
Abstract
Ribosomal proteins are a major component of ribosomes and play critical roles in protein biosynthesis. Recently it has been shown that the ribosomal proteins also function during various cellular processes that are independent of protein biosynthesis therefore called extraribosomal functions. In this study we have, for the first time, determined the expression profile of 12 ribosomal proteins (Sa, S8, S11, S12, S18, S24, L7, L13a, L18, L28, L32, and L35a) in normal epithelia of human colorectal mucosa using immunohistochemistry (IHC) and then compared their expression patterns with those of colorectal cancer. In the normal mucosa, ribosomal proteins were largely associated with the ribosomes of mucosal epithelia, and the expression level of ribosomal proteins, except for S11 and L7 proteins, was markedly increased in associated with maturation of the mucosal cells. On the other hand, these ribosomal proteins were markedly decreased in colorectal cancer compared with the normal mucosa. By contrast, S11 and L7 ribosomal proteins were rarely associated with the ribosomes of colorectal epithlia except immature mucosal cells, whereas their expression levels were significantly enchanced in colorectal cancer cells. In addition, L7 ribosomal protien was detected in the secretory granules of the enterochromaffin cells in the colorectal mucosa and in carcinoma cells expressing chromogranin A. These results indicate that the expression of ribosomal proteins is differentially regulated not only in normal mucosa but also in carcinoma of human colorectum, and suggest an extraribosomal function of L7 ribosomal protein in neuroendocrine function.
R
A differential expression of specific ribosomal protein genes is reported in several pathological conditions (Go and Taniguchi 1998; Bevort and Leffers 2000). In particular, upregulation of transcripts for several ribosomal proteins has been shown in several malignancies, including carcinomas of the colorectum, prostate, and esophagus (Pogue-Geile et al. 1991; Vaarala et al. 1998; Wang et al. 2001). However, most of these studies are based on analyses carried out at the transcriptional level. Because the expression of ribosomal proteins is not only transcriptionally but also translationally controlled in vertebrates, analysis at the translational level is important (Mager 1988).
Colorectal mucosa and colorectal cancer have been widely utilized as a model system to investigate cell differentiation, cell growth, and carcinogenesis (Vo-gelstein et al. 1988; Augenlicht et al. 1999). However, the immunohistochemical (IHC) localization of ribosomal proteins in colorectal mucosa and cancer was largely unknown due to the limited availability of antibodies specific for human ribosomal proteins. In this study we have demonstrated the expression profile of 12 ribosomal proteins including Sa, S8, S11, S12, S18, S24, L7, L13a, L18, L28, L32, and L35a in human normal colorectal mucosa by IHC using a well-characterized antibody panel for the ribosomal proteins (Nadano et al. 2000; Nishida et al. 2002). We compared their expression patterns with those in colorectal adenocarcinoma cells. The subcellular localization of S12 and L7 ribosomal proteins has been also shown by immunoelectron microscopy.
Materials and Methods
Tissue Specimens of Colorectal Cancer
Surgical specimens were obtained from 18 primary colorectal carcinoma patients who underwent surgical resection at the Aizawa Hospital, Matsumoto, Japan from November, 2000 to March, 2001 after informed consent was obtained. These patients (12 men and 6 women, age range 51–86 years) included 13 cases of well-differentiated adenocarcinomas and five cases of moderately differentiated adenocarcinomas. The patients had not received any therapeutic agents or irradiation before surgery. Tissue samples from colorectal carcinomas and their cut end of the normal-looking mucosa were examined in this study. Histological diagnosis of colorectal cancer was made according to the World Health Organization Histologic Classification (Jass and Sobin 1989).
