
Abstract
1,25-dihydroxyvitamin D3 has anti-mitotic, pro-differentiating, and pro-apoptotic activity in tumor cells. We demonstrated that the secosteroid can be synthesized and degraded not only in the kidney but also extrarenally in intestinal cells. Evaluation of 1,25-dihydroxyvitamin D3-synthesizing CYP27B1 hydroxylase mRNA (real-time PCR) and protein (immunoblotting, immunofluorescence) showed enhanced expression in high- to medium-differentiated human colon tumors compared with tumor-adjacent normal mucosa or with colon mucosa from non-cancer patients. In high-grade undifferentiated tumor areas expression was lost. Many cells co-expressed CYP27B1 and the vitamin D receptor. We suggest that autocrine/paracrine antimitotic activity of 1,25-dihydroxyvitamin D3 could prevent intestinal tumor formation and progression. (
Keywords
1,25-
An adequate 1,25-D3 serum level is essential to maintain calcium homeostasis. Because of the hormone's anti-mitotic, pro-differentiating, pro-apoptotic activity, it has also been suggested to provide protection against tumor progression (Lamprecht and Lipkin 2003). However, treatment of tumor patients with 1,25-D3 or with synthetic vitamin D analogues is not feasible due to the hypercalcemic effect at the pharmacological (nanomolar) doses necessary to achieve antimitotic action. Recently, another physiological link between vitamin D and cancer prevention and/or therapy has been found. In addition to the kidney, several other tissues express enzymes necessary for vitamin D synthesis. We were the first to demonstrate that human colonocytes in culture are able to synthesize 1,25-D3 from 25-D3 (Cross et al. 1997) and that in freshly isolated colon tumor cells a wide spectrum of vitamin D metabolites is present (Bareis et al. 2001). Our further studies by semiquantitative RT-PCR showed increasing levels of CYP27B1 and of VDR mRNA during early colon tumor progression (Cross et al. 2001). CYP27B1 mRNA was overexpressed also in cervical, breast, and ovarian carcinomas compared with normal tissue (Friedrich et al. 2003). This suggested to us an autocrine/paracrine protective effect of 1,25-D3 synthesized in colon tumor cells during premalignancy and early malignancy. In late-stage high-grade colon cancer there is apparent failure of this protective system (Cross et al. 2001). However, with present methodology it is not possible to verify this by measuring tissue accumulation of 1,25-D3.
In view of our preliminary observations and the fact that the expression level of CYP27B1 seems to be related to the level of cell differentiation, at least in human colon tumor cell cultures (Bareis et al. 2002), evaluation of the exact site of CYP27B1 expression within single tumors appears to be essential. Therefore, we investigated the localization of CYP27B1 in colorectal tumor and, in parallel, quantified its expression by real-time PCR and by immunoblotting. All tumors were adenocarcinoma type and were graded according to WHO criteria (Jass and Sobi 1989). For real-time RT-PCR and immunoblotting, we analyzed only highly or moderately differentiated tumors (10 randomly chosen patients with G1/G2 colon adenocarcinomas that were pT1, pT2 and pT4 one each, and seven were pT3), and normal tumor-adjacent mucosa from the same patient. Non-cancerous colon tissues were obtained from diverticulitis patients after stoma re-operation. Permission from the local Ethics Commission was obtained.
Real-time RT-PCR was performed on an ABI Prism 7700-Sequence Detection System (Applied Biosystems; Foster City, CA). We quantified expression of CYP27B1 mRNA by the comparative ΔΔCT method using cytokeratin 8 (CK 8), an invariant marker of simple epithelia as reference gene and a cDNA stock from Caco-2 cells as calibrator.
