
Abstract
In situ zymography is a very important technique that shows the proteolytic activity in sections and allows researchers to observe the specific sites of proteolysis in tissues or cells. It is normally performed in non-fixed frozen sections and is not routinely performed in calcified tissues. In this study, we describe a technique that maintains proteolytic activity in fixed and decalcified sections obtained after routine paraffin sectioning in conventional microtome and cryostat sections. We used adult rat hemimandibles, which presented bone, enamel, and dentine matrices; the substrate used was dye-quenched-gelatin. Gelatinolytic activity was colocalized with MMP-2 using fluorescent antibodies. Specific proteolytic activity was observed in all sections, compatible with metalloproteinase activity, particularly in dentine and bone. Furthermore, matrix metalloproteinase-2 was colocalized to the sites of green fluorescence in dentine. In conclusion, the technique presented here will allow in situ zymography reactions in fixed, decalcified, and paraffin-embedded tissues, and we showed that paraformaldehyde-lysine-periodate–fixed cryostat sections are suitable for colocalization of gelatinolytic activity and protein labeling with antibodies.
Keywords
Protease activity is essential for many physiological and pathological processes, such as embryonic development, organ and tissue morphogenesis, tissue remodeling, wound healing, inflammation, and tumor progression (Crawford and Matrisian 1994–1995; Ray and Stetler-Stevenson 1994; Abiko et al. 1999; Koyama et al. 2000; Waas et al. 2002; Yan and Blomme 2003). The ability to see the activity of proteases in tissues is very important to determine their role.
There are many techniques used to observe and quantify proteases. IHC localizes the presence of specific proteases in tissues, but zymogens cannot be normally differentiated from active proteases by this technique (Frederiks and Mook 2004). ELISAs are also widely used to quantify proteases, especially in fluids, with the disadvantages of not discriminating between active and inactive proteases and no possibility of determining the localization of the enzyme in the tissue. Zymography and Western blotting are assays used to analyze enzyme activity and the amount of protein, respectively, but do not allow the localization of proteases in tissues (Frederiks and Mook 2004). In situ zymography was developed to observe the presence of activated protease in tissues (Galis et al. 1994), with the possibility to colocalize proteolytic activity with specific proteases using antibodies directed to those proteases.
Initially, in situ zymography was performed using an autoradiographic emulsion containing gelatin (Galis et al. 1994, 1995). The gelatinolytic activity resulted in decreased amounts of silver in specific sites that generated dark spots after radiographic development. Fluorescein-coupled gelatin was introduced as an advance in the localization of gelatinolytic activity in tissues (Galis et al. 1995). However, the precise localization of the proteolytic activity in tissues was only improved with the introduction of dye-quenched (DQ)-gelatin, which is a gelatin substrate heavily labeled with FITC molecules, so that fluorescence is quenched unless proteolysis occurs (Oh et al. 1999; Goodall et al. 2001; Lindsey et al. 2001; Pirilä et al. 2001; Teesalu et al. 2001; Wang and Lakatta 2002; Zhang and Salamonsen 2002; Mook et al. 2003; Lee et al. 2004; Sakuraba et al. 2006). Using DQ-substrates, proteolytic activity is observed as fluorescence in a conventional fluorescence microscope. Other quenched fluorogenic substrates developed and commercially available are DQ-casein, DQ-collagen type I, DQ-collagen type IV, and DQ-elastin (Frederiks and Mook 2004).
