
Abstract
Leptin is a hormone that plays important roles in nutritional status and in obesity. By means of immunocytochemistry, two populations of leptin-secreting cells were found in the lower half of the gastric mucosa. One consists of numerous large cells located around the gastric pits, the Chief epithelial cells, whereas the second refers to much smaller cells, strongly stained, few in number, and scattered between the gastric pits, the endocrine cells. By double immunostaining, leptin and pepsinogen were colocalized in the Chief cells, whereas the endocrine cells were positive only for leptin. Immunoelectron microscopy showed that leptin is present along the rough endoplasmic reticulum–Golgi-granules secretory pathways of the Chief and endocrine cells. On the other hand, leptin-receptor (long and short forms) immunolabelings, although absent in the gastric epithelial cell plasma membranes, were present in enterocytes at the level of their apical and basolateral membranes. Duodenal, jejunal, and ileal enterocytes displayed similar labelings for the leptin receptor. Thus, exocrine and endocrine secretions of leptin together with the presence of leptin receptors on enterocyte plasma membranes constitute a gastroenteric axis that coordinates the role played by leptin in the digestive tract.
I
In addition to adipocytes, several other organs such as the stomach, placenta, and brain are able to synthesize leptin (Masuzaki et al. 1997; Bado et al 1998; Ahima et al. 2000; Havel 2000; Wilkinson et al. 2000). In particular, the gastric mucosa appears so far to be the only tissue secreting leptin in an exocrine rather than endocrine fashion. Immunohistochemistry revealed that Chief cells (Buyse et al. 2004) of the fundic mucosa contain leptin in their secretory granules. Upon stimulation by nervous or hormonal factors, leptin is released in the gastric juice linked to a protein of high molecular weight, and this complex is targeted in its intact form to the duodenum (Guilmeau et al. 2003). Indeed, exocrine leptin has been shown to exert pleiotropic effects on the stomach and intestine. On the stomach, it controls meal size in cooperation with cholecystokinin, stomach emptying, and cytoprotection of the gastric mucosa (Brzozowski et al. 1999; Attoub et al. 2000; Barbier et al. 2000). In rats, duodenal infusion of leptin increases plasma levels of cholecystokinin, a satiety factor, which in turn stimulates leptin secretion by the gastric mucosa. This constitutes a positive feedback loop (Guilmeau et al. 2003). In the small intestine, leptin is involved in the inflammatory process as suggested by ob/ob mice resistant to intestinal inflammation (Siegmund et al. 2002). Moreover, leptin regulates secretion of gastric hormones such as gastrin and somatostatin and controls the absorption of nutrients, increasing peptides and decreasing fat and carbohydrate absorption (Morton et al. 1998; Buyse et al. 2001). Leptin seems therefore deeply involved in the functional activities of the digestive tract.
Like other hormones, leptin exerts its effect by binding to its receptor. Leptin receptor belongs to the type I cytokine receptor family and presents an amino acid sequence homology with the 130-kDa glycoprotein, the gp130 (Tartaglia et al. 1995; Lee et al. 1996). Several isoforms of leptin receptor exist: one long form (OB-Rb), four shorter forms (OB-Ra, c, d, f), and a soluble one (OB-Re) (Ahima and Osei 2004). Leptin affinity to all receptors is similar (Liu et al. 1997). Membrane receptors have the ability to activate intracellular mediators including JAK/STAT, IRS, PI3K, and AMP kinase (Bjorbaek and Kahn 2004). OB-Rb is the longest form and is able to activate all pathways, whereas other isoforms with shorter intra-cellular domains have a more limited role in signal transduction (Bjorbaek et al. 1997).
Various leptin receptor isoform mRNAs were identified in the gastric mucosa as well as in duodenum, jejunum, and ileum (Tartaglia et al. 1995; Sobhani et al. 2000; Buyse et al. 2001; Goiot et al. 2001). However, despite extensive studies on the role of leptin in the gastrointestinal tract, the endocrine secretion of leptin by the gastric mucosa and the precise location of leptin receptors in the digestive tract are still a matter of debate. In particular, it remains to be defined if leptin receptors are present on the luminal or basolateral membranes of enterocytes, which is of primary importance to understand the mechanisms of action of leptin. On the other hand, it was suggested that gastric leptin secreted in an endocrine fashion must act on liver (Guilmeau et al. 2004). Indeed, food intake increases plasma leptin levels very rapidly (15 min) in fasted rats. As white adipocytes from fasted rats secrete only small amounts of leptin (Gettys et al. 1996), it is possible that other tissues (such as the fundus) participate in rapidly increasing plasma leptin levels after food intake.
The aim of the present study is to review and to further identify and characterize exocrine and endocrine leptin-secreting cells in the gastric mucosa and to determine the precise localization of leptin receptors in the digestive tract. We now report that leptin is present in Chief cells as well as in typical endocrine cells of the gastric mucosa at the level of the fundus. Moreover, leptin receptors were revealed all along the small intestine, both at apical and basolateral membranes, but are absent in gastric epithelial cell membrane. This suggests that leptin plays a key role in a gastroenteric axis, triggered by the gastric mucosa to exert a paracrine action on the intestine: exocrinesecreted leptin should act on enterocyte apical membrane receptors whereas endocrine leptin from fundus and other sources must bind to enterocyte basolateral membrane receptors.
Materials and Methods
Biological Material
Stomach, duodenum, jejunum, and ileum were sampled from 100 to 115 g Sprague-Dawley male rats and processed for biochemical and morphological studies. Rats were fasted for 18 hr and anesthetized with urethane prior to surgery. Gastric juice was collected during 30 min upon a carbachol (Sigma-Aldrich; Oakville, Canada) injection (12 mg/kg body weight) from a series of rats.
Fasted rats had free access to water and were kept in individual cages. The animals were handled following the guidelines of the Canadian Council on Animal Care (CCAC).
Antibodies
Polyclonal antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, CA): rabbit antibody against leptin (Ob A20), goat antibody against pepsinogen (AB 9013–1), rabbit antibody against the extracellular portion of the leptin receptor (OBR H300), and goat antibody against the intracellular domain of leptin receptor long-form RB-Ob (OBR C20). Secondary antibodies to goat (rhodamine AP184R) and rabbit (FITC AP187F) IgGs were purchased from Chemicon (Temecula, CA).
Immunocytochemistry
Light Microscopy. Entire stomach and pieces of duodenum, jejunum, and ileum were fixed in Bouin's fixative and embedded in paraffin. Five-μm-thick sections were mounted on Superfrost slides (Fisher Scientific; Montréal, Canada) and processed for immunohistochemistry. Tissue sections were deparaffinized and washed twice in 10 mM phosphate buffer, pH 7.4, 150 mM NaCl (PBS). After a 1-min incubation with 1 mg/ml pepsin in 0.01 M HCl, samples were washed three times in PBS and covered with ovalbumin 1% for 30 min. Incubation with the chosen antibody (anti-leptin 10 μg/ml, anti-pepsinogen 1 μg/ml, or anti-leptin receptor 10 μg/ml) was carried out overnight at 4C. Sections were then washed twice in PBS, incubated with a secondary antibody for 1 hr, washed three times in PBS, and mounted with 1% DABCO in 50% glycerol.