Equal amounts (1 μg) of total RNA were reverse-transcribed for single-strand cDNA using SuperScript II (Invitrogen; Groningen, The Netherlands). Primers and TaqMan probes were designed with Primer Express (Applied Biosystems) and are located on different exons to prevent amplification from contaminating genomic DNA. CYP27B1: forward, 5′-AGTTGCTATTGGCGGGAGTG-3′; reverse, 5′-GTGCCGGGAGAGCTCATACA-3′; probe: 5′-ACACGGTGTCCAACACGCTCTCTTGG-3′; CK 8: forward, 5′-GATCTCTGAGATGAACCGGAACA-3′; reverse, 5′-GCTCGGCATCTGCAATGG-3′; probe: 5′-CTCAAAGGCCAGAGGGCTTCCCTG-3′.
Immunoblotting was performed as described previously (Bareis et al. 2002). Briefly, total protein was prepared by homogenizing tissue samples in 1 ml boiling lysis buffer (10 mM TRIS, pH 7.4, with 1% SDS) and was separated by 12% SDS-PAGE and blotted to a nitrocellulose membrane. A sheep anti-CYP27B1 (The Binding Site; Heidelberg, Germany) and horseradish peroxidase (HRP)-conjugated anti-sheep IgG (Sigma-Aldrich; St Louis, MO) were used to detect CYP27B1 protein levels. CK 8, the internal control, was detected with a mouse anti-CK 8 antibody (Cymbus Biotechnology; Chandlers Ford, UK) and HRP-labeled anti-mouse IgG.
For immunofluorescence analysis, we investigated colon tissue samples derived from 30 patients aged 41–87 years (equal gender distribution) whose colon mucosa had premalignant to highly malignant lesions [two polyps, 12 adenomas, eight G2 tumors (two pT1, two pT2, three pT3, one pT4), eight G3 tumors (two pT2, three pT3, and three pT4)] and compared these with normal mucosa. Kidney sections were used as positive control.
We deparaffinized and rehydrated 5 μm paraffin-embedded human tissue sections. Antigen retrieval was performed with citrate buffer (10 mM, pH 6.0) at 600 W three times for 5 min in a microwave oven. Sections were permeabilized in PBS/0.2% Tween-20 and blocked with 5% rabbit serum in PBS/0.05% Tween-20. Sheep anti-CYP27B1 (The Binding Site) diluted 1:150 in PBS/0.05% Tween-20 was applied for 1 hr at room temperature. Negative controls were: sheep IgG 60 μg/ml (Jackson ImmunoResearch Laboratories; West Grove, PA) or the CYP27B1 antibody preabsorbed with the specific immunogen. After extensive washing in PBS/0.05% Tween-20, Cy3-conjugated rabbit anti-sheep IgG (1:10,000) (Jackson) was applied for 1 hr at RT. After washing the sections were mounted in Vectashield medium (Vector Laboratories; Burlingame, CA) and viewed in a fluorescence microscope Nikon Eclipse E400.
For double immunostaining, sections were treated as described above. First staining was performed with rat anti-VDR (Chemicon; Temecula, CA) diluted 1:50 overnight at 4C. IgG of the same isotype (Jackson) was used as negative control at a concentration of 1 μg/ml. FITC-conjugated rabbit anti-rat secondary antibody (Jackson) was applied at a concentration of 1:400 for 1 hr at RT. The slides were washed in PBS/0.02% Tween-20 and nonspecific sites were blocked again with 5% rabbit serum for 30 min at RT before the second staining. The second primary antibody sheep anti-CYP27B1 (The Binding Site) and Cy3-conjugated rabbit anti-sheep secondary antibody were applied as described above.
Expression of CYP27B1 was semiquantitatively evaluated in each tumor and in the adjacent normal mucosa from the same patient (++++ = strong expression, +++ = medium expression, ++ = low expression, + = barely detectable expression).
Student's t-test was used to evaluate the outcome of the real-time PCR and immunoblotting experiments. Differences were considered significant at p <0.05.