Light Microscopic IHC for Ribosomal Proteins
All tissue samples were fixed in 20% buffered formalin for 48 hr, embedded in paraffin, and serially sectioned at 3-μm thickness. In parallel, tissue samples from three cases were immediately frozen in OCT compound (Sakura Finetek; Torrance, CA) at −80C and frozen sections were serially sliced at 10-μm thickness and fixed in absolute ethanol for 15 min at room temperature (RT). IHC was performed by the indirect method as described previously (Ichikawa et al. 1999). Deparaffinized tissue sections and frozen sections were treated with 0.3% H2O2 in methanol and then blocked with 1% normal goat serum in Tris-buffered saline (TBS), pH 7.4. Then the tissue sections were incubated with a panel of anti-ribosomal protein antibodies against Sa, S8, S11, S12, S18, S24, L7, L13a, L18, L28, L32, and L35a at 4C overnight. These primary antibodies were produced by immunizing synthetic peptides specific for the ribosomal proteins, and the specificity of antibodies was confirmed by using immunoblots of the isolated ribosomal proteins (Nadano et al. 2000) as well as an inhibition assay using the synthetic peptides employed for immunogens (Nishida et al. 2002). In addition, rabbit polyclonal antibodies against chromogranin A, a distinct marker for enterochoromaffin cells (Ho et al. 1989), were purchased from DAKO (Carpinteria, CA). After washing with TBS, they were incubated with anti-rabbit immunoglobulins conjugated with horseradish peroxidase for 1 hr at RT. Peroxidase activity was then visualized using a 3,3'-diaminobenzidine tetrahydroxychloride (DAB)/H2O2 solution. A control experiment was performed by omitting the primary antibodies from the staining procedure, and no specific staining was found. Counterstaining was performed with hematoxylin.
Ultrastructural IHC for Ribosomal Proteins
Tissue samples from the normal colorectal mucosa were analyzed for ultrastructural IHC for ribosomal proteins S12 and L7, using a postembedding method as described before with slight modification (Hidaka et al. 2000). Fresh tissue samples obtained from three patients at surgery were immediately fixed in a 4% paraformaldehyde solution in 0.1 M phosphate buffer, pH 7.4, for 16 hr at 4C. After rinsing in the same buffer, they were dehydrated through graded alcohols and embedded in LR White resin. Semithin sections prepared from these tissue blocks were stained with toluidine blue. Ultrathin sections were then prepared from trimmed blocks and collected on single-slot copper grids. Immuno-staining using anti-S12 and -L7 ribosomal proteins was performed as follows. Ultrathin sections were placed for 1 hr on droplets of 0.1 M PBS containing 1% bovine serum albumin (BSA) at RT. These grids were then left overnight at 4C on drops of PBS containing 1% BSA and primary antibody against S12 or L7 ribosomal protein. After rinsing on drops of PBS, sections were exposed for 4 hr at RT to second antibody, anti-rabbit IgG labeled with colloidal gold (15 nm in diameter) purchased from Polysciences (Warrington, PA).
After rinsing on drops of PBS and distilled water at RT, these grids were dried and then stained with uranyl acetate and lead citrate. Ultrathin sections thus prepared were observed using a transmission electron microscope JEM1010 (JEOL) at 80-kV accelerating voltage. Negative controls were carried out by omitting primary antibody from the procedure, and no specific binding was found.
Evaluation
To evaluate the results of the IHC staining for ribosomal proteins, a scoring system based on criteria described previously was used (Basolo et al. 1996; Ichikawa et al. 1999). Briefly, the staining intensity was scored from 0 to 3 (0, absent; 1, weak; 2, moderate; 3, strong). In addition, the proportion of ribosomal protein-positive cells relative to all epithelial cells in the tissue specimens was graded from 0 to 3 (0, <5%; 1, 6–10%; 2, 11-50%; 3, >51%). By adding the two individual scores, the total score was determined. These observations were done by scanning the entire tissue specimen at a low-power magnification (X40 or X100) and then confirmed under high-power magnification (X400).
Statistical Analysis
Statistical analysis was performed using the StatView 4.5 software package purchased from Abacus Concepts (Tucson, AZ). The Mann-Whitney non-parametric test was used to compare the staining score and categorized data, and values of p<0.05 were considered significant.