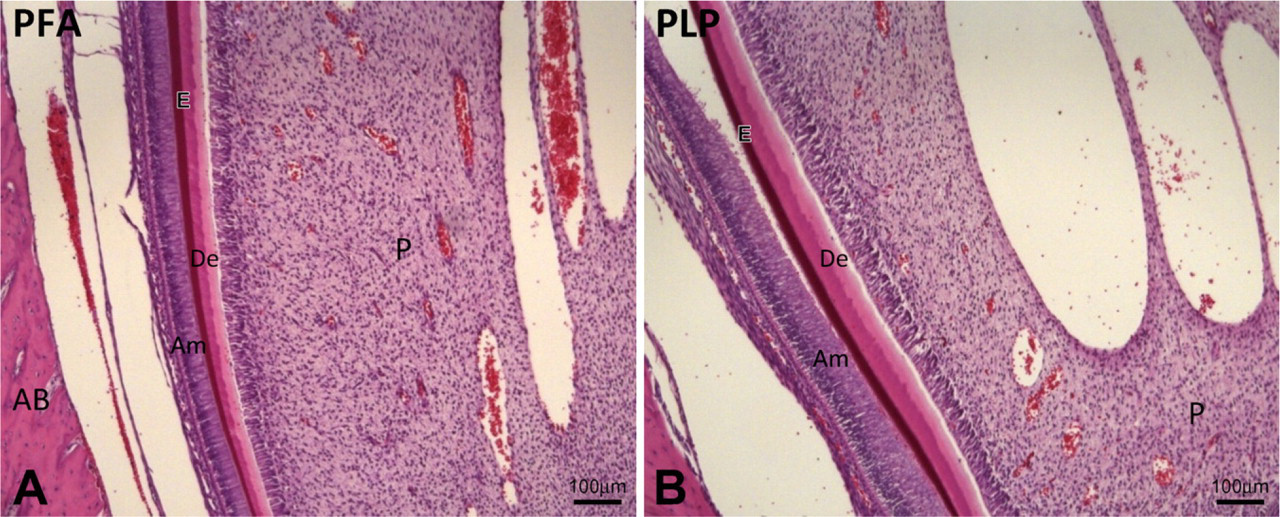
Longitudinal sections of the odontogenic zone from a rat lower incisor showing secretory stage ameloblasts.
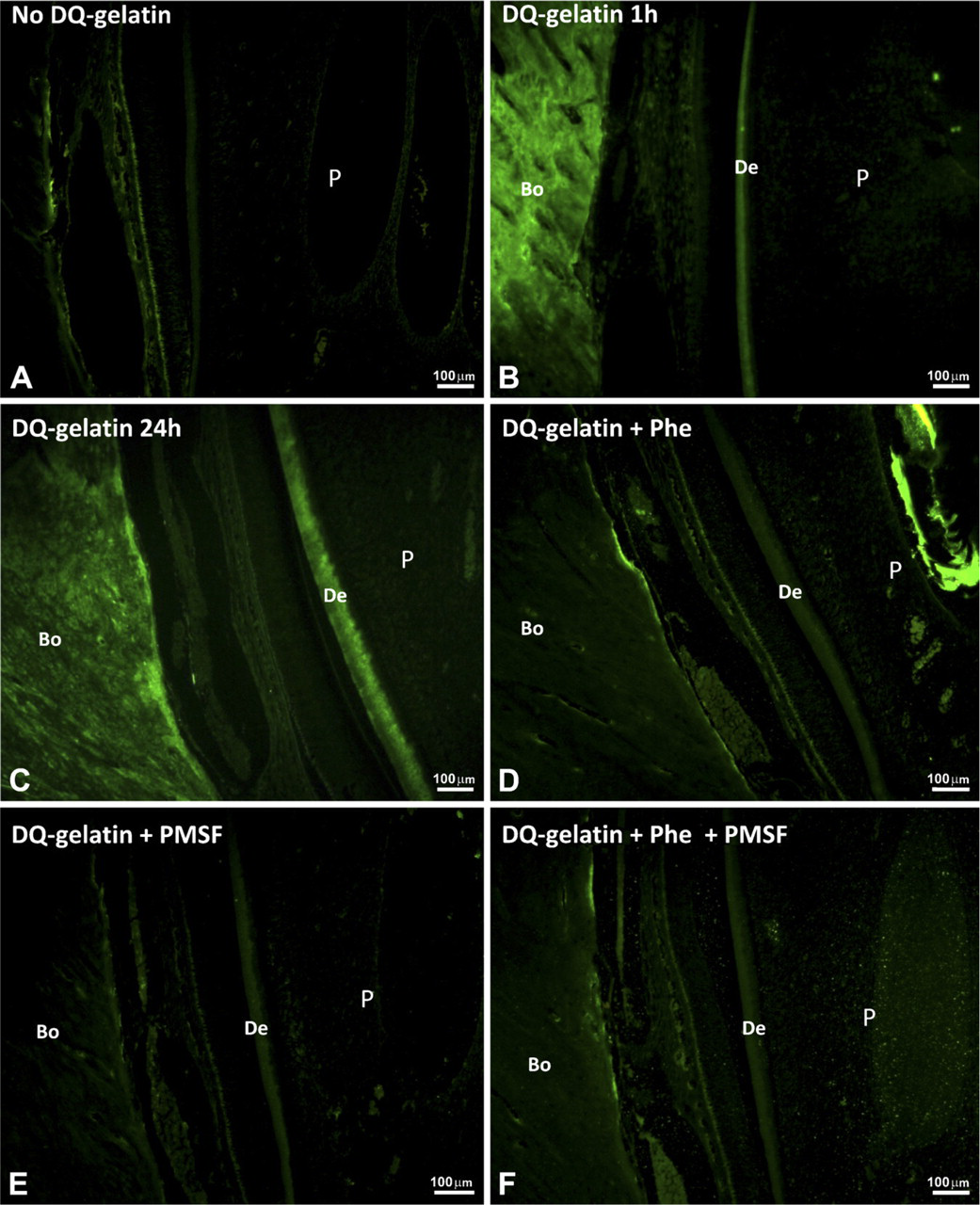
Gelatinolytic activity in forming rat incisor is shown as green fluorescence in sections fixed with PLP.
Proteases are widely distributed in craniofacial tissues (Abiko et al. 1999) such as oral mucosa (Tsai et al. 2003; Patel et al. 2005; Rajendran et al. 2006), gingiva (Ejeil et al. 2003; Smith et al. 2004; Kumar et al. 2006; Naveau et al. 2007), salivary glands (Kato et al. 1995; Broverman et al. 1998; Nagel et al. 2004), tooth buds (Abiko et al. 1999), dentine (van Strijp et al. 2003; Mazzoni et al. 2007), and forming enamel (Gerlach et al. 2000a, b; Goldberg et al. 2003; Bourd-Boittin et al. 2004). The presence of proteases in craniofacial tissues has been characterized by most authors by IHC (Saruta et al. 2005; Fukumoto et al. 2006; Takamori et al. 2008). Zymography was also used in a variety of studies (Iamaroon et al. 1996; Abiko et al. 1999; Gerlach et al. 2000a, b; Goldberg et al. 2003; Bourd-Boittin et al. 2004; Sakuraba et al. 2006). However, to date, only two studies showed the proteolytic activity in calcified craniofacial tissues in situ (Sakuraba et al. 2006; Sakakura et al. 2007), and many important aspects of proteolysis are not known yet in those tissues.