When expression of CYP27B1 mRNA relative to CK 8 was evaluated, nine of ten well- to moderately differentiated tumors (G1/G2) had higher expression than the adjacent normal mucosa from the same patient (Figure 1A). We referred CYP27B1 mRNA to the epithelial cell marker since tumor samples contain variable amounts of epithelial cells and, according to our immunofluorescence data, colon CYP27B1 was expressed almost exclusively in epithelial cells. CYP27B1 mRNA levels were significantly elevated in tumor samples compared with paired normal adjacent mucosa (p <0.05) or with mucosa from non-cancer patients (p <0.01) (Figure 1A). Table 1 shows the collective mean ΔCT values for CK 8 and CYP27B1 normalized to 18S and, for CYP27B1, also normalized to CK 8. These data confirm the upregulation of CYP27B1 mRNA levels and show downregulation of CK 8 mRNA levels in tumor tissue. The collective mean ΔCT value for CK 8 in normal mucosa from non-cancer patients was significantly different from the mean levels both in adjacent mucosa and in tumor tissue (p <0.001). The average ΔCt value for CYP27B1 in tumors was significantly decreased (corresponding to increased mRNA expression) compared with either normal or adjacent mucosa, although the differences were more significant when CYP27B1 was normalized to CK 8.
Immunoblotting analysis showed similar results. CYP27B1 protein expression level was significantly higher in tumor tissue (p <0.05) compared with adjacent mucosa or with normal mucosa (Figure 1B).
Raw average ΔCT values
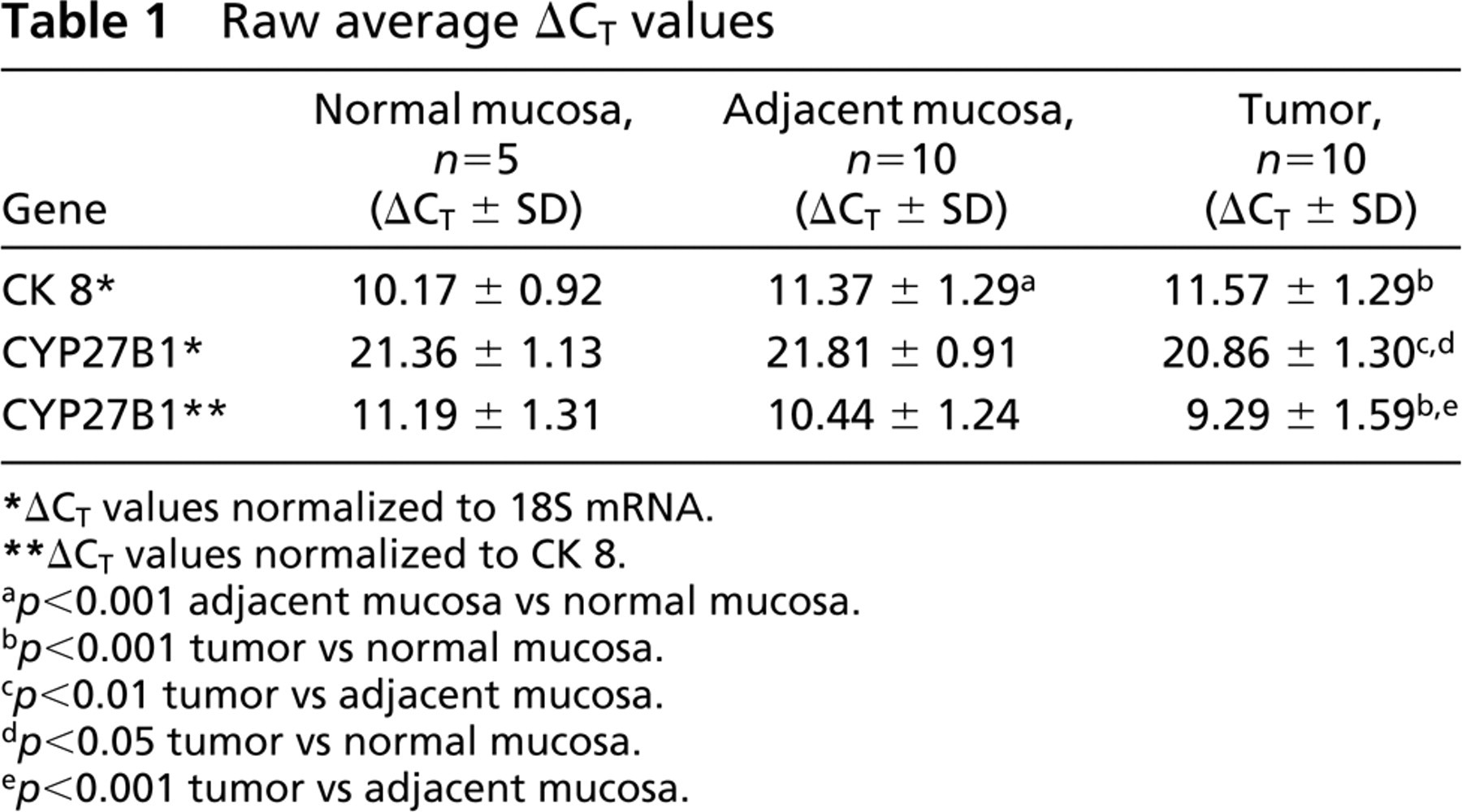
ΔCT values normalized to 18S mRNA.
ΔCT values normalized to CK 8.
p > 0.001 adjacent mucosa vs normal mucosa.
p > 0.001 tumor vs normal mucosa.
p > 0.01 tumor vs adjacent mucosa.
p > 0.05 tumor vs normal mucosa.
p > 0.001 tumor vs adjacent mucosa.
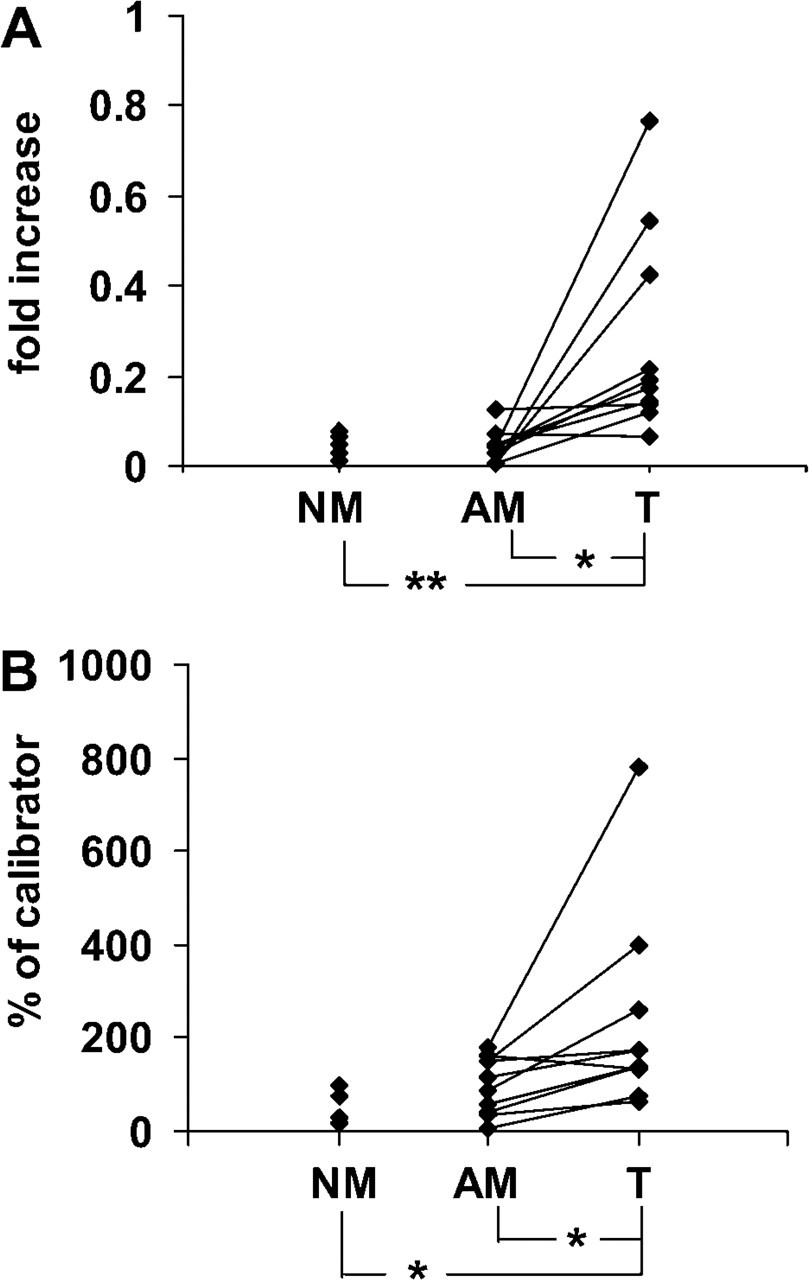
Expression of CYP27B1 mRNA and protein in normal colon mucosa from non-cancer patients (NM), mucosa adjacent to tumor border (AM) and colon tumors (T). (
Immunofluorescence analysis demonstrated that, as expected, distal tubules in the kidney strongly expressed CYP27B1, whereas proximal tubules had weaker and more variable expression (Figure 2A). In the non-cancerous colon the mucosa showed barely any positivity (Figure 2C), corresponding with the low CYP27B1 levels detected by real-time RT-PCR and immunoblotting. In colon adenomas we found intense staining for CYP27B1, and this seemed typical for well-differentiated tumors (Figure 2D). The high expression was maintained also in moderately differentiated (G2) adenocarcinomas (Figure 2E). Analyzing the simultaneous expression of CYP27B1 (red) and of VDR (green), we found many cells co-expressing these proteins (see Figure 2E inset). In high-grade (G3) tumors, CYP27B1 positivity was