Results
Expression Profile of Ribosomal Proteins in Normal Colorectal Mucosa
To determine the expression profile of various ribosomal proteins in normal colorectal mucosa, IHC using an antibody panel for the 12 ribosomal proteins was performed.
Initially, using tissue samples derived from three patients, we found that the staining patterns of 12 ribosomal proteins detected in 20% formalin-fixed and paraffin-embedded tissue sections were basically similar to those detected in absolute ethanol-fixed frozen tissue sections. Therefore, we used formalin-fixed paraffin-embedded tissue sections for further analysis.
Among the ribosomal proteins examined, 10 of the proteins, Sa, S8, S12, S18, S24, L13a, L18, L28, L32, and L35a, exhibited a similar staining pattern. They were strongly expressed in the supranuclear region of colorectal epithelia, including columnar and goblet cells, but were barely detectable in the nuclei of the epithelial cells (Figures 1A and 1C). Interestingly, these ribosomal proteins were expressed more abundantly in mature epithelial cells of the upper crypts rather than in immature epithelial cells of the lower crypts, suggesting that the biosynthesis of these ribosomal proteins is significantly enhanced in association with maturation of the mucosal epithelia.
Because the above 10 ribosomal proteins exhibited similar localization patterns, we selected S12 riboso-mal protein to determine its subcellular localization using immunoelectron microscopy. As shown in Figure 1G, gold particles indicating the presence of S12 ribosomal protein were largely associated with rough endoplasmic reticulum (rER) of the colorectal epithelia. In addition, gold particles were scattered in the cytoplasm around the rER. A small number of gold particles were also found in the nuclear matrix of the mucosal epithelia (Figure 1H). These IHC results establish that these antibodies specifically recognize not only the ribosomes associated with the rER but also free ribosomes in cytoplasm, as well as preribosomes in the nucleus.
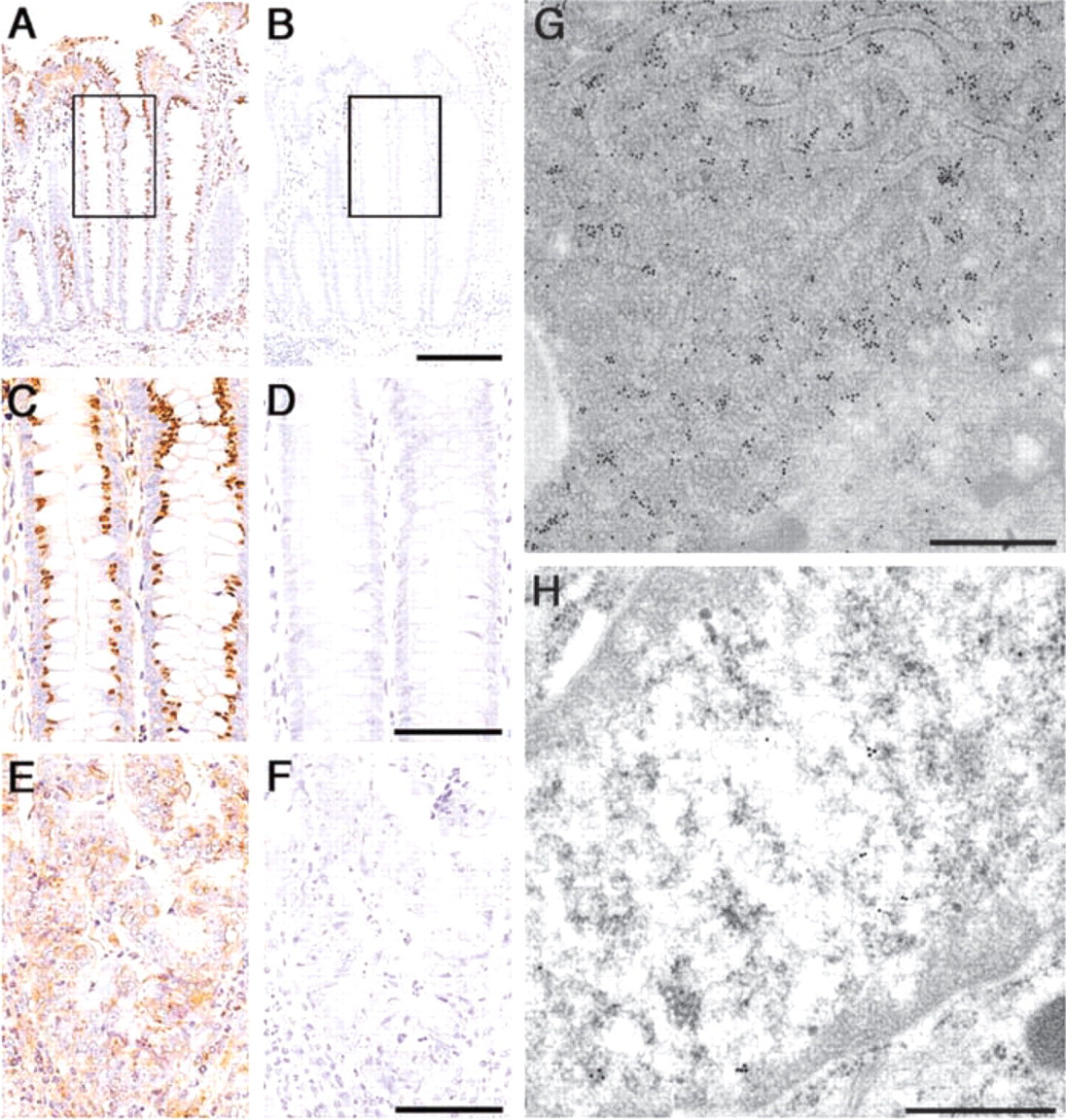
Expression of S12 ribosomal protein in normal mucosa and in carcinoma of the colon. (
