
Abstract
Matrix metalloproteinase-7 (MMP-7) is upregulated during carcinogenesis and its expression correlates with metastasis of human endometrial and gastrointestinal carcinomas. In the present study, we have developed a new method to localize the activity of MMP-7 within tissues. Polyethylene terephthalate films were uniformly coated with crosslinked carboxymethylated transferrin (CCm-Tf) as a substrate and incubated with frozen tissue sections mounted on the films. CCm-Tf on the films was degraded selectively by MMP-7, but showed little or no susceptibility to MMP-1, -2, -3, -9, or -13; MT1-MMP; MT3-MMP; or ADAMTS4. Although some serine proteinases such as elastase also digested CCm-Tf, CCm-Tf films impregnated with serine proteinase inhibitors prevented the digestion. When frozen sections of human endometrial carcinoma and lung carcinoma tissues were incubated on CCm-Tf films or those treated with proteinase inhibitors, the activity was detected in the carcinoma cell nests, where MMP-7 was immunolocalized. The present in situ zymography using CCm-Tf may be a useful method to analyze the functions of MMP-7 in pathophysiological conditions.
Keywords
We have previously developed gelatin-based film in situ zymography (FIZ-GN), which uses uniform thin layer of crosslinked gelatin on polyethylene terephthalate support films. FIZ-GN was applied to many tissue samples and detected gelatinolytic activity within tissues under various pathophysiological conditions, such as cancers of the brain (Nakada et al. 1999), thyroid (Nakamura et al. 1999), oral cavity (Shimada et al. 2000), esophagus (Koyama et al. 2000), liver (Kaneyoshi et al. 2001) and kidney (Kamiya et al. 2003), and nonneoplastic diseases including rheumatoid arthritis (Yamanaka et al. 2000), cardiovascular diseases (Nishikawa et al. 2003), autoimmune renal disease (Alport syndrome) (Rao et al. 2003), and ultraviolet damage of skin (Inomata et al. 2003). This method was suitable to localize gelatinolytic activities (i.e., mainly MMP-2 and MMP-9, in the tissues) and used to analyze the effect of synthetic MMP inhibitors within cancer tissues (Ikeda et al. 2000; Wada et al. 2003; Yamamoto et al. 2003). On the other hand, similar in situ zymography using the quenched fluorogenic substrate DQ-gelatin has been developed (Mook et al. 2003). The method is considered to be superior in the sensitivity and quantification for the gelatinolytic activity of MMP-2 and MMP-9, because fluorescence is generated by cleavage of DQ-gelatin at sites of gelatinolytic activity (Frederiks and Mook 2004). However, there are so far no such methods to localize MMP-7 activity within tissues.
In the present study, we attempted to develop a new in situ zymography for MMP-7 by preparing a suitable substrate of MMP-7. Because most of the MMP-7 substrates, including type IV collagen, gelatin, fibronectin, laminin, aggrecan (Imai et al. 1995), entactin (Sires et al. 1993), and tenascin (Imai et al. 1994), are also degraded by other MMPs, and some substrates are difficult to obtain in a large scale, we focused on carboxy-methylated transferrin (Cm-Tf), which was originally developed as a substrate of MMP-3 (Okada et al. 1986), but preferably digested by MMP-7 (Imai et al. 1995). We have finally succeeded in developing film in situ zymography using crosslinked Cm-Tf (CCm-Tf) substrate to localize MMP-7 activity in human endometrial carcinoma and lung carcinoma tissues.
Materials and Methods
Substrate and Film Preparations
Bovine serum transferrin (Vital Products Inc.; Boynton Beach, FL) was carboxymethylated using nonradioactive iodoacetic acid as previously described (Nagase 1995). Briefly, transferrin was dissolved in 50 mM Tris-HCl buffer (pH 8.6) containing 10 mM EDTA. Dithiothreitol was added to the solution and stood for 2 hr at room temperature to cleave disulfide bonds. Then, excess amount of iodoacetic acid was added under an alkaline condition for carboxymethylation of thiol groups. Excess reagents were removed from Cm-Tf by dialysis against distilled water. Crosslinking of Cm-Tf was conducted by using a crosslinking agent, 1,2-bis (vinyl-sulfonylacetamide) ethane. The level of crosslinking was controlled by amounts of the crosslinking agent. To make the adequately crosslinked CCm-Tf films, the aqueous Cm-Tf solution containing 1.6% crosslinking agent was coated on polyethylene terephthalate support films and heat-dried to crosslink the substrate layer. The concentration of cross-linking agent was decreased to 0.8% for preparation of the weakly crosslinked CCm-Tf films. CCm-Tf-coated films had 2.5-μm thick CCm-Tf layer. As previously reported (Iwata et al. 2001), gelatin films (FIZ-GN) were also prepared by coating gelatin solution containing crosslinking agent onto polyethylene terephthalate support films to form a 7-μm thick gelatin layer.
