
Abstract
We report on a balanced complex chromosomal aberration detected in a fetus after amniocentesis. The pregnancy was achieved after intracytoplasmic sperm injection. GTG-banding revealed a complex structurally rearranged karyotype with a translocation between chromosomes 5 and 15 and an additional paracentric inversion in the der(15) between bands 5q11.2 and 5q15. Ag-NOR staining showed an interstitial active nuclear organizer region in the der(15). Molecular cytogenetic analyses using whole-chromosome–painting probes, comparative genomic hybridization, and multicolor banding did not point to further structural aberrations or imbalances. Therefore, a complex rearrangement with three breakpoints has occurred, and the karyotype can be described as 46,XX,der(5)t(5;15)(q11.2;p12),der(15)t(5;15)(q11.2;p12)inv(5)(q11.2q15).
C
A 40-year-old woman was referred for prenatal diagnosis and subsequent karyotyping of amniocytes because of assisted fertilization, maternal age, and a 4-mm choroid plexus cyst. The parents had opted for ICSI after a spermiogram revealed asthenoteratozoospermia with slightly decreased sperm number and explicitly reduced sperm motility, as well as conspicuous sperm morphology. The cause of the abnormal sperm morphology and motility could not be identified, and the karyotypes of both partners were normal at a resolution level of 400 bands per haploid chromosome set in lymphocytes.
Karyotyping of GTG-banded chromosomes from cultured amnion cells at the 18th week of pregnancy revealed a structurally rearranged karyotype with a derivative chromosome 5 consisting mainly of chromosome 5p material and a derivative chromosome 15 apparently composed of nearly the whole chromosome 15 and a rearranged part of the long arm of chromosome 5, presumably an inversion (Figure 1A). This aberrant karyotype was found in all metaphases analyzed from two independent cultures, indicating that the constitutional aberrant karyotype was present in the fetus. There were no hints of mosaicism in 50 cells analyzed. Staining the nuclear organizer regions (NORs) indicated that the der(15) contained interstitial active NORs (Figure 1B). After GTG-banding the karyotype appeared to be balanced, on the basis of 400 bands per haploid genome, and hence has been described as 46,XX,der(5)t(5;15)(q11.2;p12),der(15) t(5;15)(q11.2;p12)inv(5)(q11.2q15).