On the other hand, distinct expression patterns were found in two ribosomal proteins, S11 and L7. S11 ribosomal protein was barely expressed in the ribosomes of colorectal epithelia, except for a few immature mucosal cells located in the crypt base (Figures 2A and 2C). L7 ribosomal protein was also modestly expressed in the normal colorectal epithelia, except for a small number of immature epithelia located in the crypt base, like S11 ribosomal protein (Figures 3A and 3D). Surprisingly, significant amounts of L7 ribosomal protein were frequently found in the cytoplasm of enterochromaffin cells expressing chromogranin A (compare Figures 3D and 3E). However, enterochromaffin cells that express chromogranin A did not always express L7 ribosomal protein. Subcellular localization of L7 ribosomal protein in the enterochromaffin cells was further examined using immunoelectron microscopy, and it was clear that L7 ribosomal protein was detected in the secretory granules of the enterochromaffin cells (Figures 3J and 3K). However, the amounts of L7 ribosomal proteins in the secretory granules significantly differed from granule to granule, and secretory granules that barely contained L7 ribosomal protein were also found (Figure 3K). These results suggest that L7 protein plays a role in neuroendocrine function of the enterochromaffin cells.
Expression of Ribosomal Proteins in Colorectal Cancer
Expression of ribosomal proteins in colorectal cancer was examined using the same antibody panel for ribosomal proteins and we compared their expression with that of the normal mucosa (Figure 4). In 17 (94.4%) of 18 patients with colorectal carcinoma, the expression level of 10 ribosomal proteins, Sa, S8, S12, S18, S24, L13a, L18, L28, L32, and L35a, in the supranuclear region of carcinoma cells was weaker than that in the normal colorectal mucosa (Figure 4), and particularly significant differences (p<0.05) were obtained in S12, S24, L32, and L35a ribosomal proteins (compare Figures 1C and 1E). When the expression level of these ribosomal proteins was analyzed according to the histological classification of colorectal cancer, four ribosomal proteins, S18, S24, L18, and L32, were much more strongly expressed in well-differentiated adenocarcinoma than in moderately differentiated adenocarcinoma. However, no significant difference was obtained (data not shown).