Preparations of Proteinase Solutions
MMP-1, -2, -3, -7, -9, and -13; membrane-type 1 MMP (MT1-MMP = MMP-14); MT3-MMP (MMP-16); and ADAMTS4 (aggrecanase-1) were purified according to previous methods (Okada et al. 1986; Okada et al. 1990; Okada et al. 1992; Imai et al. 1995; Knauper et al. 1996; Ohuchi et al. 1997; Shimada et al. 1999; Nakamura et al. 2000). These MMPs, except for MT1-MMP, MT3-MMP, and ADAMTS4, were activated by incubation with p-aminophenylmercuric acetate, and their concentrations were titrated using tissue inhibitors of metalloproteinase-1 or tissue inhibitors of metalloproteinase-2. Human liver cathepsin B, human liver cathepsin D, bovine kidney cathepsin H, and human liver cathepsin L (Calbiochem; San Diego, CA) were adjusted to 1.6 U/ml with 20 mM acetate buffer (pH 5.0) containing 1 mM EDTA and 200 mM NaCl. Bovine pancreas chymotrypsin (Biogenesis; Dorset, UK), human plasmin (American Diagnostica Inc.; Stamford, CT), porcine pancreas elastase (Sigma-Aldrich Japan; Tokyo, Japan), trypsin (Sigma-Aldrich Japan) and human neutrophil cathepsin G (Cortex Biochem; San Leandro, CA) were adjusted to 0.3, 0.5, 0.2, 8, and 2.7 U/ml with 50 mM Tris-HCl buffer (pH 7.5) containing 5 mM CaCl2, respectively.
Specificity Test of CCm-Tf Films to Proteinases
Specificity of CCm-Tf films against metalloproteinases was examined by incubation with 0.1 μM MMP-1, -2, -3, -7, -9, and -13; MT1-MMP; MT3-MMP; and ADAMTS4 in 50 mM Tris-HCl buffer (pH 7.5) containing 150 mM NaCl, 10 mM CaCl2 and 0.05% Brij-35. Two μl of each enzyme solution was spotted on the surface of CCm-Tf and FIZ-GN films. The films were incubated at 37C for 24 hr in a moist chamber, stained with staining solution containing 0.3% Biebrich Scarlet (Sigma-Aldrich Japan; Tokyo, Japan), 3.3% trichloroacetic acid and 50% ethanol in water for 6 min and then destained with water for 10 min. Specificity of CCm-Tf films against cathepsins and other serine proteinases was examined in a similar way.
Pretreatment of CCm-Tf Films with Proteinase Inhibitors
To inhibit serine proteinases (trypsin, chymotrypsin, and elastase), CCm-Tf films were pretreated with a solution of 100 μg/ml aprotinin (Wako Pure Chemical; Osaka, Japan) and 1.5 mg/ml elastatinal (Peptide Institute Inc.; Osaka, Japan) in water before use. For inhibition of MMPs or both MMPs and serine proteinases, the films were pretreated with 19.8 mg/ml 1,10-phenanthroline monohydrate (Sigma-Aldrich Japan), a mixture of 100 μg/ml aprotinin and 1.5 mg/ml elastatinal, or a mixture of 19.8 mg/ml 1,10-phenanthroline monohydrate, 100 μg/ml aprotinin, and 1.5 mg/ml elastatinal in 50% ethanol. A portion of the inhibitor solutions (125 μl) was put on a glass slide, and the coated side of a CCm-Tf film was applied to it. After incubation for 10 min, the CCm-Tf layer became fully swollen with the inhibitor solution. Then, the films were peeled off from the glass slide and dried spontaneously by holding them vertical. The pretreated films were stored in a refrigerator until use.
To evaluate the proteinase specificity of inhibitor-treated films, 2 μl of 0.1 μM active MMP-7, 0.2 U/ml porcine pancreas elastase, 8 U/ml bovine pancreas trypsin, and 2.7 U/ml cathepsin G in 50 mM Tris-HCl buffer (pH 7.5) containing 150 mM NaCl and 5 mM CaCl2 were spotted on the surface of CCm-Tf films, incubated at 37C for 20 hr and stained as described previously.
Reproducible Lysis of CCm-Tf Using Frozen Sections of MMP-7 Solution
Active MMP-7 was mixed with 20% aqueous polyvinylpyrrolidone solution at concentrations ranging from 3 to 400 μg/ml, snap-frozen in liquid nitrogen, and stored at - 80C until used. The frozen sections (14 μm thick) were prepared with a cryostat microtome and mounted on CCm-Tf films. The films with sections were incubated at 37C for 24 hr in a moist chamber and then stained with staining solution for 4 min. An image of each film was taken by CCD camera and converted to monochrome using Adobe Photoshop (Adobe System Inc.; San Jose, CA). Then, the average optical density within the degradation spots was measured using Image-Gauge (Fuji Photo Film; Tokyo, Japan).
