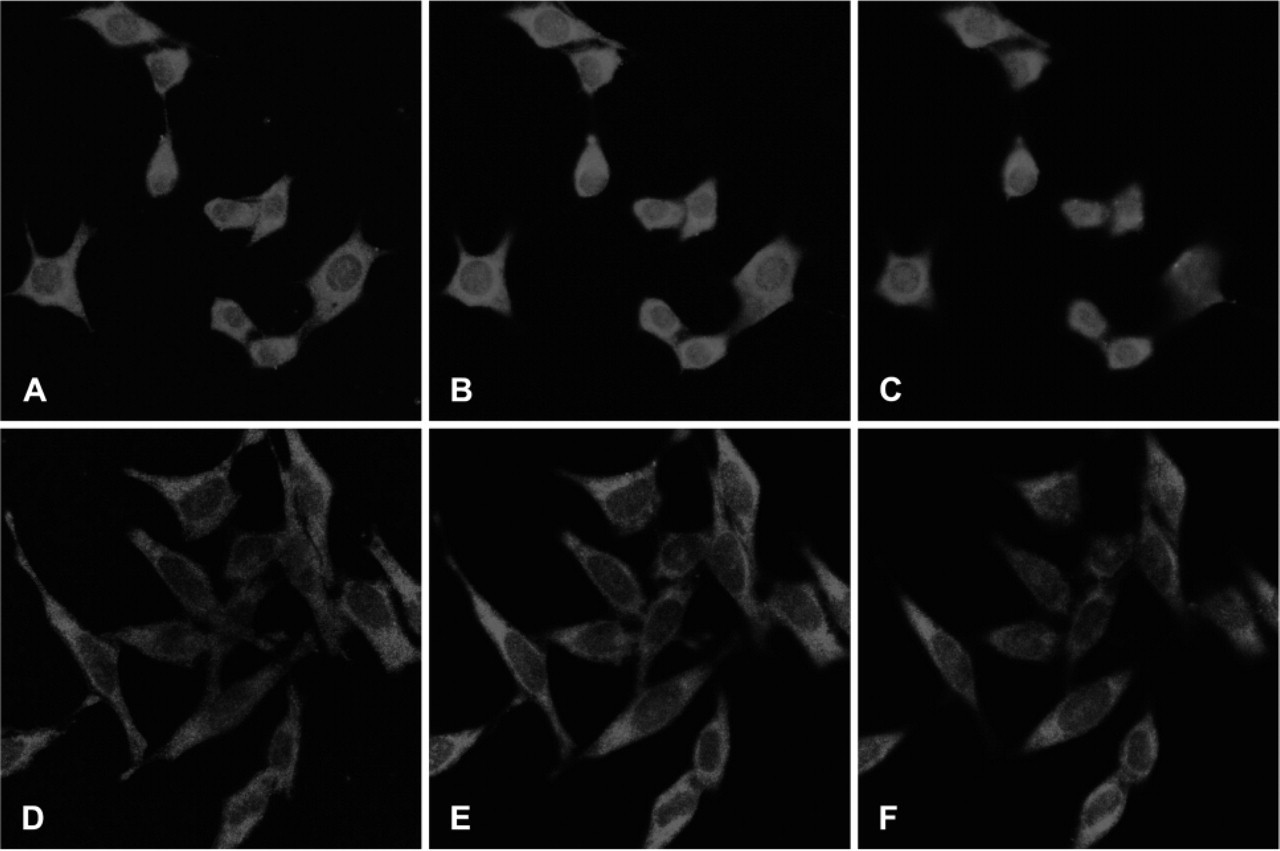
Confocal microscopy of RKO and HeLa cells stained by immunofluorescence for thymidylate synthase. Two-mm optical sections were taken through cells from bottom to top. RKO cells,
Nuclear TS Expression in Colorectal Cancer and Normal Specimens
To extend our findings utilizing cultured cells to clinical colorectal specimens, we examined normal mucosa and colorectal cancer samples from eight patients using immunohistochemistry. Cancer specimens ranged from low to high in the intensity of TS staining, both within the same section and from different patients. An example of a strongly staining region in one colorectal cancer paraffin block is shown in Figure 6. Panel A shows a region stained with anti-TS antibody and Panel B shows a closely related serial section in which the same antibody was competed with rhTS protein. The strong TS signal was virtually eliminated by the presence of the competitor. Noteworthy, TS was detected in both the cytoplasm and the nucleus (A); both signals were similarly decreased by the competitor. Secondary antibody alone showed no signal (data not shown). Interestingly, some nuclei in normal colorectal mucosa and stromal cells as well as nuclei in lymphocytes in tonsillar tissue also showed TS expression (data not shown). These results provide the strongest immunohistochemical evidence to date that TS is present in the nuclei of some normal and cancer cells.