The major challenge to perform in situ zymography in craniofacial tissues is to prepare thin sections of mineralized tissues without losing proteolytic activity, because those sections are routinely prepared using formaldehyde-based fixatives, followed by decalcification and inclusion in paraffin. Most studies using in situ zymography were done in soft tissues, in which proteolytic activity was observed in non-fixed tissues from which sections were directly cut in a cryostat. In this study, we describe the feasibility of performing in situ zymography in adult craniofacial tissues using the technique described by Rocha et al. (2006) that enables the localization of alkaline phosphatase activity in bone based on the original paraformaldehyde-lysine-periodate (PLP) fixation (McLean and Nakane 1974) or in the 4% paraformaldehyde fixation and EDTA-glycerol decalcification procedure (Mori et al. 1988). The techniques described in this study maintain proteolytic activity in sections prepared and handled routinely in histology laboratories. This assay was developed using adult rat hemimandibles sections, and the proteolytic activity was shown using DQ-gelatin. Gelatinolytic activity was colocalized with MMP-2 using fluorescent antibodies.
Materials and Methods
This study was approved by the Animal Ethics Committee (protocol number 01.1364.53.4). Six 3-month-old Wistar rats were used in this protocol. The animals received water and food ad libitum. After the animals were killed, hemimandibles were removed.