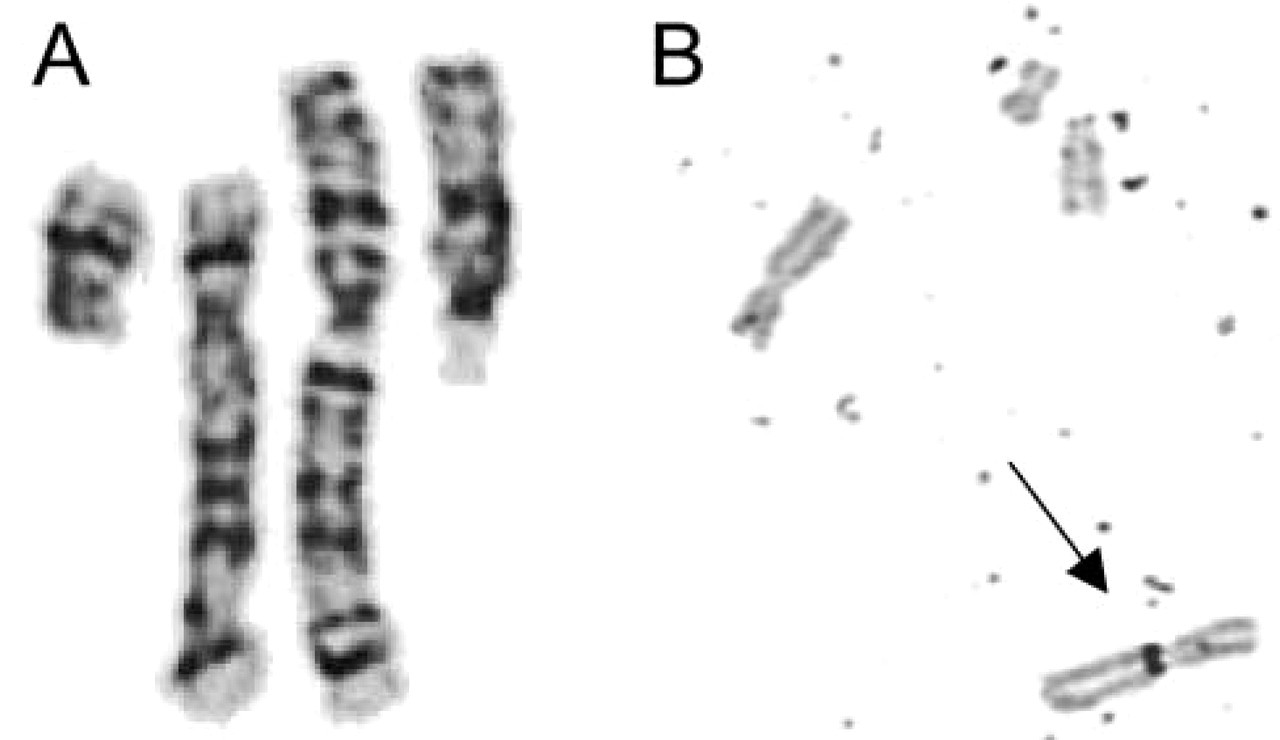
GTG-banding of the chromosomes involved in the rearrangement
Because conventional cytogenetics alone was not sufficient to elucidate the rearrangement with satisfactory accuracy, we performed additional molecular cytogenetic analyses to define the exact composition of the derivative chromosomes. An even more complex rearrangement was excluded by hybridizing chromosome-painting probes for chromosomes 5 and 15 (Qbiogene; Carlsbad, CA) following the manufacturer's instructions. No euchromatin of these two autosomes could be detected on any other chromosome, and no interruption of the hybridization signal was visible on the two derivative chromosomes (Figure 2A). Therefore, no other chromosome seemed to be involved in the rearrangement. This was confirmed by comparative genomic hybridization (CGH) performed as described elsewhere (Tönnies et al. 2001). CGH did not hint at duplications or deletions at the breakpoints or at any other chromosomal imbalances, which cannot be excluded by hybridizing whole-chromosome paints alone (Figure 2B). Hybridization and comparison of the relative positions of locus-specific yeast artificial chromosome clones on the der(15) and the normal chromosome 5 pointed toward the existence of a paracentric inversion of chromosome 5q material in the der(15) (Figure 2C).
This was confirmed by multicolor banding (MCB) (Figure 2D). Hybridization of probes specific for subcentromeric regions of chromosome 5 showed normal signals on the der(5) (Figure 2D). MCB and subcentromere-specific multicolor fluorescence in situ hybridization (subcenM-FISH) were done as described previously (Chudoba et al. 1999; Liehr et al. 2002; Starke et al. 2003). All applied molecular cytogenetic techniques supported the assumptions of the conventional cytogenetic analysis.
To exclude a uniparental disomy (UPD) due to trisomy rescue after a possible 3:1 segregation with interchange trisomy for chromosome 15, we extracted genomic DNA from the amniocytes by standard procedures and performed methylation testing on the imprinting center of the Prader-Willi syndrome (PWS)/Angelman syndrome (AS)–critical region, as described by Zeschnigk et al. (1997). Methylation-specific PCR revealed two bands for the fetal DNA and consequently excluded a UPD 15 for the PWS/AS critical region (Figure 2E).