conspicuously absent in undifferentiated areas (Figure 2F), whereas in more differentiated tubular structures, even G3 tumors displayed considerable positivity (Figure 2G). Staining with sheep IgG (Figure 2B) or with the antibody blocked with the immunogenic peptide (not shown) was always negative.
Table 2 contains a detailed description of the immunohistochemical evaluation of all colon tumor samples. In premalignant tissues, such as hyperproliferative polyps and adenomas, and in low-grade tumors, CYP27B1 expression was elevated in 50% of patients, whereas the majority of normal adjacent mucosa outside low-grade tumors was negative. High-grade tumors (G3) are heterogeneous with respect to tubular structure. In areas with complete loss of differentiation, i.e., of a solid or trabecular pattern, CYP27B1 expression was no longer present. In two patients we observed increased CYP27B1 positivity, but only in areas with a rudimentarily conserved tubular pattern. The adjacent mucosa was clearly positive in 4 of 8 patients.
The immunofluorescence analysis supports our previous data (Bareis et al. 2001; Cross et al. 2001) that CYP27B1 expression is elevated in tumor tissue compared with normal colon mucosa during early stages of colon tumor progression. However, high-grade undifferentiated adenocarcinomas are devoid of CYP27B1 expression except in areas with preserved differentiated structures.