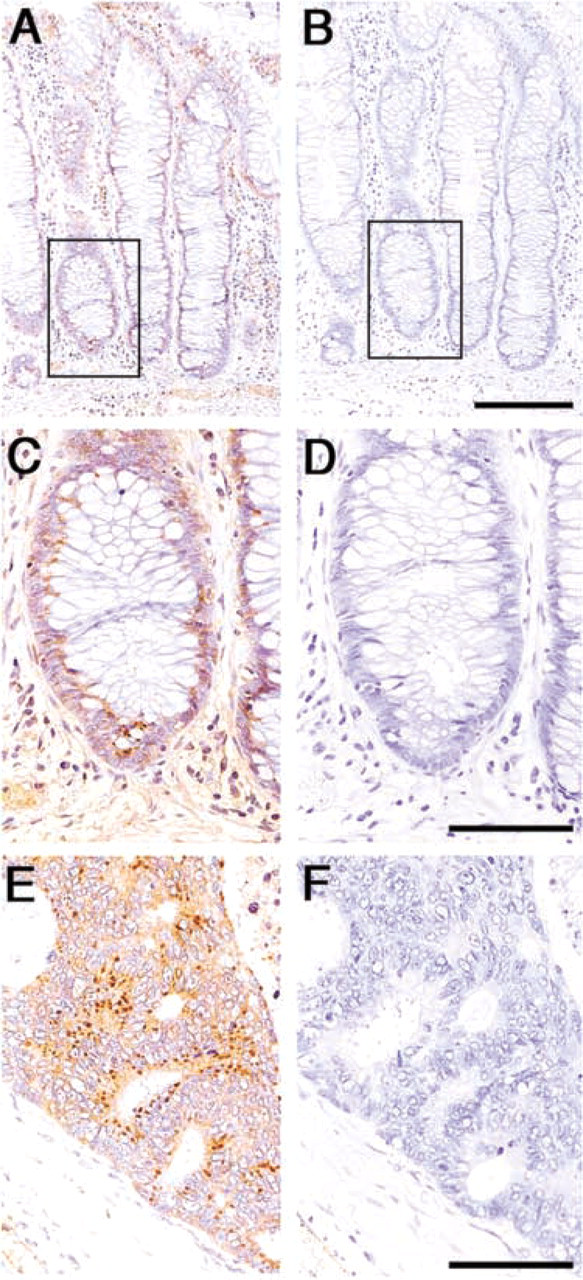
Expression of S11 ribosomal protein in the normal mucosa and in carcinoma of the colon. (
In contrast, the expression level of S11 and L7 ribosomal proteins in the supranuclear region of carcinoma cells was significantly increased in the carcinoma compared with the normal mucosa with statistical differences (p<0.05) (Figures 2E and 4). Although no significant differences were obtained, these two ribosomal proteins tended to be expressed more abundantly in moderately differentiated adenocarcinoma than in well-differentiated adenocarcinoma (data not shown).