Tissue Preparation
The hemimandibles were immersed in PLP fixative (2% paraformaldehyde, 0.075 M lysine, and 0.01 M sodium periodate, pH 7.4) or in 4% paraformaldehyde (PFA), pH 7.4, at 4C for 48 hr. Thereafter, hemimandibles were washed with PBS containing increasing volumes of glycerol (5%, 10%, and 15%) and decalcified with EDTA-glycerol, pH 7.3 (1.25 g EDTA, 1.25 g NaOH, and 15 ml glycerol in 100 ml of distilled water) at 4C for 1 month. The material was washed in the following solutions for 12 hr each: (a) 15% sucrose and 15% glycerol, (b) 20% sucrose and 10% glycerol, (c) 20% sucrose and 5% glycerol, (d) 20% sucrose, (e) 10% sucrose, (f) 5% sucrose, and (g) 100% PBS.
The odontogenic area of each hemimandible was selected, cut longitudinally, and included in a paraffin of lower fusion point (54C) or frozen in O.C.T. compound by liquid nitrogen. Five-μm longitudinal sections were cut in a microtome (model RM2165; Leica, Nussloch, Germany) or in a cryostat (model CM1510; Leica) and laid on silanized microscope slides. Some sections were stained with hematoxylin and eosin (HE). In situ zymography reactions were done in deparaffined non-stained sections within a month of sectioning.
In Situ Zymography Using DQ-gelatin
The mixture of DQ-gelatin (DQ-gelatin, E12055; Molecular Probes, Eugene, OR) was prepared as follows: 1 mg/ml DQ-gelatin dissolved 1:10 in 50 mM TrisCaCl2. Forty μl of this mixture was applied on top of the sections, which were incubated at 25C for 1 hr or 37C for 24 hr in a dark humid chamber. The gelatinolytic activity was observed as green fluorescence (absorption maxima, ∼495 nm; fluorescence emission maxima, ∼515 nm) by fluorescence microscopy (Fluorescence Microscope Model Axioskop 50 using camera AxioCam MRC; Carl Zeiss, Oberkochen, Germany).
Negative control sections were incubated in the same way as described above, but without DQ-gelatin. Some sections were incubated either with an metalloproteinase inhibitor, 1,10-phenanthroline (Phe), with a serine protease inhibitor, PMSF (Sigma Chemicals; St. Louis, MO), at 2 mM, or with both inhibitors.
Matrix Metalloproteinase-2 (MMP-2) Immunolabeling
Sections of tissue fixed with PLP and cut in a cryostat were incubated with a mixture of DQ-gelatin. All incubation steps were carried out in a dark humid chamber at room temperature. After 1 hr, the sections were rinsed three times in 0.01 M PBS and then were incubated for 1 hr with anti-MMP-2 monoclonal antibody (mouse anti-MMP-2 antibody MAB3308; Chemicon, Temecula, CA) diluted 1:100 in PBS. After rinsing with PBS, the sections were incubated with rhodamine (Rhodamine Conjugated Anti-Mouse Antibody; Chemicon) diluted 1:200 in PBS for 1 hr and then rinsed three times with PBS. 4′,6-Diamidino-2-phenylindole (DAPI) was applied for 3 min, and specimens were washed with PBS and analyzed by fluorescence microscopy.
Results
The sections fixed with PLP or PFA and stained with HE showed a normal odontogenic zone. Enamel matrix and dentine were observed, and secretory stage ameloblasts were present (Figure 1).
After a 1-hr incubation at 25C, sections fixed with PLP and treated with DQ-gelatin exhibited some fluorescence in the dentine and bone, indicating gelatinolytic activity (Figure 2B) that increased after a 24-hr incubation (Figure 2C). The sections incubated without DQ-gelatin showed only a faint signal, mainly caused by autofluorescence of cells, and the gelatinolytic activity in dentin and bone was not observed (Figure 2A). Sections treated with DQ-gelatin + Phe, DQ-gelatin + PMSF, or both inhibitors showed decreased gelatinolytic activity in dentine and alveolar bone after 1 hr (Figures 2D–2F); the same happened after 24 hr (data not shown).