In Situ Zymography for Frozen Sections of Human Carcinoma Tissues
Tissues of human endometrial adenocarcinoma (15 cases) and non-small-cell lung adenocarcinoma (five cases) were obtained from patients who underwent surgical resection at the Keio University Hospital, Tokyo, Japan. Written informed consent was obtained from each patient for the experimental use of the tissues. The samples were embedded without fixation in Tissue-Tek OCT compound (Sakura Finechemical Co. Ltd.; Tokyo, Japan), snap-frozen in liquid nitrogen, and stored at - 80C until use. The serial frozen sections (6 μm thick) were prepared with a cryostat microtome and mounted on CCm-Tf films, which were prepared by coating with the aqueous Cm-Tf solution containing 1.6% crosslinking on polyethylene terephthalate support films and heat drying to crosslink the substrate layer as described previously. Immediately after mounting the sections on the films, the films were incubated at 37C for 8–16 hr in a moist chamber and then stained with staining solution containing 0.3% Biebrich Scarlet for 4 min. After washing for 10 min with water, the films were treated with Mayer's hematoxylin to counterstain nuclei for 2 min, washed for 10 min with water, and kept in 20% glycerol for 5 min. After drying the films spontaneously by keeping them vertical, they were cover-slipped using Tissue-Tek SCA cover slipping film (Sakura Finetek Japan; Tokyo, Japan) with a drop of xylene and imaged under optical microscope. The CCm-Tf substrate in contact with active MMP-7 within the tissue sections was digested, and thus zones of MMP-7 activity were negatively stained. Serial frozen sections were also subjected to in situ zymography using CCm-Tf films impregnated with 1,10-phenanthroline monohydrate, a mixture of aprotinin and elastatinal or a mixture of 1,10-phenanthroline monohydrate, aprotinin, and elastatinal.
Immunohistochemistry of MMP-7
Serial frozen sections were immunostained with monoclonal antibody against MMP-7 (141-7B2; Daiichi Fine Chemical Co. Ltd.; Takaoka, Japan) or nonimmune mouse IgG according to our previous method (Ueno et al. 1999). Briefly, the sections were mounted on 3-aminopropyl-triethoxysilane-coated slides and fixed with 4% paraformaldehyde. After blocking endogenous peroxidase and nonspecific binding with 0.3% H2O2 and 10% normal horse serum, the slides were reacted with 4 μg/ml anti-MMP-7 antibody or 4 μg/ml nonimmune mouse IgG and then with an avidin-biotin-peroxidase complex (DAKO; Glostrup, Denmark). Color was developed with 0.03% 3,3'-diaminobenzidine tetrahydrochloride in 50 mM Tris-HCl buffer, pH 7.6 containing 0.006% H2O2. Counterstaining was performed with hematoxylin. Serial sections were also stained with hematoxylin and eosin.
Results
Effect of Crosslinking of Cm-Tf on Susceptibility to MMPs and Trypsin
Without crosslinking, Cm-Tf frequently came off from the coated films during staining and destaining procedures. Thus we crosslinked Cm-Tf for the experiment. Trypsin, MMP-3, and MMP-7 degraded weakly crosslinked Cm-Tf, whereas MMP-2 and MMP-9 showed negligible activity (Figure 1A). However, when Cm-Tf was properly crosslinked with 1.6% crosslinking agent, activities of trypsin and MMP-3 were greatly decreased, and only MMP-7 showed definite digestion (Figure 1B). On the other hand, excessive crosslinking resulted in a decrease in the sensitivity even for MMP-7 (data not shown). Thus we could obtain the maximal sensitivity for MMP-7 by controlling the degree of crosslinking.
Specificity of CCm-Tf Films to Proteinases
MMP specificity of CCm-Tf films was compared with that of FIZ-GN films by incubation of the films with the solutions of MMP-1, -2, -3, -7, -9, and -13; MT1-MMP (MMP-14); MT3-MMP (MMP-16); and ADAMTS4. As shown in Figure 2A, MMP-1, -2, -3, -7, -9, and -13 showed strong digestion spots on FIZ-GN films, whereas MT1-MMP, MT3-MMP, and ADAMTS4 had only weak or negligible spots. Digestion of gelatin was also obtained with cathepsins B, G, H, and L (Figure 2A) and other serine proteinases (trypsin, chymotrypsin, elastase, and plasmin) (data not shown). These data indicate that FIZ-GN is susceptible to many proteolytic enzymes. On the other hand, although MMP-7 had a clear digestion spot on CCm-Tf films, MMP-1, -2, -9, and -13; MT1-MMP; MT3-MMP; and ADAMTS4 showed negligible or no activity (Figure 2B). Thus CCm-Tf films are considered to be selective to MMP-7 among the MMPs. Cathepsins B, D, H, and L had no digestion on CCm-Tf films (Figure 2B), but serine proteinases including trypsin, chymotrypsin, elastase, plasmin, and cathepsin G had some degrading activity to CCm-Tf (Figure 2B for cathepsin G and data not shown for other pro-teinases). However, the activity of cathepsin G on CCm-Tf films appeared to be much weaker than that shown by FIZ-GN (Figures 2A and B).