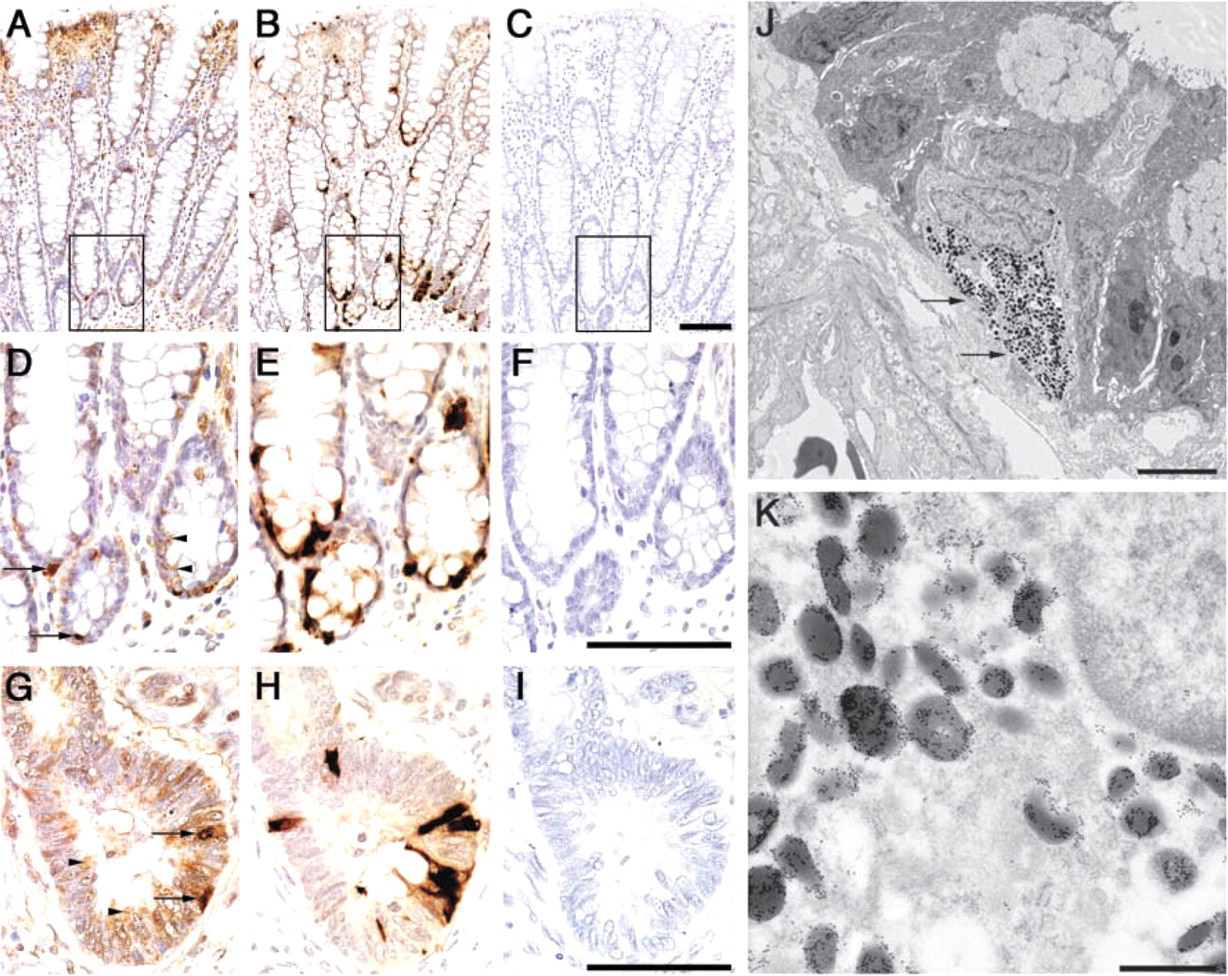
Expression of L7 ribosomal protein in the normal mucosa and in carcinoma. (
In nine (50%) of 18 patients examined, chromogranin A was detected in a small number of carcinoma cells, indicating the neuroendocrine differentiation of the carcinoma cells (Figure 3H). Interestingly, some of the carcinoma cells also appeared to express L7 ribosomal protein (compare Figures 3G and 3H), again suggesting an extraribosomal function of L7 in neuroendocrine function.
Discussion
In humans, 80 ribosomal proteins for males and 79 ribosomal proteins for females have been identified and characterized at the molecular level (Wool et al. 1996; Warner and Nierras 1998). However, their precise function and tissue expression have not been fully understood. In particular, systematic analysis of the immunolocalization of these proteins in various human tissues has not been reported. In the present study we have, for the first time, demonstrated the expression profile of 12 ribosomal proteins in normal human colorectal mucosa and compared their expression patterns with those of colorectal cancer cells using an antibody panel for the ribosomal proteins. A battery of polyclonal antibodies against 26 human ribosomal proteins was originally developed for comprehensive analysis of these proteins (Nadano et al. 2000). In the present study, we selected 12 of 26 antibodies applicable to IHC analysis. We found that these antibodies clearly identify ribosomal proteins, not only in ethanol-fixed frozen tissue sections but also in formalin-fixed and paraffin-embedded tissue sections, without any antigen retrieval procedures. Therefore, these antibodies are applicable to investigations of the expression pattern of ribosomal proteins in various human tissues under normal as well as pathological conditions using archival paraffin blocks fixed with 20% buffered formalin.