Putative Nuclear Export Sequence in the TS Gene
The cellular localization of proteins can be controlled by the presence of nuclear localization sequences and/or NES. Human TS appears to have a 4-2-1 leucinerich amino acid sequence motif at position 257-269 that is similar to the NES found in other species (Figure 7A) (Zhang and Xiong 2001). To test whether it might be functional in HeLa cells, we prepared a construct with the putative TS-NES at the C terminus of Green Fluorescent Protein (GFP-NES). However, both the vector alone and GFP-NES showed a similar GFP distribution—both nuclear and cytoplasmic (Figure 7B); similar results were seen in RKO cells (data not shown). As another approach to testing the putative TS-NES, a series of deletion mutants of TS were analyzed (Figure 8A). Wild type TS (wt TS) (data not shown) and full-length TS with an N-terminal histidine6 tag (H6TS313) were transiently transfected into HeLa cells. In both cases, a high level of TS was found in the cytoplasm, but a low level of TS was also seen in the nucleus of most transfected cells (Figure 8B). HeLa cells transfected with deletion mutant H6TS248 showed a moderately uniform distribution of TS in both the cytoplasm and the nucleus (Figure 8D). Expression of deletion mutant H6TS186 in HeLa cells and RKO cells appeared to be toxic because the number of transfected cells was very low. A punctate distribution of TS was seen throughout the few cells where staining was observed (data not shown).
Discussion
Confidence in immunohistochemistry requires that the primary antibody be well-characterized. For example, the specificity of a widely used anti-TS monoclonal antibody, TS106, was previously established by demonstrating that immunohistochemical staining for TS in a human colon carcinoma specimen was eliminated by competition with rhTS protein (Johnston et al. 1991). Several polyclonal anti-TS antibodies have been described but, in only one case (Okabe et al. 1997), has specificity for TS been demonstrated. The specificity of the anti-hTS rabbit polyclonal antibody developed in our laboratory was established previously (Haqqani et al. 1999). For the present study, we thought it prudent to also confirm the specificity of the primary anti-hTS sheep polyclonal antibody used in this work. Several complementary approaches were employed. Western blot analysis of HeLa cell extracts detected only a single band corresponding to 35 kDa human TS. Western blot analysis of HeLa-55 cells maintained in 5-FUdR detected both the ternary complex form and the free form of TS protein. A series of competition experiments using highly purified rhTS protein were carried out. The very strong free and complexed TS bands observed in Western blots of HeLa-55 cell extracts essentially disappeared when primary antibody preparations were preincubated with rhTS. Similarly, we obtained almost complete elimination of immunofluorescence signal in HeLa-55 cells and in immunohistochemical staining of colorectal cancer specimens stained with rhTS-pretreated anti-TS antibody. Taken together, these experimental findings provide a strong body of evidence supporting the specificity of the antibodies used in this study.