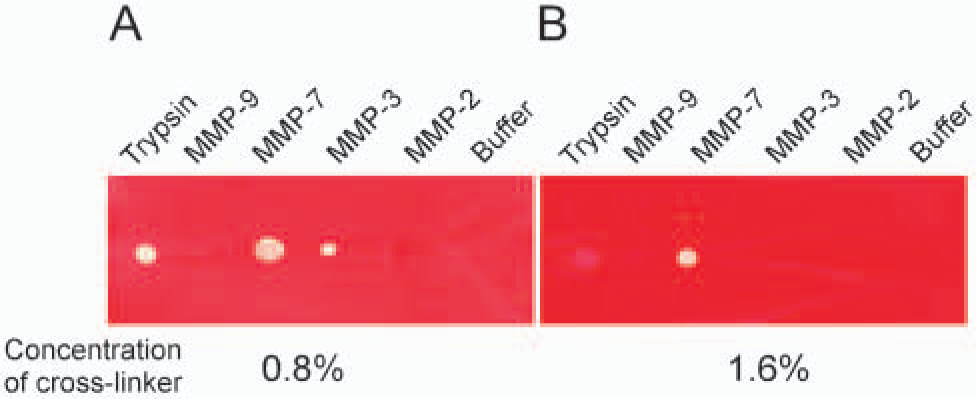
Effect of crosslinking on the specificity of carboxymethylated transferrin (Cm-Tf) films. Proteolysis of weakly crosslinked Cm-Tf with 0.8% crosslinking agent
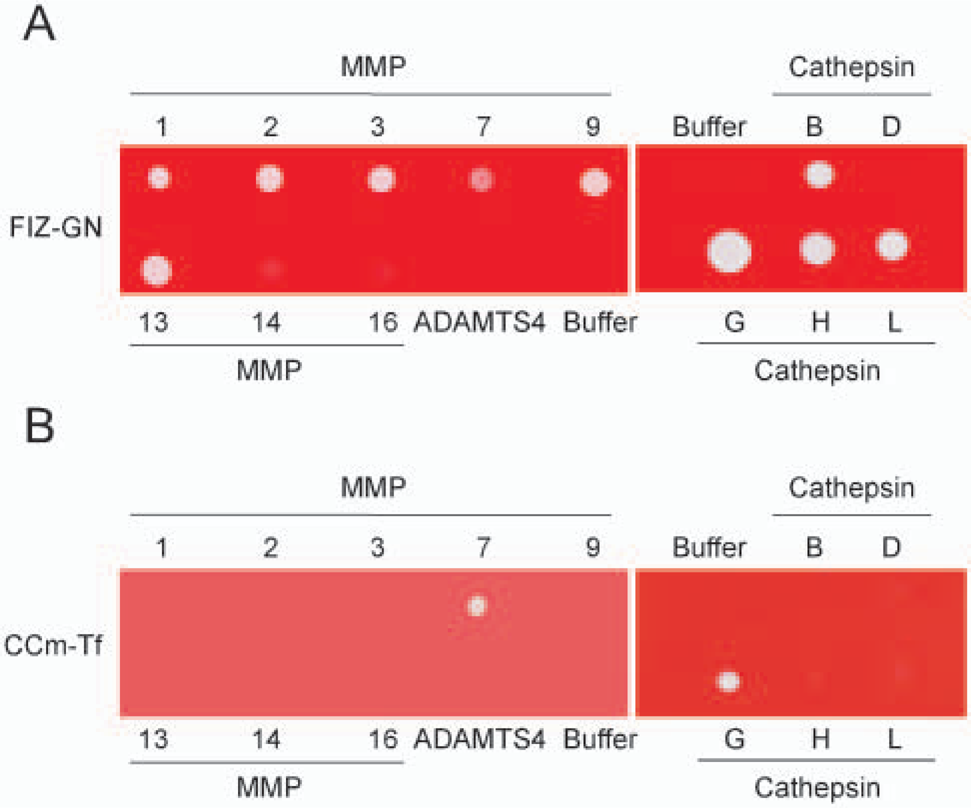
Digestion specificity of gelatin-based film in situ zymography (FIZ-GN) and crosslinked carboxymethylated transferrin (CCm-Tf) films by proteinases. Solution of each matrix metalloproteinase (MMP)-1, -2, -3, -7, -9, and -13; MT1-MMP (MMP-14); MT3-MMP (MMP-16); ADAMTS4; or cathepsin B, D, G, H, and L was spotted on the gelatin films (FIZ-GN) or CCm-Tf films, incubated, and stained as described in Materials and Methods.