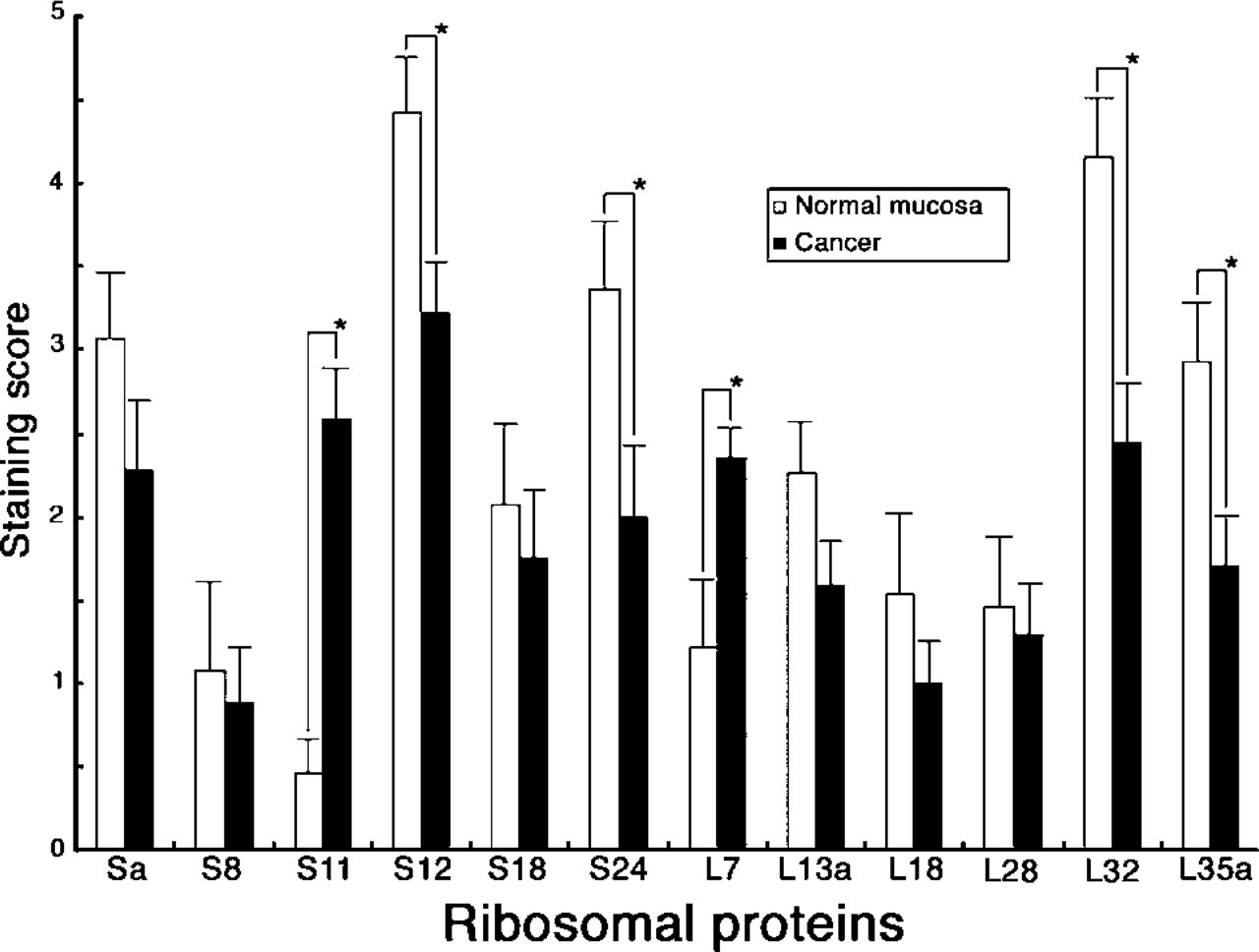
Comparison of the expression level of 12 ribosomal proteins in the ribosomes of normal colorectal mucosa and carcinoma. Bar indicates mean ± SE. Significant differences (p<0.05) between the normal mucosa and carcinoma are indicated as asterisks (∗).