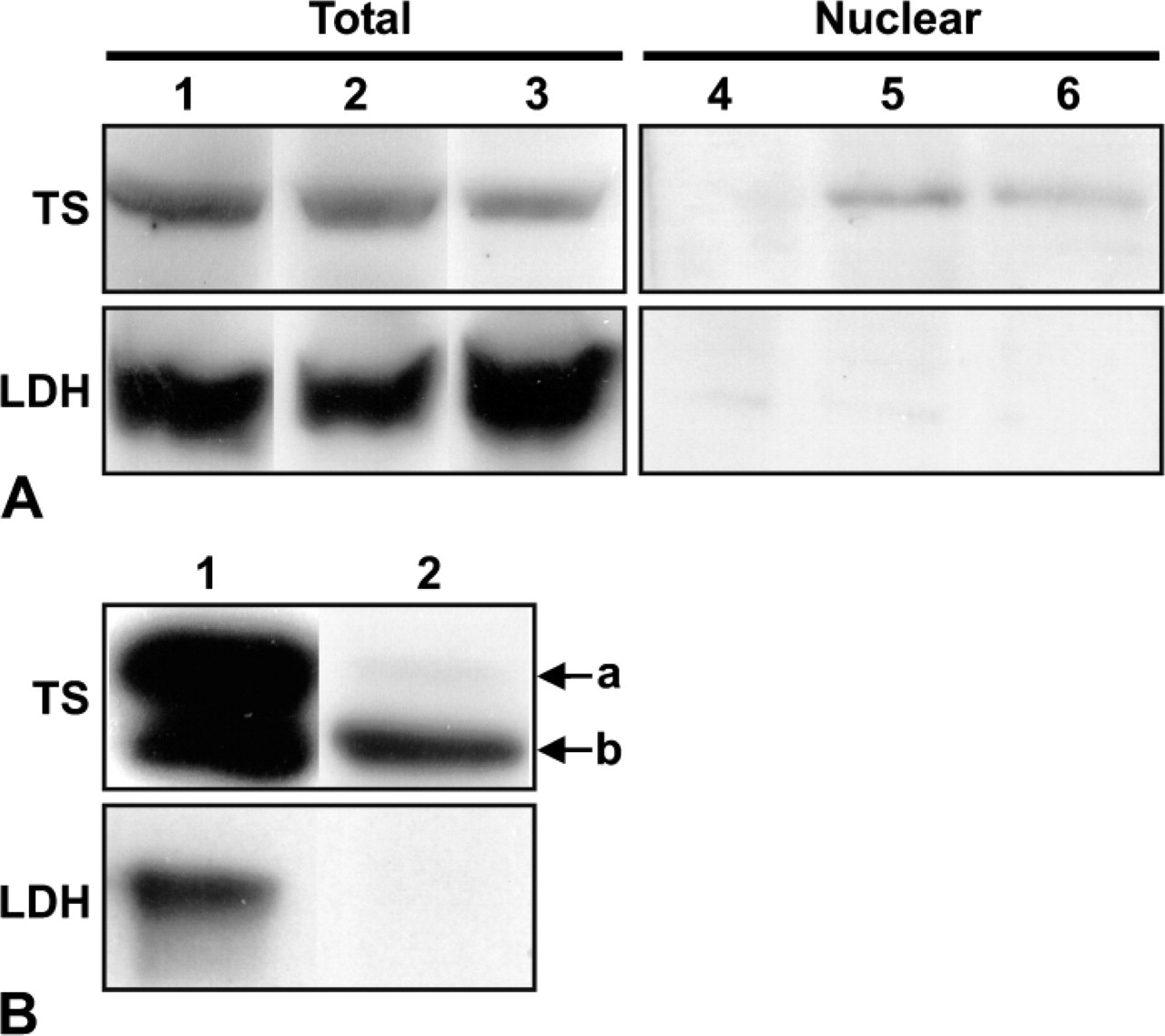
Western blot analysis for thymidylate synthase (TS) and lactate dehydrogenase (LDH) of the total cellular or nuclear extracts in several cell lines. (
TS is usually considered to be a cytoplasmic enzyme. While there have been previous observations that it may also be expressed in the nucleus of human cells (Johnston et al. 1991; Wong et al. 2001), neither of the earlier studies used a multifaceted approach to prove that low-level nuclear staining was indeed TS. Our studies are in agreement with earlier findings that low levels of TS may be present in the nucleus as well as in the cytoplasm of some cells. We used two general approaches to demonstrate the presence of nuclear TS. Confocal microscopy of HeLa-55 (a strongly overexpressing line) and RKO cells provided unambiguous evidence of TS nuclear localization. Fractionation