For double immunolabeling, after overnight incubation with rabbit anti-leptin antibody the sections were washed and incubated with the goat anti-pepsinogen antibody for 2 hr, washed with PBS, and then incubated with an anti-goat IgG–rhodamine antibody and an anti-rabbit IgG–FITC antibody. Finally, sections were washed before mounting in DABCO/glycerol (1%/50%). The sections were examined with a LEITZ DMRB fluorescence microscope and a LEICA DM-IRBE inverted confocal laser microscope (Leica; St-Laurent, Canada).
Electron Microscopy. Tissues samples were fixed by immersion with 1% (v/v) glutaraldehyde in 0.1 M sodium phosphate buffer, pH 7.4, for 2 hr at 4C. Washing in phosphate buffer, dehydration in a series of graded methanol solutions at decreasing temperature until reaching −30C, and embedding in Lowicryl (K4M CANEMCO; St-Laurent, Canada) were carried out as described previously (Bendayan 1995). For embedding in Epon, the tissue fragments were postfixed with 1% osmium tetroxide for 1 hr at 4C, dehydrated in ethanol and propylene oxide, infiltrated, and embedded in Epon 812 according to standard techniques. Thin sections were mounted on Parlodion and carbon-coated nickel grids. Grids were incubated on a drop of saturated sodium periodate solution, then on glycine (0.15 mM in PBS, pH 7.4) at room temperature, followed by ovalbumin 1% in PBS. Incubation with the antibody was carried out overnight at 4C. Tissue sections were washed on PBS, incubated on 1% ovalbumin, and then on the protein A-gold complex (10 nm) for the rabbit antibody or on anti-goat IgG–gold complex (5 nm) for the goat antibody. Grids were then washed thoroughly with PBS and distilled water. Thin sections were stained with uranyl acetate and examined with a Philips 410LS electron microscope (FEI Systems Canada; St-Laurent, Canada).
Control Experiments
Control experiments for light and electron microscopy were performed. Primary antibody was omitted from the labeling protocol or sections were incubated with antigen-preadsorbed antibody complexes.
Evaluation
Morphometrical evaluation of the gold labelings was performed using a Clemex Vision 3 image analyzer system (Clemex; Longueuil, Canada). Micrographs were recorded at X1200 magnification and analyzed at X28,000 final magnification. Labeling densities were evaluated as described previously (Bendayan 1995) and expressed as number of gold particles per micrometer square ± standard error of the mean. Student's t-test was used to carry out statistical analysis.
Results
Characterization of Leptin-secreting Cells in the Gastric Mucosa
To precisely localize leptin in the gastric mucosa, paraffin sections of the stomach were incubated with an antibody against leptin and revealed by a FITC-secondary antibody. At low magnification, cells positive for leptin were easily detected in the lower half of the gastric mucosa (Figure 1). Positive cells for leptin were present immediately after the cardia portion. At higher magnification, it became clear that we had two populations of cells. One type was located along the gastric glands and showed cytoplasmic staining for leptin, whereas the other type was found between the gastric glands and were much smaller in size but strongly stained. By double immunolabeling, leptin (Figure 2A) and pepsinogen (Figure 2B) were found colocalized in the same epithelial cells along the gastric glands. No cells were found positive for pepsinogen alone. On the other hand, the small leptin-positive cells located between the glands appeared negative for pepsinogen. These small cells displayed the typical morphology of endocrine cells, few in number and scattered throughout the mucosa (Figure 2C). Nuclei of all cells appeared negative for leptin, and controls carried out in parallel confirmed the specificity of the stainings.

Immunohistochemical staining of leptin in the gastric mucosa (fundus) of a normal rat. The tissue section (5 μM) of fundic mucosa was incubated with an anti-leptin antibody followed by an FITC-conjugated secondary antibody. The cells from the middle to the lower part of the gland display a positive reaction. No staining is present in the gastric lumen (L) and the lamina propria (LP).
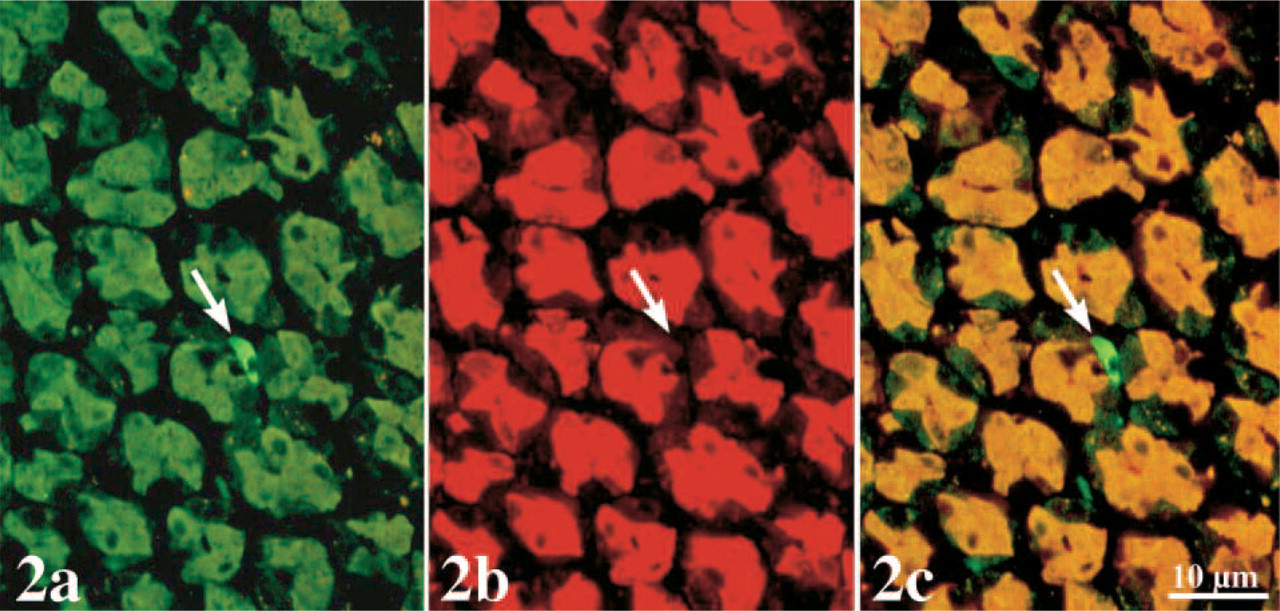
Double immunostaining of leptin
At the electron microscope level, incubation of the ultra-thin sections of the gastric mucosa with an anti-leptin antibody confirmed the results obtained by light microscopy. Chief cells were heavily stained with gold particles (Figure 3). Labeling was restricted to the rough endoplasmic reticulum (RER), the Golgi apparatus, and the numerous secretory granules (Figure 3). The secretory granules were located at the apical region of the cells close to the lumen of the gastric glands. Mitochondria and nuclei were devoid of any specific labeling by gold particles. Quantitative evaluations confirmed the subjective observation and demonstrated the existence of an increasing gradient in intensities of labeling along the RER–Golgi-granule secretory pathway (Table 1). They also ascertained the specificity of the labelings (Table 1). In addition, cells with the classical morphology of endocrine cells displayed numerous small secretory granules labeled by gold particles. RER and Golgi apparatus also displayed labeling by some gold particles. These cells were located within the connective tissue closely associated with blood capillaries (Figure 4). Double labeling for leptin and pepsinogen revealed a colocalization of both antigens in secretory granules, RER, and Golgi apparatus of the Chief cells in the gastric glands (Figure 5). No labeling for pepsinogen was detected in the small endocrine cells (Figure 2). These results indicate the presence of two types of leptin-secreting cells in the gastric mucosa, one of the exocrine type, the Chief cells, with granules oriented toward the gastric lumen and containing leptin and pepsinogen, and one of the endocrine type, located near blood capillaries containing only leptin.