In the present study we have clearly demonstrated that all of the ribosomal proteins examined are largely expressed in the ribosomes associated with the rER and in free ribosomes in the cytoplasm. Expression of the ribosomal proteins in the nucleus was not so conspicuous at the light microscopic level, but detectable amounts of ribosomal proteins in the nuclear matrix were shown by immunoelectron microscopy. These localization patterns of the ribosomal protein appear to be reasonable, since it is known that ribosomal proteins primarily synthesized in the cytoplasm are imported to the nucleolus, where they associate with newly synthesized ribosomal RNA, thus forming the preribosomes. These preribosomes are then exported back to the cytoplasm (Aitchison and Rout 2000). Recently, it was shown that the translation process occurs in the nucleus of eukaryotes (Iborra et al. 2001). Thus, the presence of S12 ribosomal protein in the nucleus of colorectal epithelia may reflect such an intranuclear translation.
We have clearly shown that the expression level of ribosomal proteins, including Sa, S8, S12, S18, S24, L13a, L18, L28, L32, and L35a, was significantly increased in association with the maturation of mucosal epithelia, including goblet cells and columnar cells. In addition, these ribosomal proteins were markedly decreased in colorectal cancer compared with normal mucosa. Seemingly these results are paradoxical, because it has been generally accepted that ribosomal proteins are abundantly transcribed in actively proliferating cells, such as cancer cells. However, a recent study using DNA microarray analysis clearly demonstrates that many ribosomal proteins, such as S8, S24, and L32, are much more abundantly expressed in differentiated ovarian tumor cells than in less-differentiated ones (Welsh et al. 2000). The IHC results presented in this study provide further evidence that, in human colorectum, biosynthesis of many ribosomal proteins, such as Sa, S8, S12, S18, S24, L13a, L18, L28, L32, and L35a, is downregulated in actively proliferating cells such as immature mucosal epithelia and cancer cells.
On the other hand, S11 and L7 ribosomal proteins appeared to be barely expressed in the ribosomes in normal mucosal epithelia except for immature mucosal cells, whereas their expression levels were significantly enhanced in carcinoma cells, making a clear contrast to the other 10 ribosomal proteins examined. Recently, S11 was shown to be specifically downregulated in apoptotic breast carcinoma MCF-7 cells induced by staurosporine (Nadano et al. 2001). Therefore, the present result suggests that S11 ribosomal protein expressed in the colorectal cancer cells may inhibit apoptosis of cancer cells. In fact, Tsujitani et al. (1996) demonstrated that apoptosis is inhibited in colorectal cancer. It is of great importance to determine if S11 and/or L7 ribosomal proteins play a role for the inhibition of apoptosis in colorectal cancer.
Finally, the present study has clearly shown that L7 ribosomal protein is expressed in the secretory granules of the enterochromaffin cells in normal colorectal mucosa and in carcinoma cells expressing chromogranin A, suggesting an extraribosomal function of L7 in neuroendocrine function. Recently, various extraribosomal functions of L7 ribosomal protein have been reported. For example, L7 is shown to be a co-regulator of the vitamin D receptor-retinoid × receptor-mediated trans-activation of genes (Berghofer-Hochheimer et al. 1998). In addition, L7 ribosomal protein also serves as a major autoantigen for systemic rheumatic disease (von Mikecz et al. 1995). However, the extraribosomal function of L7 ribosomal protein in the human colorectum has not been reported. Therefore, future study will be required to determine the role of L7 ribosomal protein in enterochromaffin cells as well as in carcinoma cells showing neuroendocrine characteristics.