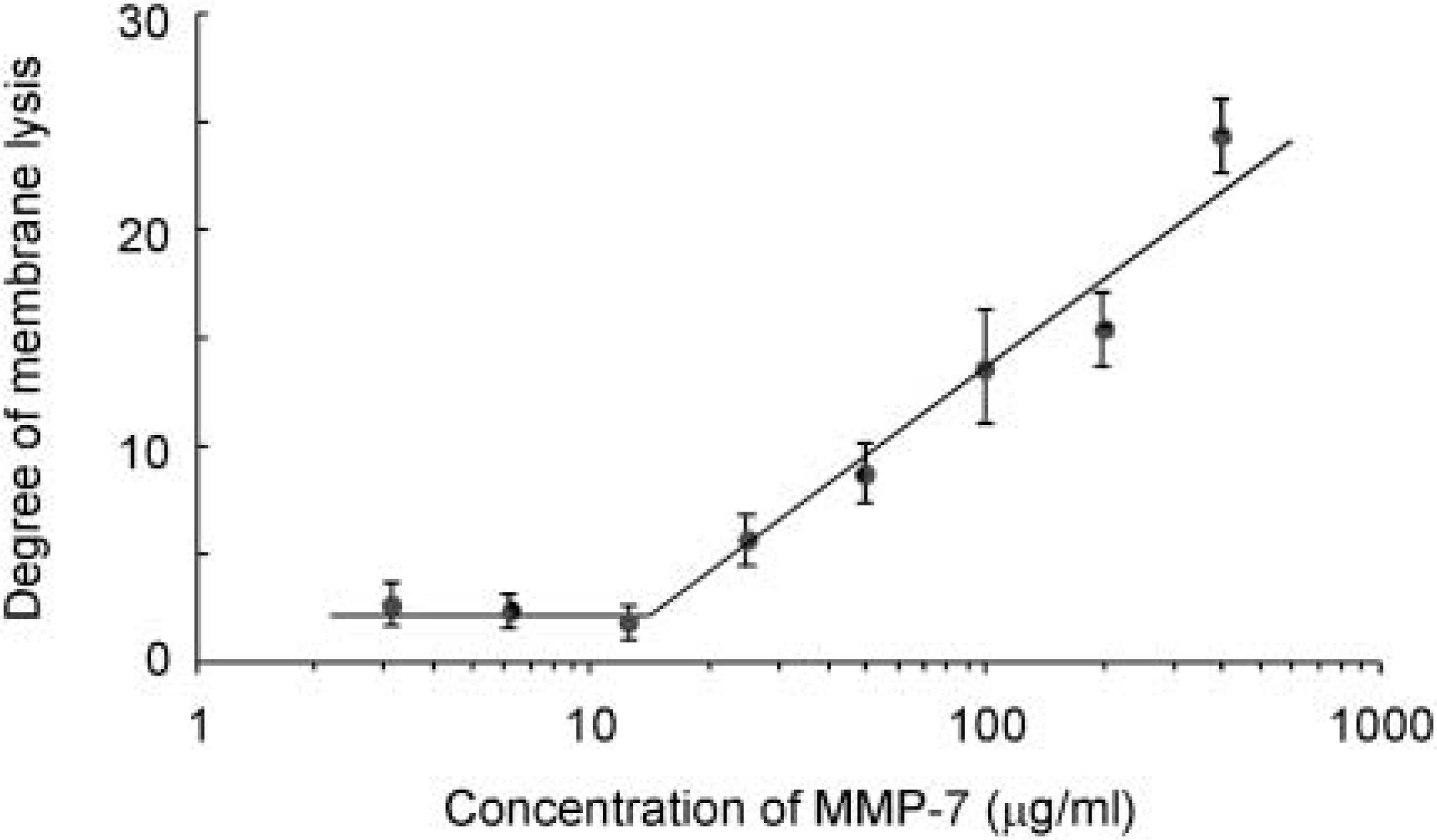
Concentration-dependent digestion of crosslinked carboxymethylated transferrin (CCm-Tf) by matrix metalloproteinase (MMP)-7. The degree of membrane lysis (arbitrary unit) was plotted as activity of MMP-7 by incubating the frozen sections of the enzyme solution with different concentrations of MMP-7 as described in Materials and Methods.