Localization of Leptin Receptor in Stomach and Intestine
Because leptin is secreted by the gastric mucosa into the gastric juice and thus vehiculated to the intestine, we further investigated the localization of leptin receptors at the level of luminal and basolateral membranes of gastric and intestinal cells. Labeling was carried out with an antibody directed against the extracellular domain of the leptin receptor. This antibody recognizes all isoforms (long and short, namely, OB-Ra, b, c, d, and f) of the leptin receptors. No labeling was found associated with the plasma membranes of gastric cells (results not shown). This observation was confirmed by electron microscopy (results not shown). On the other hand, duodenum, jejunum, and ileum showed a positive signal for leptin receptors (Figure 6). The brush border of the enterocytes displayed a strong staining, while a weaker one was present at the base of the cells (Figure 6). Ultrastructural studies with immunogold confirmed the presence of leptin receptors over enterocyte microvilli and basolateral membranes particularly at the level of the interdigitations (Figure 7 and Figure 8). Quantitation of gold particles did not reveal statistical differences in labeling densities between the three portions of the intestine (results not shown). These results indicate that leptin produced by the fundic mucosa can act primarily on the intestinal wall rather than on the stomach itself. This action may involve both the endocrine and the exocrine secretions of leptin.
Quantitative evaluation of leptin immunolabeling in different cellular compartments of the Chief cells from the rat gastric mucosa
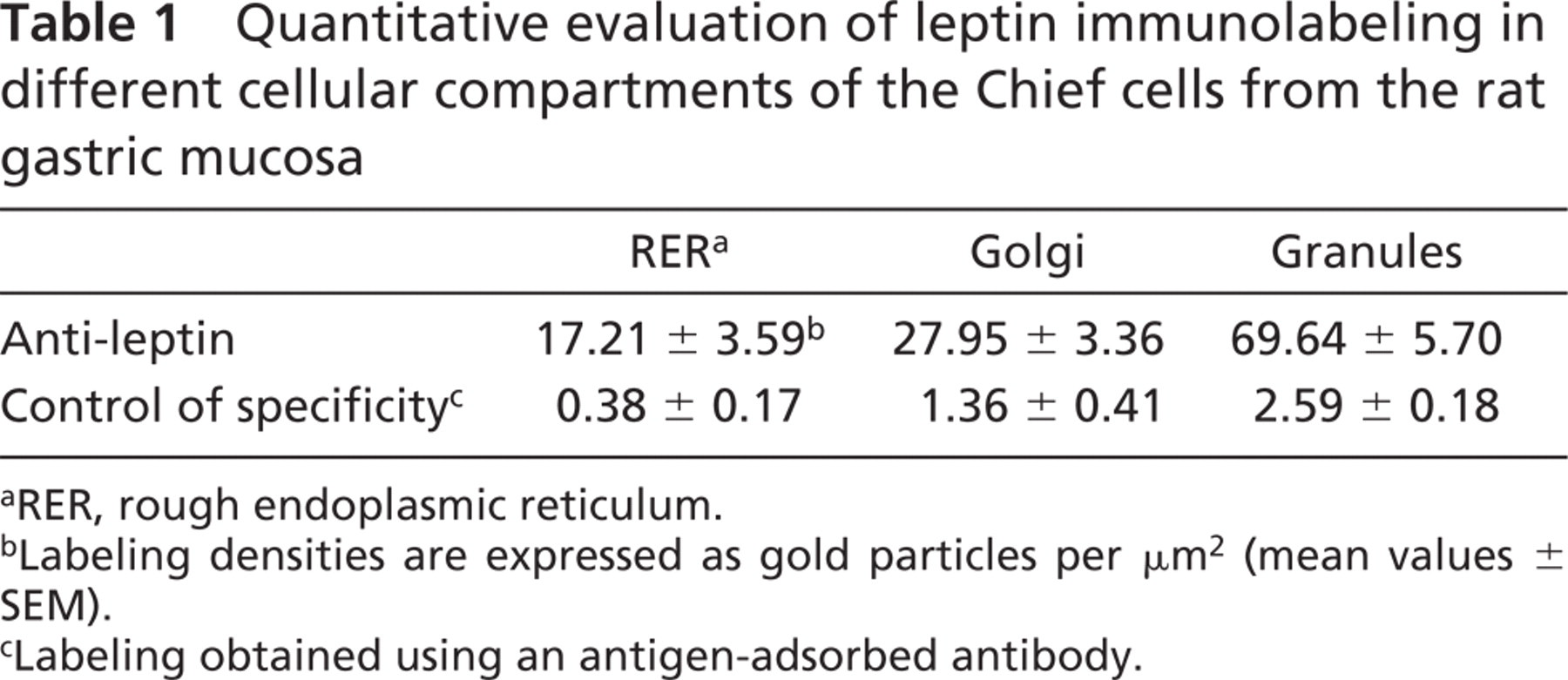
aRER, rough endoplasmic reticulum.
bLabeling densities are expressed as gold particles per μm2 (mean values ± SEM).
cLabeling obtained using an antigen-adsorbed antibody.