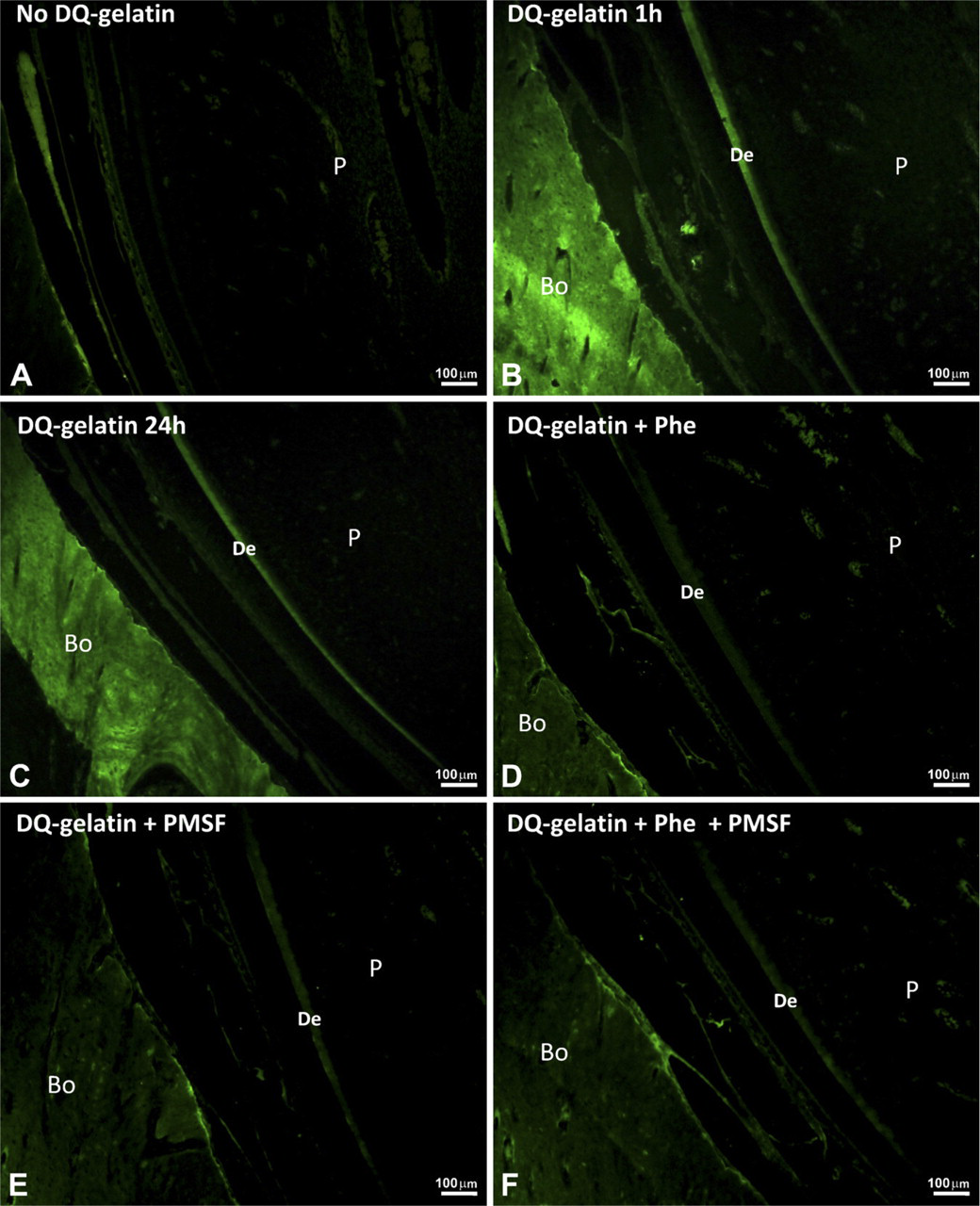
Gelatinolytic activity in forming rat incisor is shown as green fluorescence in sections fixed with PFA.