Effect of proteinase inhibitors on crosslinked carboxy-methylated transferrin (CCm-Tf) films. Proteolytic activities of elastase, matrix metalloproteinase (MMP)-7 and trypsin were examined by spotting enzyme solutions in duplicate on CCm-Tf films without inhibitor treatment
Reproducibility of Film In Situ Zymography Using CCm-Tf to MMP-7
Reproducible lysis of CCm-Tf by MMP-7 was evaluated by placing frozen sections of MMP-7 embedded in polyvinylpyrrolidone solution on CCm-Tf films. Proteolytic spots of four frozen sections were analyzed by densitometry. As shown in Figure 3, lysis level of CCm-Tf by MMP-7 increased with concentrations of MMP-7, showing a linear pattern at the range between 25 μg/ml and 400 μg/ml of MMP-7. These data demonstrate the concentration-dependent lysis with good reproducibility.
Selective Inhibition of Serine Proteinases
To reinforce the specificity of CCm-Tf films, we prepared CCm-Tf films impregnated with inhibitors. Although CCm-Tf films without inhibitor treatment were susceptible to serine proteinases (elastase, plasmin, chymotrypsin, and trypsin) and MMP-7 (Figure 4A and data not shown for plasmin or chymotrypsin), impregnation of aprotinin (an inhibitor to trypsin, chymotrypsin, and plasmin) and elastatinal (that to elastase) to CCm-Tf films almost completely prevented the digestion by these serine proteinases but not by MMP-7 (Figure 4B and data not shown for plasmin and chymotrypsin). In addition, activities of all these serine proteinases and MMP-7 were undetectable on CCm-Tf impregnated with the mixture of aprotinin, elastatinal, and 1,10-phenanthroline (a metalloproteinase inhibitor) (Figure 4C and data not shown for plasmin or chymotrypsin). However, activity of cathepsin G, another serine proteinase, was not inhibited by treatment with aprotinin, elastatinal, or 1,10-phenanthroline (data not shown). Therefore, these data suggest that the activity of MMP-7 can be differentiated from other serine proteinases except for cathepsin G by comparing the lysis patterns on the CCm-Tf films impregnated with inhibitors.
Application of Film In Situ Zymography Using CCm-Tf to Clinical Samples