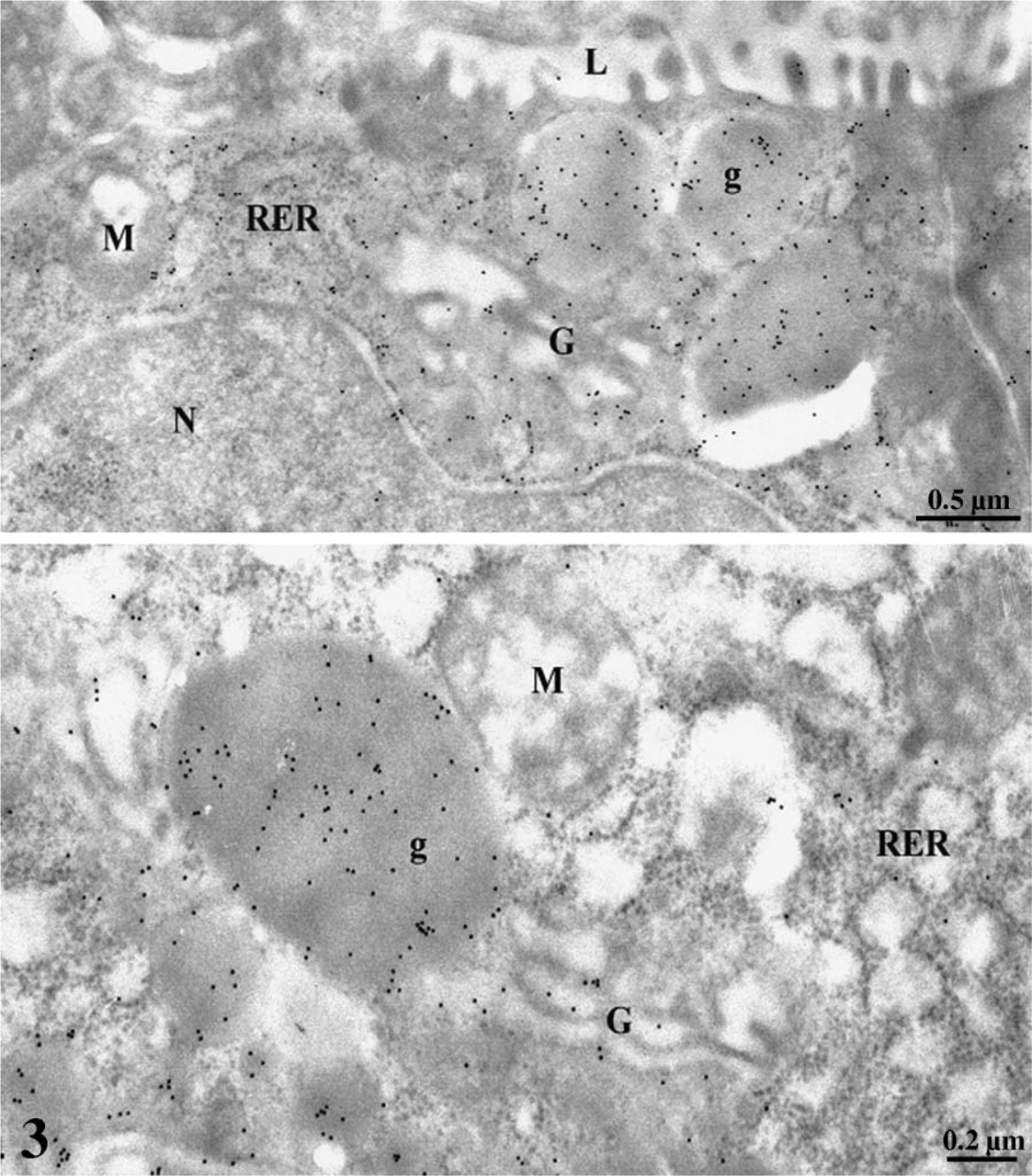
Immunogold demonstration of leptin in gastric Chief cells. The rough endoplasmic reticulum (RER), the Golgi apparatus (G), and the secretory granules (g) display a labeling by gold particles demonstrating the presence of leptin in these compartments. Mitochondria (M) and nucleus (N), as well as the gastric lumen (L), are rather devoid of specific labeling.
Discussion
The gastric mucosa has been shown to be involved in the regulation of food intake by producing hormones such as orexigenic ghrelin and anorexigenic leptin that act on the hypothalamus (Bado et al. 1998; Wynne et al. 2004). Plasma leptin is tightly correlated to the total amount of body fat in the preprandial state and increases quickly and potently after feeding or upon injection of cholecystokinin in rodents (Bado et al. 1998). Interestingly, fasting decreases circulating leptin and leptin secretion by white adipocytes, whereas it has no effect on the fundic content of leptin (Ballinger 1999). This observation suggests that, in addition to the production of leptin by white adipocytes, leptin originating from the gastric mucosa has an important role in the short-term regulation of food intake by participating in the rapid postprandial increase of circulating leptin. The present study has examined the secretion of leptin by the gastric mucosa and the presence of its receptor along the digestive tract.
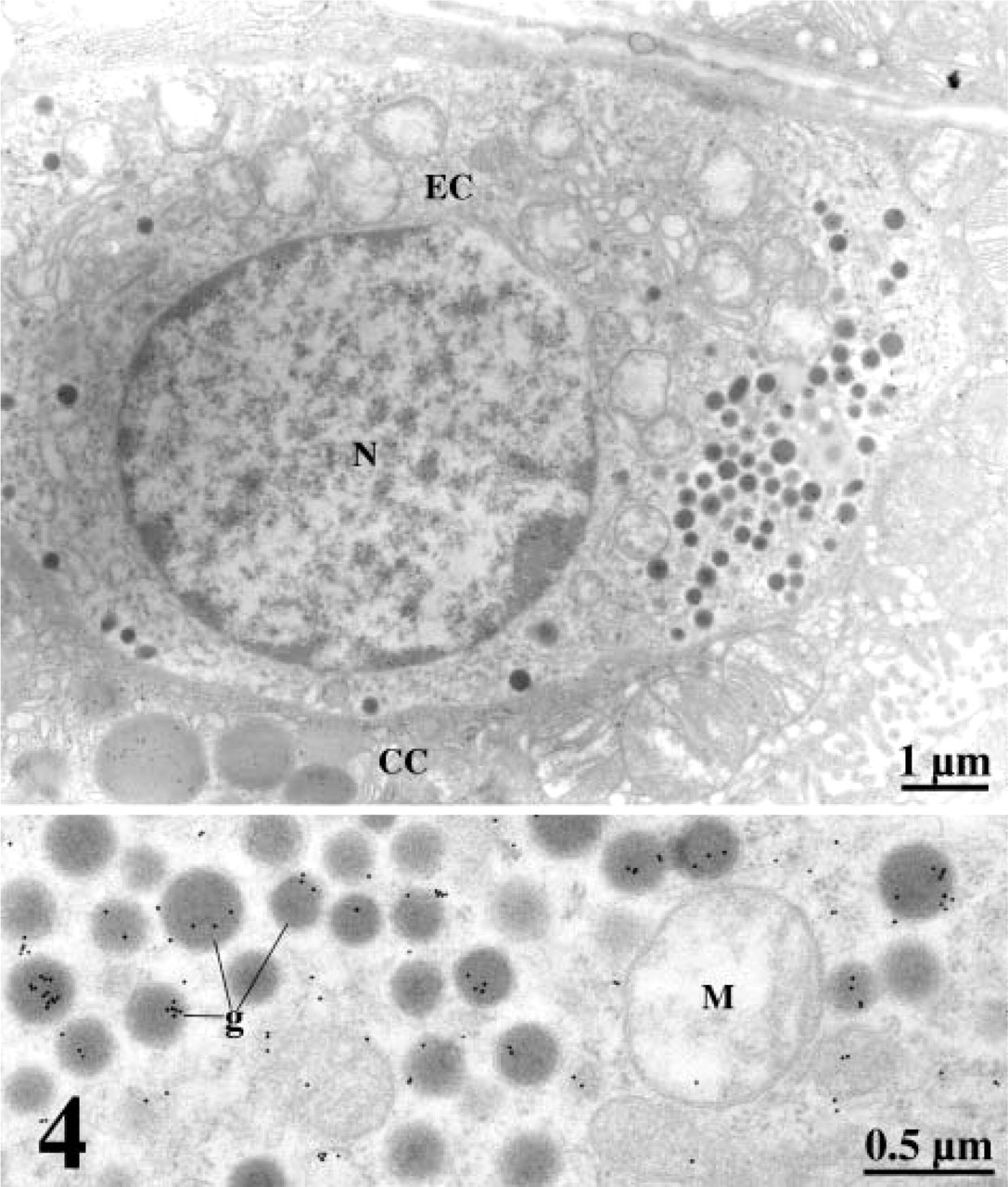
Immunolabeling of leptin in an endocrine cell (EC) of the rat gastric mucosa. Leptin was revealed in small dense granules (g). Labeling is evident at higher magnification. Nucleus (N), Chief cells (CC), Mitochondria (M).