of HeLa-55, RKO, and HCT-116 cells into nuclear and cytoplasmic fractions followed by Western blotting confirmed that a significant fraction of total cellular TS was located in the nuclear compartment. Our observations about nuclear expression of TS were not limited to cultured cells. Nuclear TS expression was also observed in both normal and cancerous colorectal tissue. Indeed, nuclear TS expression was found in germinal centers of normal tonsillar tissue. Interestingly, in tissue sections, nuclear TS was not uniformly expressed (both in normal and cancerous cells), but it was uniformly expressed in RKO colorectal cancer cells (data not shown). Among cultured cancer cell lines, only some expressed nuclear TS.
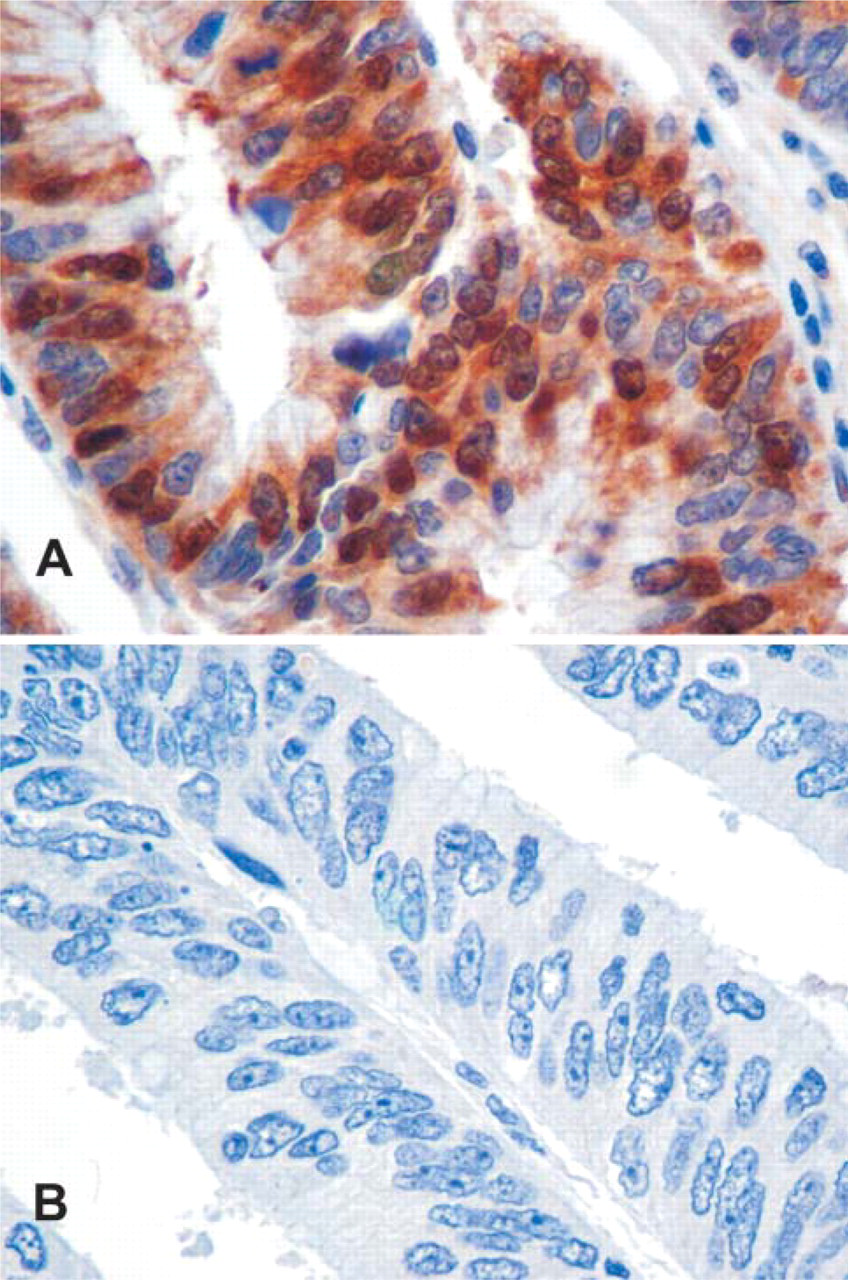
Immunohistochemical analysis for thymidylate synthase (TS) of colorectal tissue sections.