We have reported that MMP-7 is highly expressed and activated in human endometrial carcinoma tissues (Ueno et al. 1999). Thus frozen sections of the carcinomas were incubated on CCm-Tf films (Figure 5A) and those impregnated with aprotinin and elastatinal (Figure 5B), and strong digestion was observed in a similar pattern around the carcinoma cell nests mainly within the lumina of the tubular structures. In contrast, when serial sections were incubated on CCm-Tf films pretreated with a mixture of 1,10-phenanthroline, line, aprotinin, and elastatinal, the lysis around the carcinoma cell nests was almost completely inhibited (Figure 5C). The digestion around the carcinoma cell nests was also inhibited on CCm-Tf films impregnated with 1,10-phenanthroline, although the lysis observed on polymorphonuclear leukocytes infiltrated in the carcinoma tissues was not completely blocked on CCm-Tf films with 1,10-phenanthroline or a mixture of 1,10-phenanthroline, aprotinin, and elastatinal (data not shown). Immunohistochemically, MMP-7 was localized to such carcinoma cell nests (Figures 5D-5F). These results, together with high specificity of CCm-Tf to MMP-7, strongly suggest that the detected activity in the endometrial carcinoma cells is due to MMP-7.
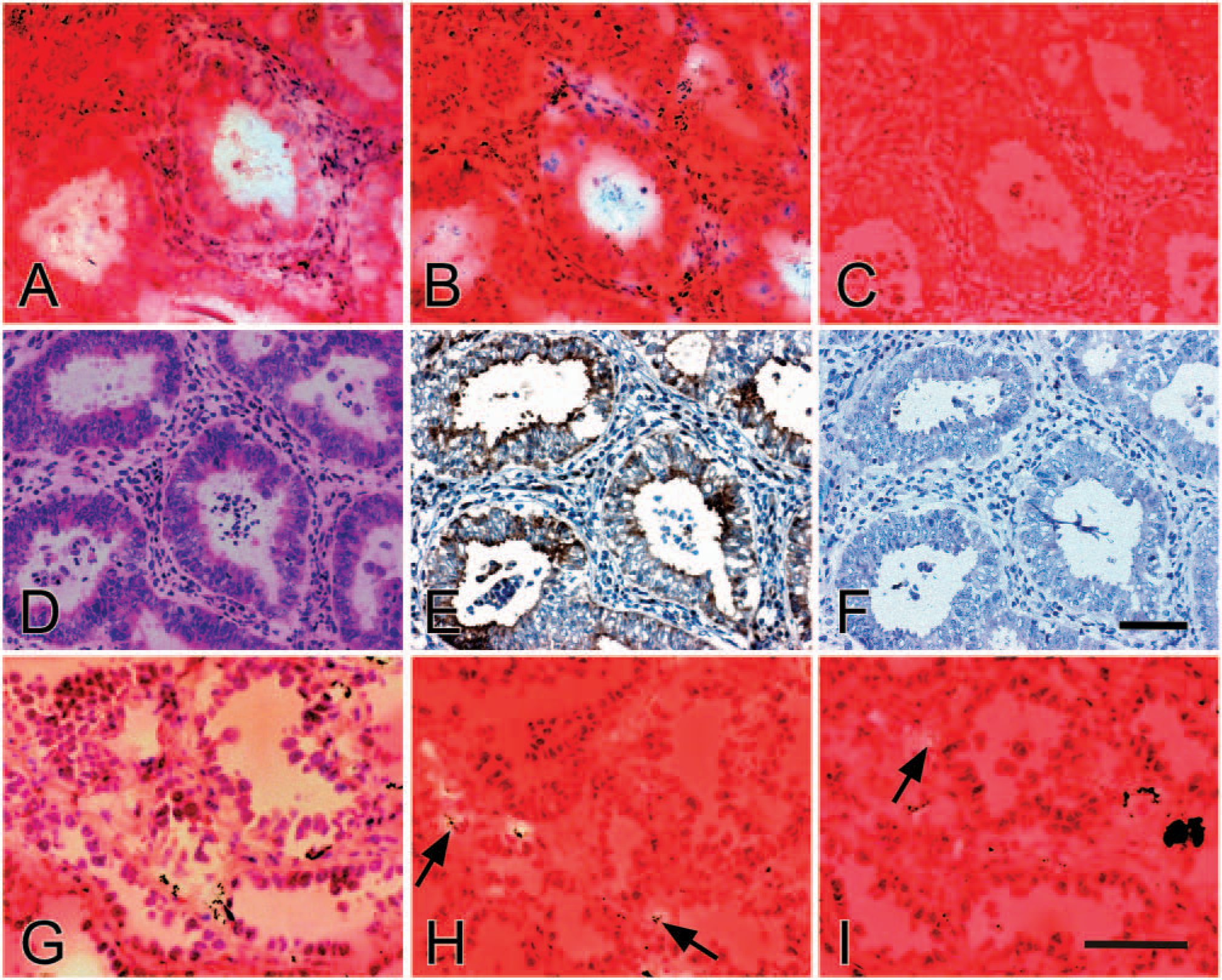
Localization of matrix metalloproteinase (MMP)-7 activity in the endometrial and lung carcinoma tissues by in situ zymography using crosslinked carboxymethylated transferrin (CCm-Tf) films and immunohistochemistry. Serial frozen sections of the endometrial carcinoma tissue were subjected to in situ zymography with CCm-Tf films
Similarly, frozen sections of human lung adenocarcinoma tissues, which are known to express MMP-7 (Kawano et al. 1997), were subjected to in situ zymography using CCm-Tf films. As shown in Figure 5G, marked digestion was obtained mainly around the carcinoma cell nests. When serial sections were incubated on CCm-Tf films impregnated with 1,10-phenanthroline, digestion around the carcinoma cell nests, but not on polymorphonuclear leukocytes infiltrated in the carcinoma tissue, was inhibited (Figure 5H). In addition, the lysis also completely disappeared on CCm-Tf films impregnated with a mixture of 1,10-phenanthroline, aprotinin, and elastatinal, although weak digestion was occasionally observed on polymorphonuclear leukocytes (Figure 5I). MMP-7 was immunolocalized to adenocarcinoma cells as described previously (Kawano et al. 1997) (data not shown). Altogether, these results show that in situ zymography using CCm-Tf films is useful to detect the activity of MMP-7 in the human endometrial and lung carcinoma tissues when combined with CCm-Tf films impregnated with proteinase inhibitors.