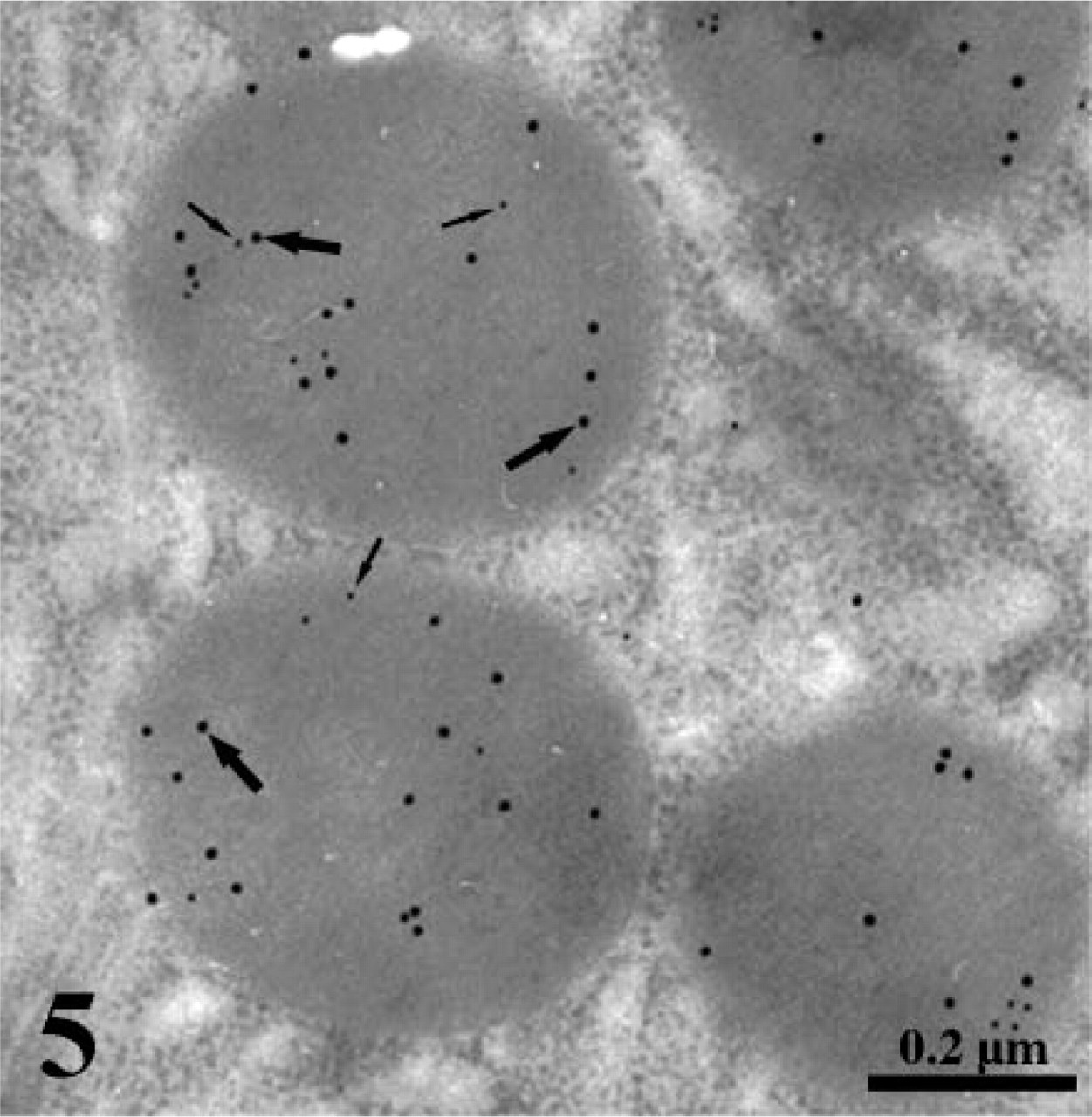
Double immunogold labeling of leptin (small arrows) and pepsinogen (large arrows) in secretory granules of a Chief cell. Anti-leptin and anti-pepsinogen incubations were followed by protein A-gold and anti-goat IgG-gold, respectively. Both antigens are colocalized in the same granules.
Our results have shown that the stomach secretes leptin through both an exocrine and an endocrine pathway. Immunostaining for leptin has demonstrated its presence in epithelial cells located in the lower half of the gastric mucosa. Leptin-positive cells were present all along the fundic mucosa of the rat, immediately after the cardia portion. In the fundic mucosa, two leptin-containing cell types were identified. The first consists of cells located in the gastric glands, the Chief cells positive for both leptin and pepsinogen. Parietal (oxyntic) cells were negative for leptin. Interestingly, colocalization of leptin and pepsinogen in the same secretory granules of the Chief cells suggests a rapid secretion of leptin into the gastric lumen after the onset of food intake, which is in accordance with a role of leptin in the short-term control of food intake.