A very rare complex chromosomal aberration was diagnosed after chromosome analysis of cultured amniocytes. Conventional and molecular cytogenetic analysis showed a balanced karyotype [46,XX,der(5)t(5;15)(q11.2;p12),der(15)t(5;15)(q11.2;p12)inv(5)(q11.2q15)]. The risk for phenotypic abnormalities associated with a de novo reciprocal two-breakpoint translocation is estimated to be 6%, 3% above the background risk of about 3% for malformations and/or defects in all pregnancies (Warburton 1991). The risk estimation in Warburton's study is slightly higher for balanced two-breakpoint inversions (9.4%) but probably merely due to a smaller case number. De novo, apparently balanced CCRs have presumably a high risk for mental retardation and malformations. The few postnatal studies suggest that the risk for abnormal phenotypes increases with a greater number of breakpoints (Madan et al. 1997). In our case, the exact molecular cytogenetic characterization of the rearrangement did not point toward any imbalances. The rearrangement involves only three breakpoints that were defined by cytogenetic analysis: the breakpoint at 5q11.2 is involved in two events, the inversion and the translocation. Furthermore, the breakpoint in chromosome 15 localizes in the short arm of the chromosome, a region containing no relevant single-copy genes, but genes coding for rRNA. Thus, the risk for fetal malformations and developmental or mental retardation should not exceed the 9.4% given for the usual two-breakpoints inversion, although it concerns a complex rearrangement. Non-acrocentric chromosomes with interstitial NORs derived by translocations or insertions from acrocentric chromosomes are rare forms of chromosomal rearrangements without phenotypic effect, if there is no loss of euchromatic material from the non-acrocentric chromosome (Willatt et al. 2001).
Accurate definition of the chromosomal breakpoints was essential to enable proper genetic counseling. After genetic counseling and additional inconspicuous ultrasound examination, the parents decided to continue with the pregnancy; meanwhile, a healthy girl was born, with a birth weight of 3.005 g and a birth length of 49 cm.
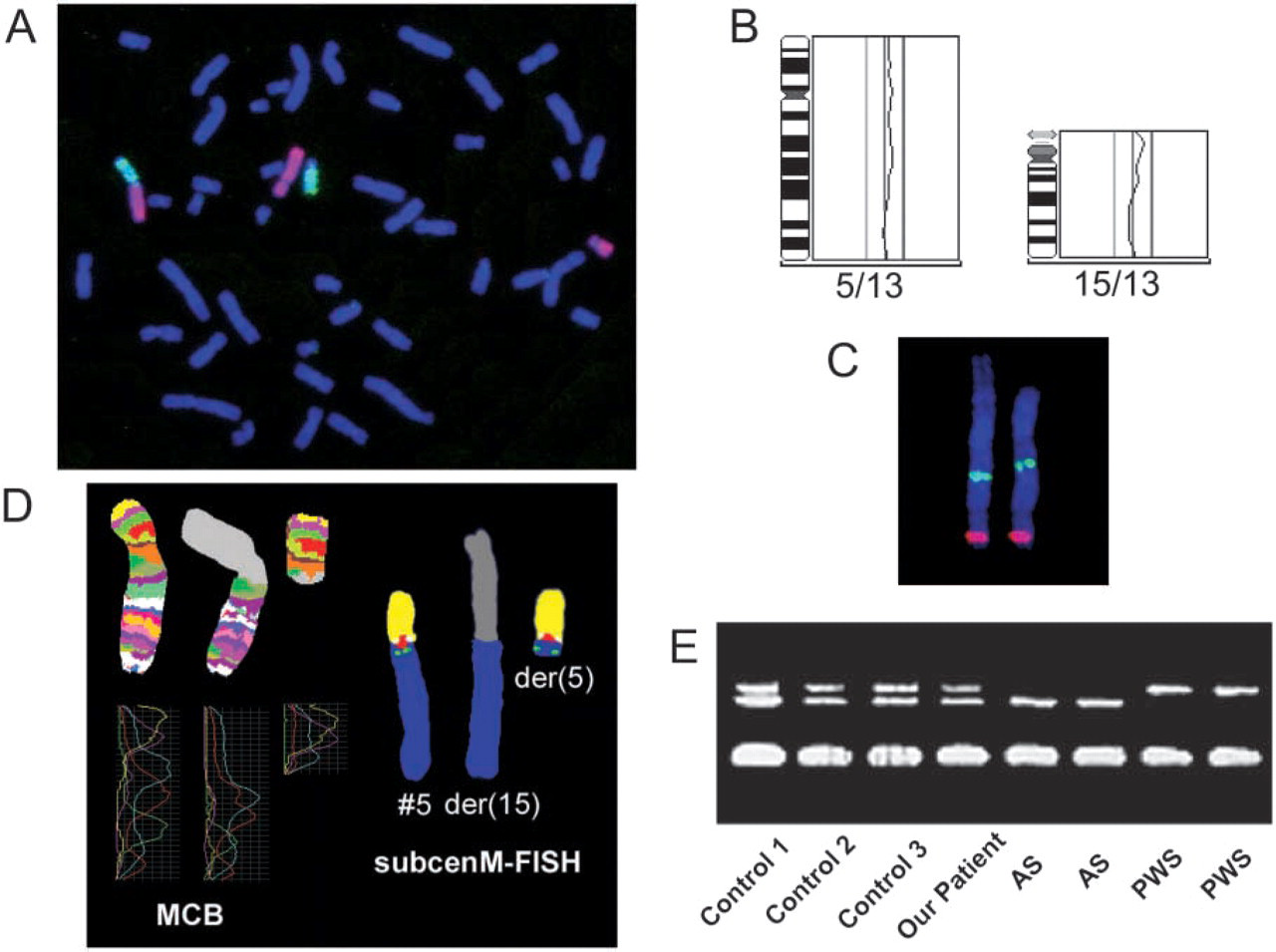
Chromosome painting of chromosomes 5 (red) and 15 (green) did not point toward an involvement of additional chromosomes in the rearrangement