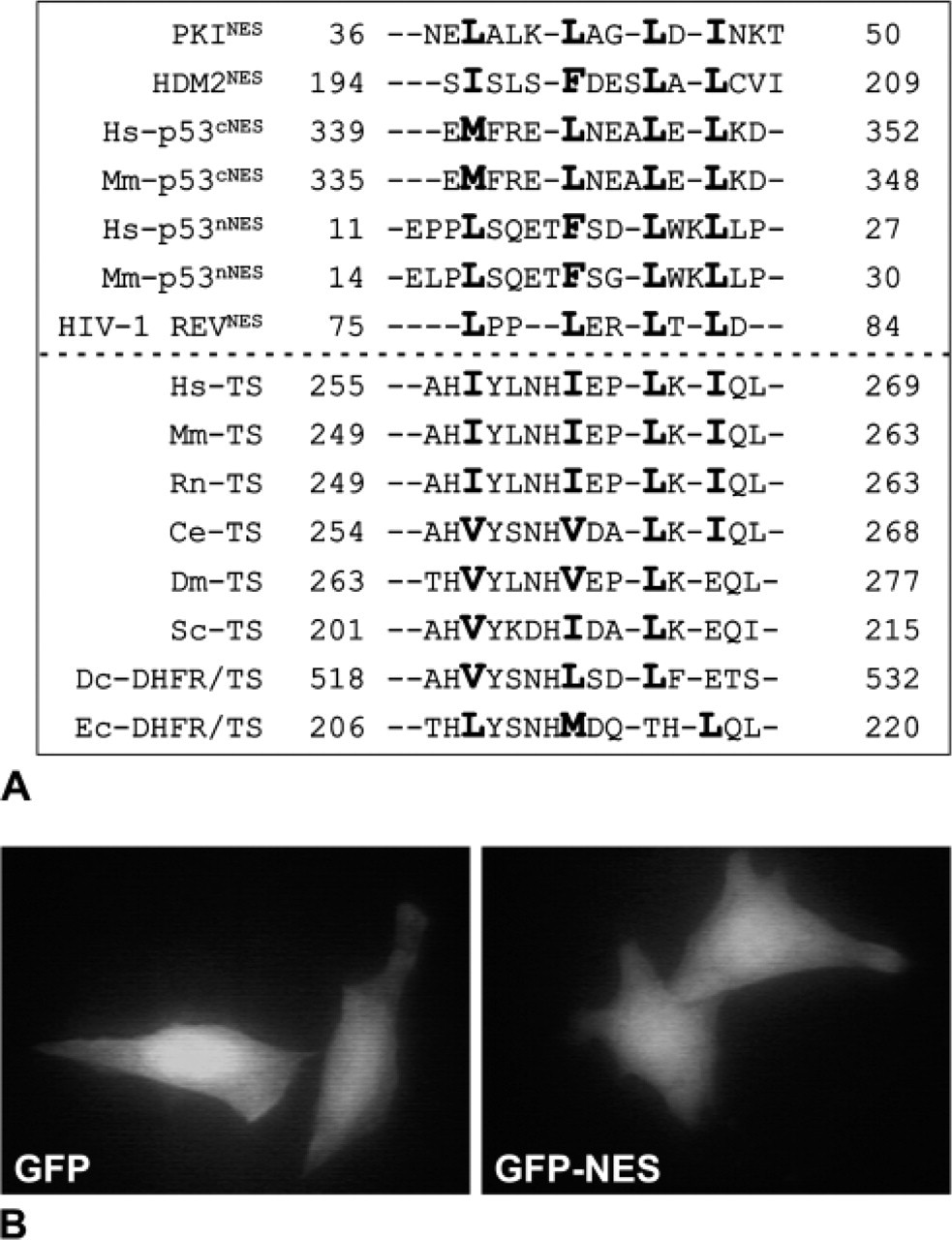
Putative leucine rich nuclear export signal (NES) of thymidylate synthase (TS). (
The biological significance of nuclear TS expression at the cellular level remains unclear. One possibility is that the presence of TS in the nucleus may be related to its reported RNA binding activity. TS protein has been reported to bind its own mRNA as well as p53 and c-myc mRNAs (Chu et al. 1995, 1999). If high levels of the nuclear forms of these RNAs were to accumulate because of slow processing into mature mRNA, then nuclear TS might be a result. At the clinical level, Wong et al. (2001) have described an unexplained association between nuclear TS expression and poorer response to 5-FU-based therapy and higher levels of TS mRNA.
Cell fractionation studies of HeLa-55 cells grown in the presence of 5-FUdR led us to an unexpected observation. TS expressed in the nucleus was limited to free TS; the ternary complex form of TS was restricted to the cytoplasmic fraction. The mechanism for this is unknown. Some possible explanations are that 5-dUMP cannot enter the nucleus, that nuclear TS is in a different configuration than cytosolic TS, or that the ternary complex form of TS cannot enter the nucleus from the cytoplasm whereas the uncomplexed form is able to. Parental HeLa cells did not show nuclear expression of TS nor did short-term culture (2 days) of these cells in 5-FUdR lead to nuclear expression of TS (data not shown). Long-term exposure to increasing concentrations of 5-FUdR, conditions used in isolating TS-overexpressing HeLa-55 cells, was required before nuclear TS expression was seen. Nuclear TS could still be detected in HeLa-55 cells grown in the absence of 5-FUdR for 5 days. Growth for several weeks in the absence of 5-FUdR was required to lower total TS levels and eliminated nuclear TS (data not shown). Transient overexpression of TS by transfection into HeLa cells led to nuclear expression of TS (Figure 8B and data not shown). These two lines of evidence suggest that strong overexpression of TS may be sufficient for TS to be present in the nucleus.