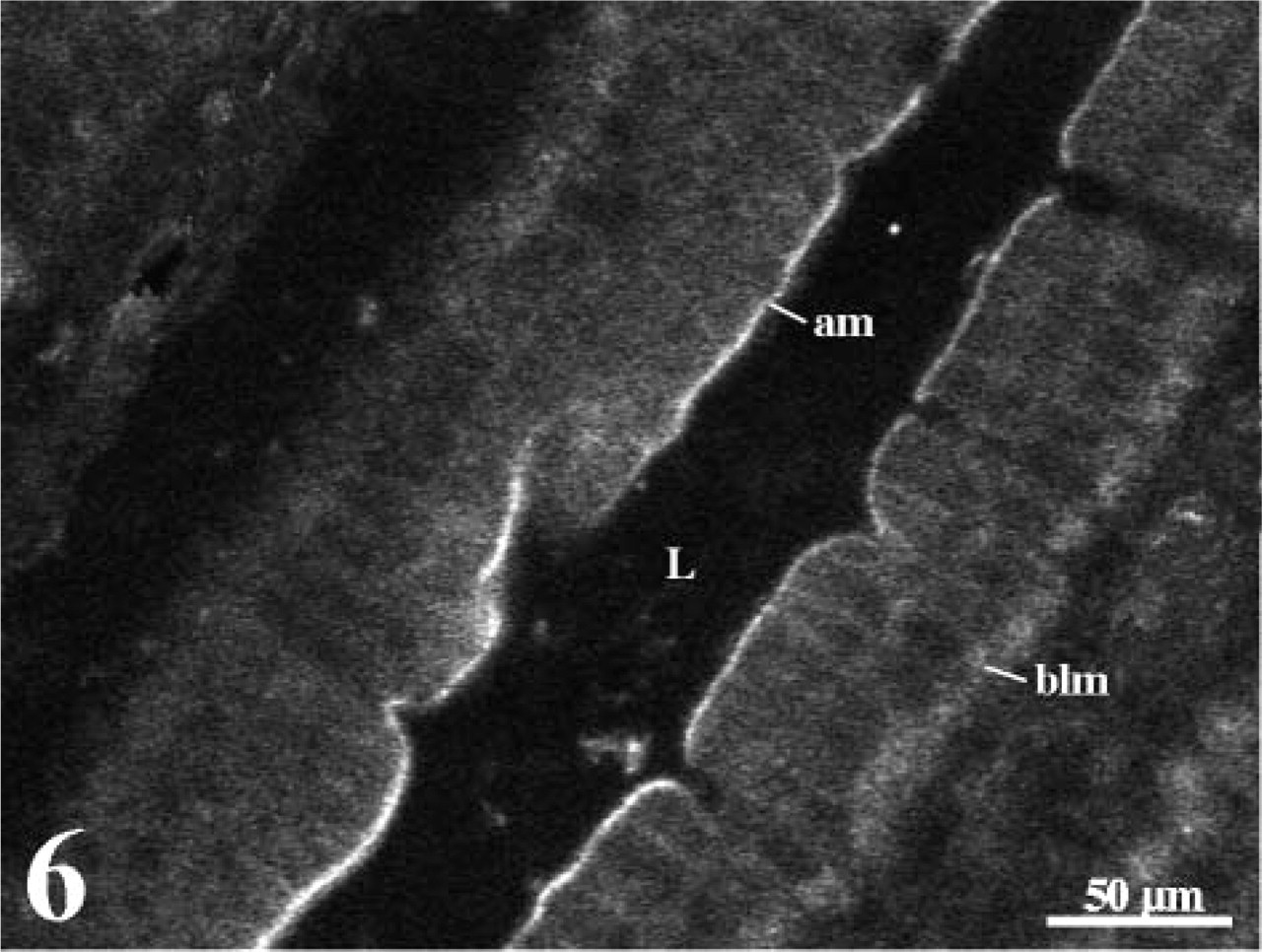
Immunostaining of leptin receptors in the rat duodenal wall. The tissue section was incubated with the anti-leptin receptor antibody followed by an anti-rabbit FITC-conjugated secondary antibody. Bright green fluorescence is present along the apical membrane (am). A lighter staining is also present on the basolateral portion (blm). No labeling is found in the lumen (L).
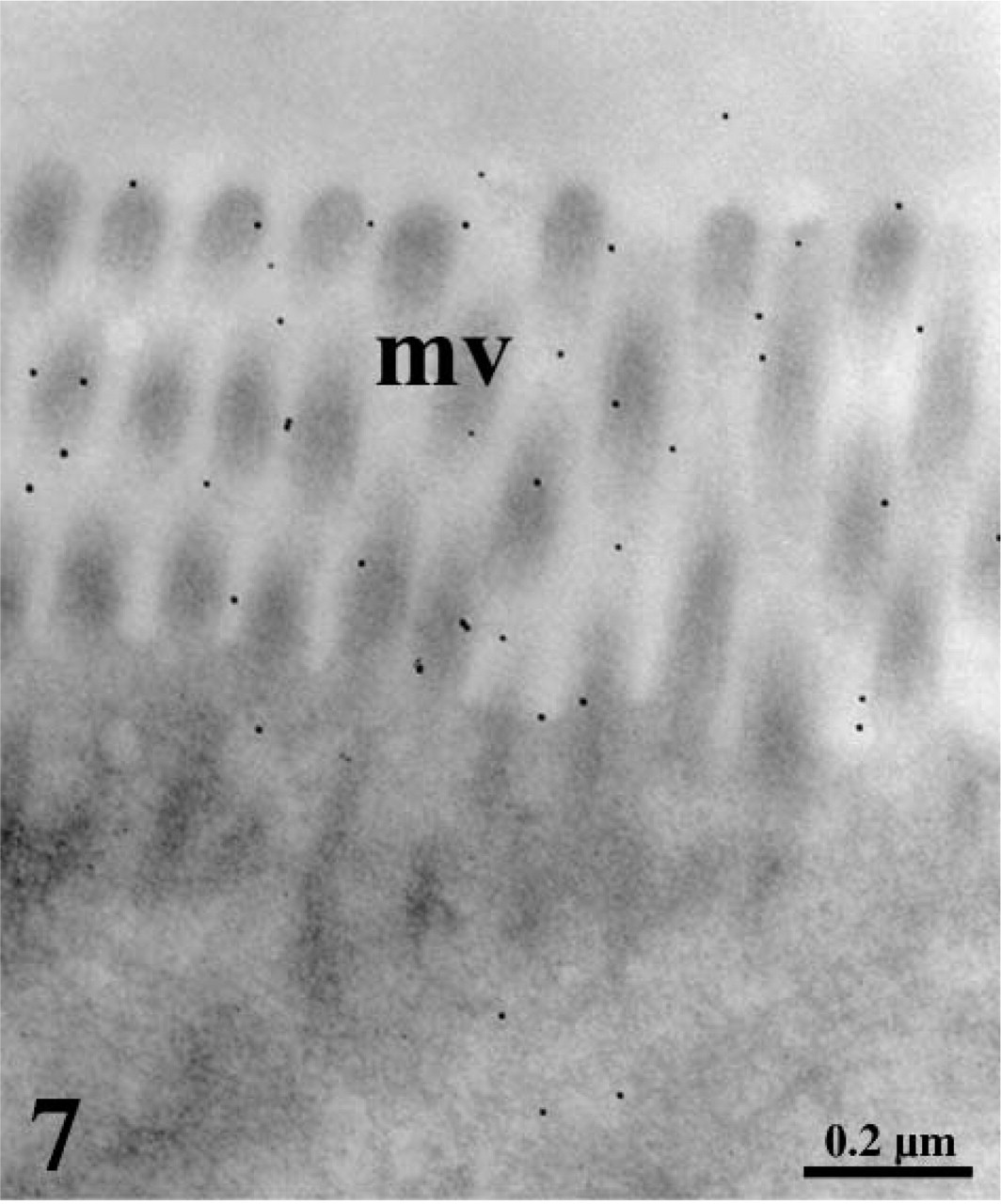
Ultrastructural localization of leptin receptors in duodenal enterocytes. The ultra-thin tissue section was incubated with the anti-leptin receptor antibody followed by protein A–gold. The microvilli (mv) display a labeling by gold particles demonstrating the presence of the receptor at the luminal surface.