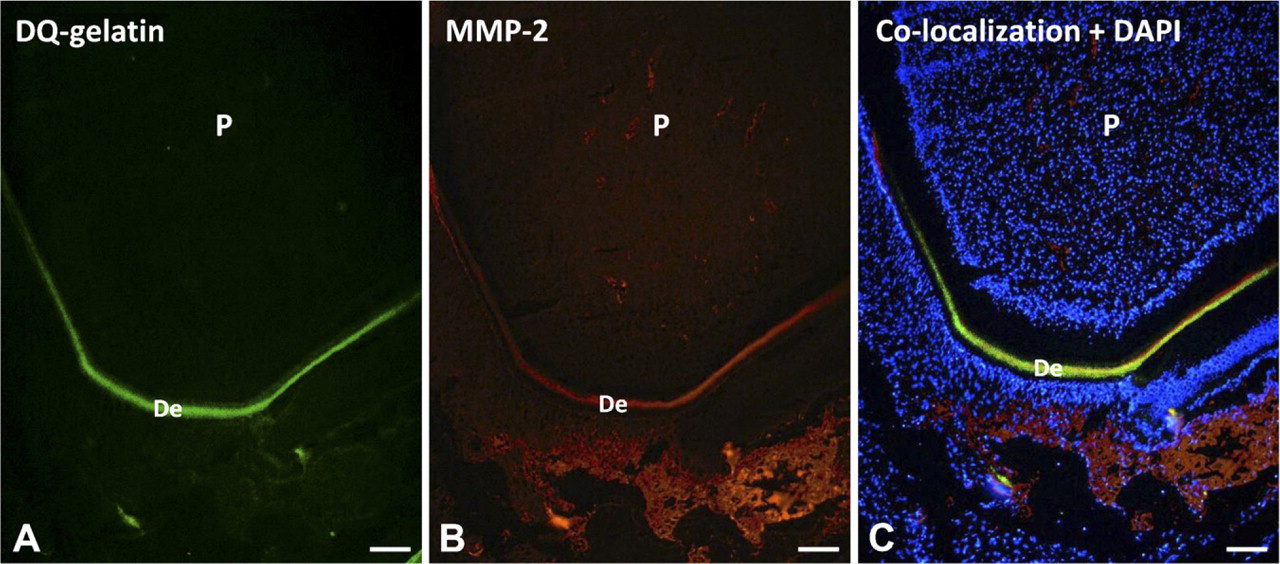
Gelatinolytic activity and metalloproteinase-2 (MMP-2) colocalization in forming rat incisor are shown in sections fixed with PLP.
A similar pattern was observed in the sections fixed with PFA. In sections incubated without DQ-gelatin, the autofluorescence of cells was observed but no gelatinolytic activity was present (Figure 3A). In sections incubated with DQ-gelatin, the gelatinolytic activity was observed in dentine and alveolar bone after a 1-hr incubation (Figure 3B). The gelatinolytic activity was still observed after a 24-hr incubation, but no increased activity was seen after this time (Figure 3C). The sections treated with inhibitors showed a decrease in gelatinolytic activity (Figures 3D–3F). No gelatinolytic activity was observed in dental enamel.
Specimens fixed with PLP and PFA were also frozen in O.C.T., and cryostat sections were cut and incubated with DQ-gelatin. Results showed the same pattern observed in Figures 2 and 3, with gelatinolytic activity concentrated in dentine and bone and faint fluorescence in sections with no substrate (data not shown). Figure 4 shows a PLP-fixed, cryostat-cut section incubated with DQ-gelatin (Figure 4A) and a monoclonal anti-MMP-2 antibody followed by a secondary antibody labeled with rhodamine (Figure 4B). DAPI was used as a nuclear stain (Figure 4C). Green fluorescence compatible with gelatinolytic activity is observed in dentine and was colocalized with MMP-2, a gelatinolytic proteinase (Figure 4C).
Discussion
In situ zymography using dye-quenched substrates was developed to improve the localization of protease activity in frozen sections of non-fixed tissues. In our study, we showed for the first time the feasibility of doing in situ zymography in fixed and decalcified sections of craniofacial tissues from adult animals. This technique was previously used to show the activity of alkaline phosphatase in bone (Rocha et al. 2006) and is based on the use of PLP fixative, as described by McLean and Nakane (1974), and EDTA-glycerol decalcification (Mori et al. 1988). Decalcification and inclusion of the tissue in low fusion point paraffin allowed sectioning in a conventional microtome, and the results suggest that gelatinolytic activity was preserved in dentine and bone of PLP- and PFA-fixed specimens. Incubation for 1 hr at room temperature was adequate to obtain a strong fluorescent signal.