Discussion
In the present study, we have developed a new in situ zymography method to detect MMP-7 activity within the tissues. Because the previous biochemical study on the substrate specificity of MMP-7 (Imai et al. 1995) demonstrated that Cm-Tf is a preferable substrate of MMP-7 to MMP-3, we used Cm-Tf for coating the films. Our preliminary study showed that Cm-Tf on the films is readily degraded by MMP-7, but also lysed by MMP-3, although it is completely resistant to MMP-2. Thus we introduced crosslinking of Cm-Tf substrate and succeeded to enhance the selectivity for MMP-7. Because the molecular weight of MMP-3 (45 kDa) is 2.4-fold larger than that of MMP-7 (19 kDa), penetration of active MMP-3 into the crosslinked substrate seems to be more difficult than MMP-7. Actually, the sensitivity to MMP-3 activity was effectively diminished to an almost negligible level on the properly crosslinked Cm-Tf films. In addition, no or negligible digestion was obtained by incubation with MMP-1, -2, -9, or -13; MT1-MMP; MT3-MMP; or ADAMTS4 under the similar conditions. Furthermore, cysteine proteinases (cathepsin B, H, and L) and aspartic proteinase (cathepsin D) showed no activity on the CCm-Tf films. Although trypsin exhibited definite activity to non-crosslinked or weakly crosslinked Cm-Tf, the susceptibility was remarkably decreased on the adequately crosslinked Cm-Tf substrate, probably because of the modification of lysine residues at the trypsin cleavage sites with the crosslinking reagent. Thus the data suggest that the CCm-Tf films are rather selective to MMP-7 among these proteinases. Nevertheless, the CCm-Tf films were weakly susceptible to some serine proteinases including trypsin, chymotrypsin, elastase, plasmin, and cathepsin G. Thus inhibition of these proteinases by serine proteinase inhibitors was necessary to determine the activity of MMP-7 in tissue sections. Our data indicate that the activities of trypsin, chymotrypsin, and plasmin can be inhibited by pretreating the films with aprotinin, and that the treatment with elastatinal is effective for the inhibition of elastase. On the other hand, the activity of cathepsin G was not effectively inhibited with aprotinin and/or elastatinal. However, because the expression of cathepsin G is restricted to neutrophils (Salvesen 1998), the activity can be readily identified and differentiated from MMP-7 activity by in situ zymography of carcinoma tissues.
In contrast to CCm-Tf films, FIZ-GN using the crosslinked gelatin films was susceptible to the proteolysis by all the MMPs tested (MMP-1, -2, -3, -7, -9, and -13; MT1-MMP; and MT3-MMP), ADAMTS4, serine proteinases (trypsin, chymotrypsin, plasmin, elastase, and cathepsin G) and cysteine proteinases (cathepsin B, H, and L), but not by an aspartic proteinase (cathepsin D). These data indicate that gelatin is a nonspecific substrate of many proteinases, and crosslinking is not so effective in improving the selectivity. Thus FIZ-GN is useful for the detection of a variety of proteolytic activities in the tissues, but not suitable for the detection of specific MMP unless single MMP species is dominantly expressed in the tissue.
In situ zymography with quenched fluorogenic DQ-gelatin has been developed for detection of gelatinolytic activities of MMP-2 and MMP-9 (Mook et al. 2003). Because in this technique fluorescent peptides are produced by cleavage of DQ-gelatin at sites of gelatinolytic activity, the method appears to provide better tissue localization of the activity with higher sensitivity and quantification than FIZ-GN, in which staining intensity decreases at gelatinolytic areas (Frederiks and Mook 2004). However, FIZ-GN and in situ zymography using CCm-Tf films have an advantage in that one can preserve the films after the incubation and take microscopic pictures of the digestion by an ordinary light microscope, whereas the fluorescent signals by in situ zymography using DQ-gelatin quickly disappear during observation by a confocal microscopy. McIntyre et al. (2004) have recently developed a polymer-based fluorogenic substrate serving as a selective proteolytic beacon for MMP-7 activity and used the method for in vivo imaging by intravenous injection of the substrate. Although they mentioned in their article that the substrate could be used for in situ zymography for detection of MMP-7 activity in frozen sections of tumors (McIntyre et al 2004), the method is not yet established. Thus, to our knowledge, our in situ zymography using CCm-Tf films is the only method to localize MMP-7 activity within tissues at the present time.
We have applied the newly developed in situ zymography for MMP-7 to localize its activity in the human endometrial and lung adenocarcinoma specimens. By using CCm-Tf films impregnated with inhibitors to serine proteinases (aprotinin and elastatinal) and metalloproteinases (1,10-phenanthroline), we could demonstrate the metalloproteinase activity in the carcinoma cell nests. Because the activity was colocalized with MMP-7 shown by immunohistochemistry, it is conceivable that the activity is derived from MMP-7 produced by the carcinoma cells. In general, living tissues contain many proteinases, some of which may degrade CCm-Tf. Therefore, combining the present method with other supplementary techniques (i.e., in situ zymography using inhibitor-containing CCm-Tf films and MMP-7 immunohistochemistry) is considered to be important to determine the MMP-7 activity within the tissues.
In summary, we have developed a new technique to detect MMP-7 activity that is applicable to localize the activity within the tissues. MMP-7 is upregulated during tumorigenesis (Takeuchi et al. 1997; Wilson et al. 1997) and progression of carcinomas such as endometrial and gastrointestinal carcinomas (Yoshimoto et al. 1993; Mori et al. 1995; Ueno et al. 1999). Thus we believe that the present technique is useful to study the role of MMP-7 in pathophysiological conditions such as tumorigenesis and cancer progression and to evaluate the effects of synthetic inhibitors to MMP-7 activity in vivo.
Footnotes
Acknowledgements
We are grateful to Dr. Tsutomu Miyasaka for helpful discussions. We also thank Keiko Sukagawa and Hisae Kondo for excellent technical assistance.