The chromosomal constitution will have considerable implications for the girl in the event of of a possible wish for having her own children in the future. If the rearrangement does not cause a general meiotic arrest, two segregant gametes may be produced by the translocation heterozygote that could lead to viable, severely affected offspring. Tertiary trisomy including the der(5) following 3:1 segregation results in trisomy for the whole short arm of chromosome 5, which is associated with severe malformations and mental retardation. Over 40 cases with partial trisomy of 5p have been reported to date (Grosso et al. 2002). The corresponding tertiary monosomy would result in a constellation with a cri-du-chat syndrome, a well-described partial monosomy resulting from deletion of the short arm of chromosome 5. A trisomy rescue after interchange trisomy for chromosome 15 would cause a PWS due to maternal UPD 15 in prospective offspring of the girl, while monosomy rescue of the paternal chromosome 15 following the corresponding interchange monosomy of chromosome 5 would result in an Angelman syndrome. Recombination products following crossing over in a meiotic inversion loop would lead to dicentric recombinant chromosomes or acentric fragments. These are lethal constellations in early embryonic development and therefore do not enhance the risk for abnormal offspring.
The findings could also have implications for further pregnancies of the couple itself, if one of the partners had a gonadal mosaicism. However, the specified unbalanced segregation products could be easily identified by karyotyping fetal cells in further pregnancies. Exclusion of a UPD 15 would require molecular genetic analysis of fetal DNA.
Although most familial complex rearrangements are transmitted through the mother, paternal transmission has been described in a few cases (Grasshoff et al. 2003; Gardner and Sutherland 2004). Infertility in male carriers of complex translocations is well known and the abnormal spermiogram could possibly be due to gonadal mosaicism for the rearrangement. A potential gonadal mosaicism—if existing in the father—could be verified by analyzing segregation patterns in sperm analysis using fluorescence in situ hybridization.
Footnotes
Acknowledgements
TL was supported in part by the EU (ICA2-CT-2000-10012).
The authors thank Antje Gerlach and Britta Teubner for excellent technical assistance and Trudy Hocking and Sharon Bain for discussing the ISCN nomenclature.