The green fluorescence observed is compatible with gelatinolytic activity, because it decreased when inhibitors were used (although inhibition was not specific by Phe or PMSF). Furthermore, MMP-2, which is a gelatinolytic proteinase, was colocalized to the sites of green fluorescence in dentine. Some green fluorescent signal (compatible with DQ-gelatin proteolysis) was observed in other parts of the tissue, where no MMP-2 was detected using antibodies. The gelatinases (MMP-2 and MMP-9) have domains that bind gelatin (the fibronectin-like domain) (Briknarová et al. 2001; Xu et al. 2004; Hornebeck et al. 2005), and these domains might bind the substrate where these proteinases are localized in the sections, but this is probably not the reason that explains the strong green fluorescence found in dentine and bone.
The fluorescent signal compatible with gelatinolytic activity and the colocalization with MMP-2 is also in agreement with studies that indicate that MMP-2 is found in dentine (Satoyoshi et al. 2001; Goldberg et al. 2003; Palosaari et al. 2003; van Strijp et al. 2003) and bone (Maruya et al. 2003; César Neto et al. 2004; Accorsi-Mendoça et al. 2008). MMP-2 is probably one of the proteinases whose activity might be preserved to some extend even using paraformaldehyde-based fixatives, because this proteinase has been shown to be resistant to harsh conditions; for example, extracted teeth still showed MMP-2 activity after complete acid demineralization of the dentine in the absence of a fixative (Mazzoni et al. 2007).
Although many studies of in situ zymography have been described in soft tissues (Abiko et al. 1999; Oh et al. 1999; Duchossoy et al. 2001; Goodall et al. 2001; Lindsey et al. 2001; Pirilä et al. 2001; Teesalu et al. 2001; Wang and Lakatta 2002; Zhang and Salamonsen 2002; Mook et al. 2003; Lee et al. 2004; Smith et al. 2004), only a few have described the activity of proteinases in calcified tissues. To our knowledge, Sakuraba et al. (2006) were the first to describe in situ zymography in tooth germs of young animals, using quenched fluorogenic substrates in non-fixed frozen sections prepared in a cryostat. However, the technique cannot be universally used, particularly in calcified tissues of adult animals, because those cannot be normally processed in a cryostat. In our view, the technique presented here has an important advantage, because more laboratories familiar with conventional histological processing will probably be able to look for enzyme activity in bones and teeth.
Another important aspect is that in situ zymography allows colocalization of the proteolytic activity with proteinases using specific antibodies. With the recent increase in the number of quenched fluorogenic substrates available and increasing number of fluorophores available to label secondary antibodies, probably many proteases can be localized to the site of proteolytic activity, and this activity can certainly be monitored over time and checked using specific inhibitors, which expands the arsenal of tools a researcher has to look for the role of one or many proteinases in calcified tissues.
Colocalization of proteolysis with antibody labeling is a reality in soft non-fixed tissues (Evans and Itoh 2007). We showed here that PLP-fixed cryostat sections are suitable for colocalization of gelatinolytic activity and protein labeling with antibodies recommended for IHC.
The fixation of calcified tissues described here may also be useful to prepare samples for embedding in resin and sectioning in hard tissues saws, without losing proteolytic activity, but detailed protocols need to be tested. This is the procedure to avoid altering the mineral content of mineralized tissues. Particularly, new aspects of mineralization of enamel in the maturation stage and the role of proteinases in mineralization will probably be possible in this way.
In conclusion, the technique described here will allow doing in situ zymography in fixed, decalcified, and paraffin-embedded tissues. The use of this technique coupled with the use of specific inhibitors and antibodies will contribute to the understanding of the specific roles played by proteinases in calcified tissues.
Footnotes
Acknowledgements
We gratefully acknowledge the support of Fundação de Pesquisa do Estado de Säo Paulo and Cobnselho Nacional de Pesquisa.
We thank Maria Elena Riul and Mônica Azevedo de Abreu, Department of Pathology, Medicine School of Ribeiräo Preto, for excellent technical assistance in preparation of the specimens.