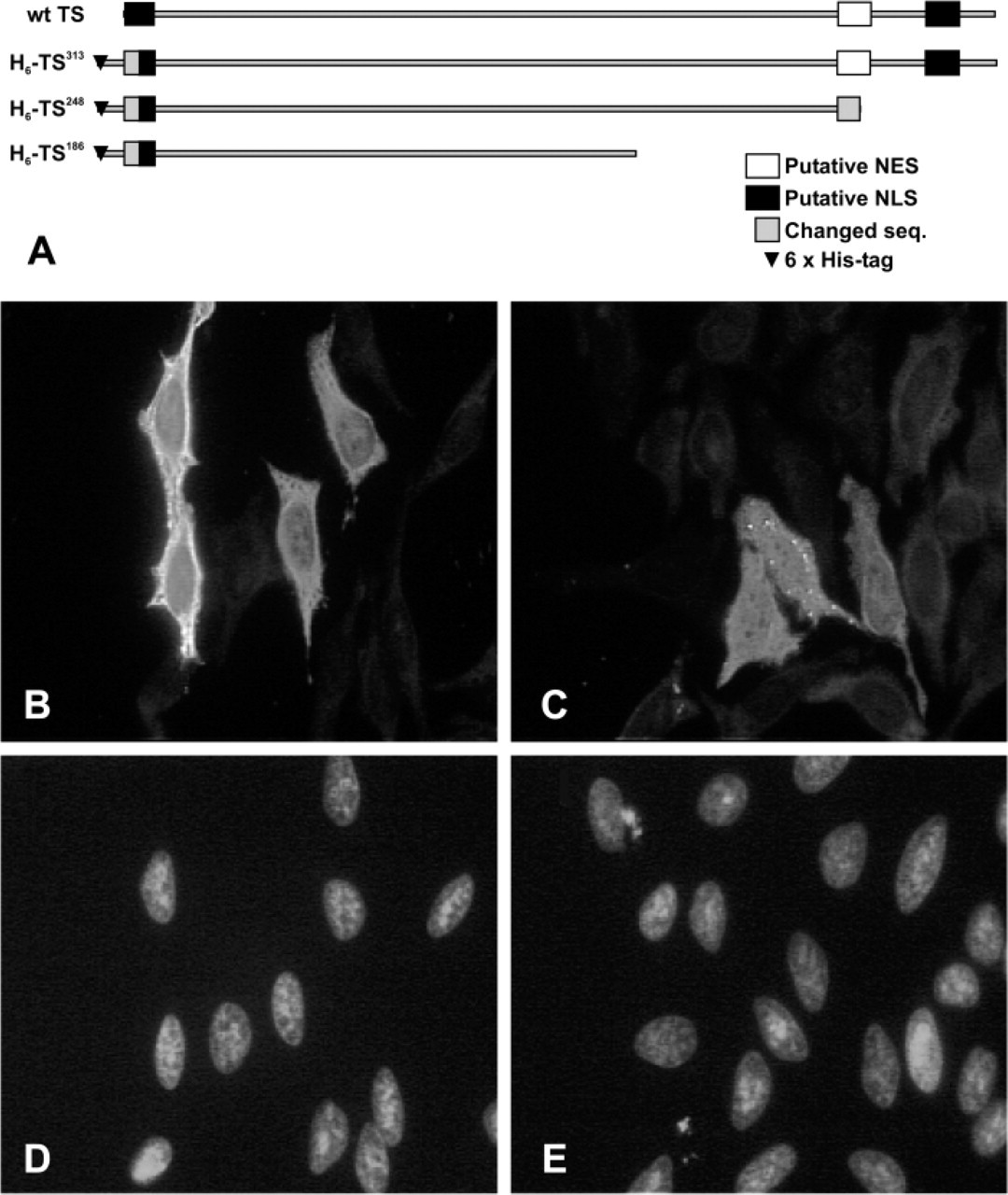
Effect on nuclear localization of deletion mutations affecting the putative thymidylate synthase (TS) nuclear export signal in transfected HeLa cells. (
Nuclear localization of proteins is governed by the presence of nuclear localization and nuclear export amino acid sequences. Although a putative NES was identified in human TS, experiments to verify its functionality were inconclusive.
In conclusion, the work presented in this paper characterizes an antibody that is available to researchers from a commercial source. While many antibodies to TS are described in the previously published literature, none were as extensively characterized as in the present report. Our work is the first to unambiguously prove, using a multifaceted approach, that the nuclear signal seen by us and others is due to TS. Small amounts of TS are present in the nucleus of some human cell lines, but not others, as well as tissue specimens. This report also documents a major difference in the cytoplasmic vs the nuclear localization of the free form vs the ternary complexed form of TS in FUdR-exposed cells.
Footnotes
Acknowledgements
We thank Dr. Jagdeep K. Sandhu for providing advice on immunofluorescence staining and Ms. Catherine Duff for providing technical assistance on the use of the confocal laser scanning microscope.