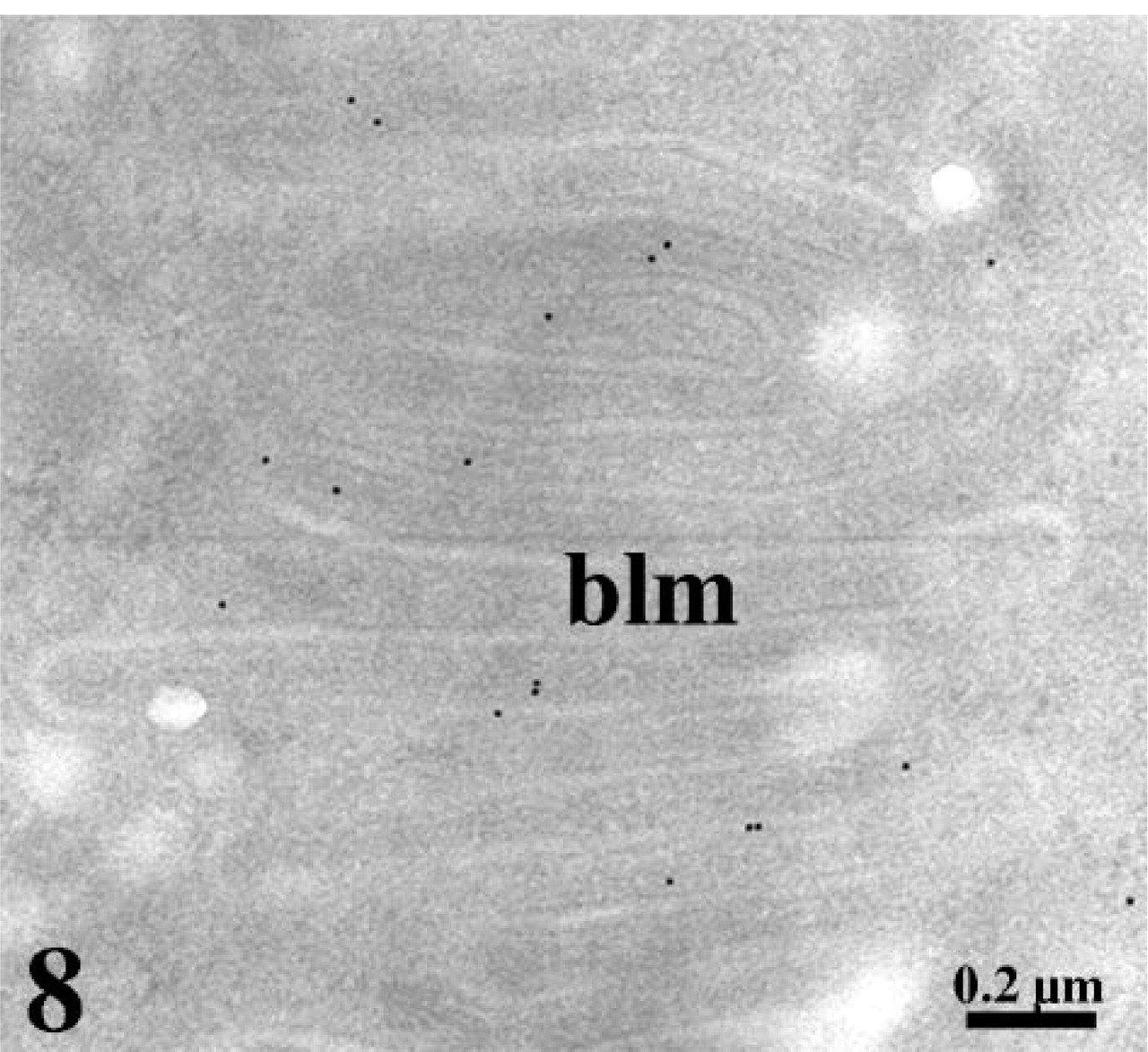
Ultrastructural localization of leptin receptors in duodenal enterocytes. The ultra-thin tissue section was incubated with the anti-leptin receptor antibody followed by protein A–gold. A weaker signal is detected at the level of the basolateral membrane (blm) of the duodenal enterocytes.
Ultrastructural studies showed significant labeling for leptin in the RER, increasing in the Golgi, and further in the secretory granules indicating that leptin is synthesized in situ and processed along the RER–Golgi-granule-regulated secretory pathway. Levels of leptin are higher in the granules than in the Golgi and higher in the Golgi than in the RER, suggesting that a concentration and/or maturation of a proleptin occurs in the Golgi before its final storage into secretory granules. Indeed, immunoblots of the gastric mucosa extracts have revealed two bands for leptin, one at 16 kDa and the other at 19 kDa, indicating the existence of a proform of leptin (Bado et al. 1998; Cinti et al. 2000).
The second type of leptin-containing cells consists of small cells, few in number, and located between the gastric glands. The staining in these cells was more intense than that of Chief cells, revealing greater concentrations of leptin. By electron microscopy, these cells showed numerous small secretory granules containing leptin and displayed the morphological characteristics of endocrine cells. Similar observations were reported by Cinti et al. (2000). Not all endocrine cells present in the mucosa were positive for leptin, underlining the existence of a specific type of endocrine cells for leptin. In these cells, the labeling was also detected in the RER and in the Golgi complex, suggesting a regulated type of secretion through the RER–Golgi-granule classical secretory pathway. Close association of these cells with capillaries strongly suggests release of leptin in the interstitial medium toward the bloodstream. This may well explain the rapid increase in circulating leptin observed in postprandial situations.
Leptin needs to bind to its membrane receptor to exert its effect. The presence of leptin in the gastric juice suggests that leptin receptors should be located in the luminal cell membrane along the digestive tract. Similarly, the presence of endocrine cells and the rapid rise in postprandial plasma levels must imply a paracrine action of gastric leptin on the gastric and intestinal tissues. We were unable to detect leptin receptors on gastric cell membranes on either the apical or on the basolateral one, although others have reported leptin receptors on Chief cell apical membranes (Mix et al. 2000; Sobhani et al. 2000). Discrepancies may arise from the techniques and specificity of the reagents used. Nevertheless, it has been shown that leptin acts on the stomach indirectly through the vagal nerve, triggering the release of acetylcholine. Similarly, leptin elicits release of intestinal hormones, which act as feedback regulators for leptin by binding their respective basolateral receptors (Bado et al. 1998; Sobhani et al. 2002). One of these hormones is cholecystokinin, which acts on gastric mucosa to enhance leptin release. It must be noted that white adipocytes do not secrete leptin in response to cholecystokinin, which suggests a targeted feedback regulation of leptin in the digestive tract (Ballinger 1999).
On the other hand, leptin receptors were found in the intestinal enterocytes, particularly on their brush border and on their basolateral membrane interdigitations at the level of duodenum, jejunum, and ileum. No major differences in receptor density were detected between the three portions of the small intestine. Previous studies on the localization of leptin receptors in intestinal tissue are scarce (Lostao et al. 1998; Barrenetxe et al. 2002) and did not report any staining at the enterocyte basolateral membranes. However, the presence of these receptors was indirectly indicated by studies on the effect of leptin on intestinal integrity and function. Leptin regulates absorption of nutrients including sugars, fat, and proteins. In vitro, leptin was shown to have an inhibitory effect on the intestinal uptake of galactose, whereas it has a reverse effect in vivo (Lostao et al. 1998; Pearson et al. 2001; Alavi et al. 2002). Leptin, acting on apical cell membranes but not on the basolateral ones, increases uptake of peptides by stimulating the recruitment of a specific transporter to the apical membrane (Buyse et al. 2001). It also regulates expression of proteins involved in lipid transport (Dube et al. 2001; Stan et al. 2001). Furthermore, apo A-IV synthesis is decreased by leptin in vivo (Liu et al. 2003). Luminal leptin is involved in the integrity of the gastric and intestinal mucosa and in their immune response (Morton et al. 1998; Brzozowski et al. 2000; Buyse et al. 2001; Konturek et al. 2001) and acts as a growth factor (Alavi et al. 2002).
It is interesting to compare pathways of synthesis and secretion of leptin by the gastric cells and the adipocytes. It appears that this hormone is secreted through a constitutive pathway by the white adipocytes and through a regulated pathway by the gastric Chief cells (Bado et al. 1998; Levy and Stevens 2001; Cammisotto and Bukowiecki 2004). For the gastric mucosa, leptin is processed through the RER–Golgi-granule-regulated secretory pathway, whereas no true secretory granule was recorded in adipocytes. Furthermore, leptin secretion by adipocytes is a continuous process that increases only 1 hr after stimulation. Also, obese animals exhibit a marked increase of their adipose tissue, with a concomitant increase in leptin content and in circulating leptin, whereas gastric amount of leptin for the same obese animals shows no changes (Ballinger 1999). All these characteristics indicate a rather constitutive secretion of leptin by adipocytes. These differences in secretory patterns support a different regulation of the ob gene expression by the gastric mucosa and by adipose tissue.
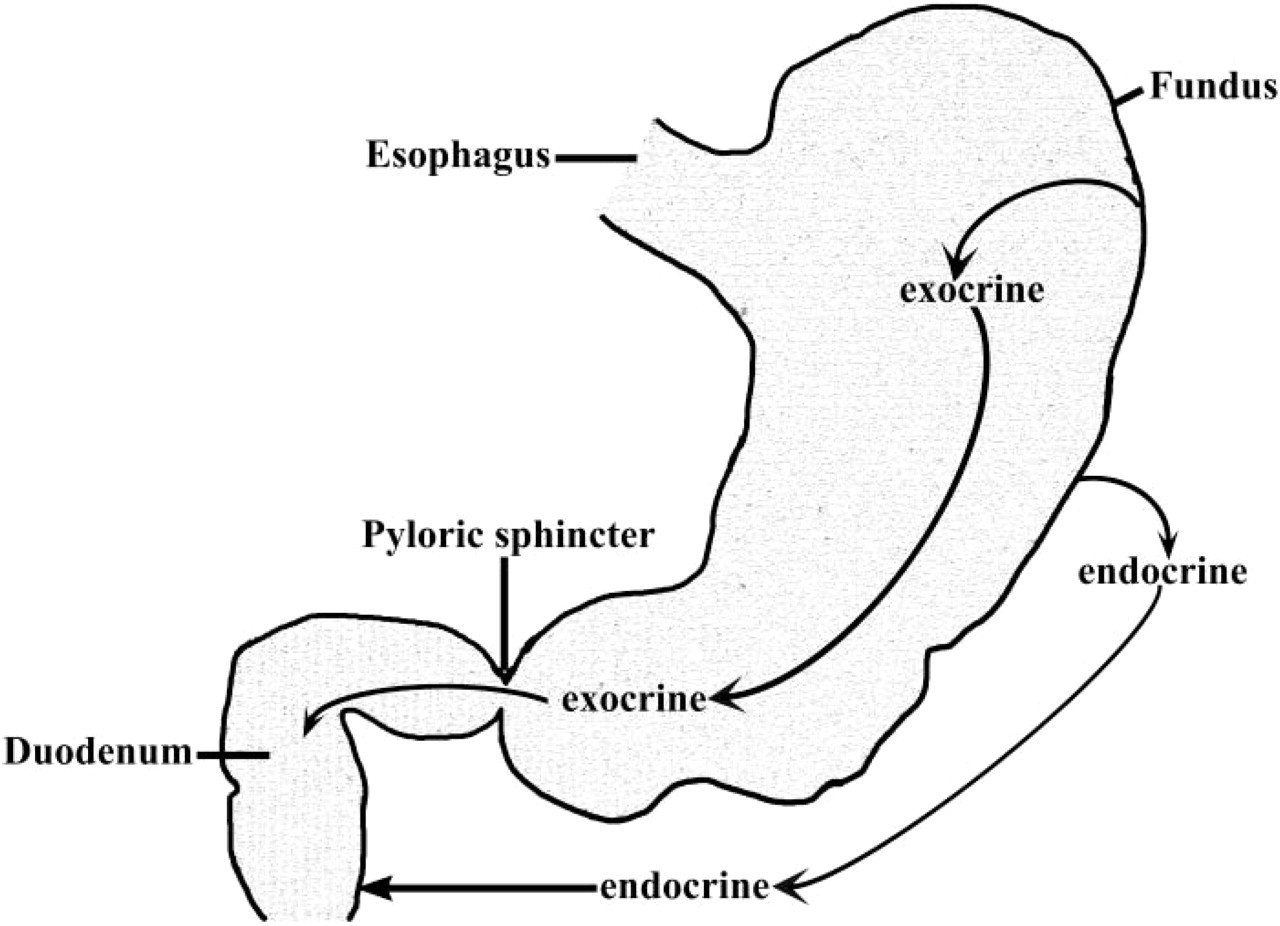
Summary of leptin secretion and site of leptin action in the digestive tract. Leptin is secreted by exocrine Chief cells into the gastric juice and by endocrine cells toward the bloodstream. Exocrine and endocrine leptin act on intestinal leptin receptor located, respectively, on the apical and basolateral plasma membranes.
Taken together, these observations highlight the existence of multiple secretions of leptin. Exocrine leptin is secreted into the gastric juice together with pepsinogen shortly after the beginning of food intake and acts on the luminal membrane of intestinal enterocytes to regulate their function. On the other hand, endocrine gastric secretion of leptin may be responsible for a paracrine action rapidly acting on leptin receptors located at the basolateral membranes of intestinal cells. These observations are summarized in Figure 9. Parallel to this, adipocyte leptin contributes to circulating levels that also act at long term on target tissues including enterocytes. Further studies will be needed to understand the respective roles of exocrine and endocrine secretions on the digestive tract in the postprandial state. It will also be interesting to determine the fate of leptin and of its binding protein within the intestinal mucosa as many peptides and proteins bound to chaperones have been shown to be internalized by enterocytes and to be delivered to the bloodstream (Bruneau et al. 2000).
Footnotes
Acknowledgements
This work was supported by a grant from the Canadian Institutes for Health Research.