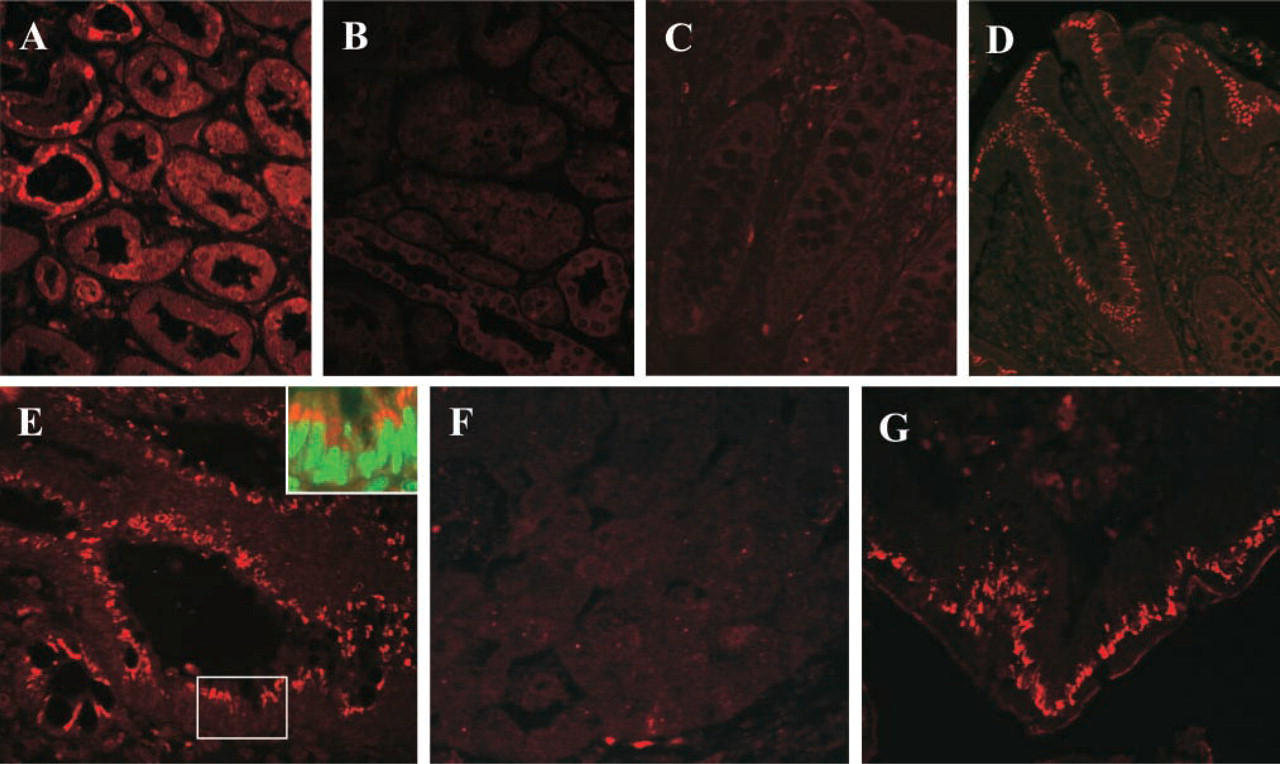
Localization of CYP27B1 (red) by immunofluorescence in (
Our real-time PCR data differ from those presented by Tangpricha et al. (2001) and Ogunkolade et al. (2002). They did not observe a rise in CYP27B1 expression in tumors, possibly because they did not consider the biological grade of the tumor (Tangpricha et al. 2001). We also suggest that well-advanced tumors contain fewer epithelial cells. Since, according to our immunofluorescence data, CYP27B1 is expressed almost exclusively in epithelial cells in the colon, we used CK 8 as internal control when we evaluated whole tissue pieces by real-time RT-PCR and by immunoblotting. Using a ubiquitous cellular marker such as actin instead may lead to underestimation of CYP27B1 expression, especially in tumor tissue (Ogunkolade et al. 2002).
During early tumor progression we previously observed a parallel increase of VDR and of CYP27B1 mRNA (Cross et al. 2001). When we double stained colon tumors for CYP27B1 and VDR, many colonocytes expressed both CYP27B1 and VDR (Figure 2E inset), although some cells were positive only for VDR. This suggests that 1,25-D3 synthesized in colonocytes and bound to its receptor could exert its anti-mitotic function in both an autocrine and a paracrine fashion.
Evaluation of CYP27B1 expression in human colon tissue by immunofluorescence
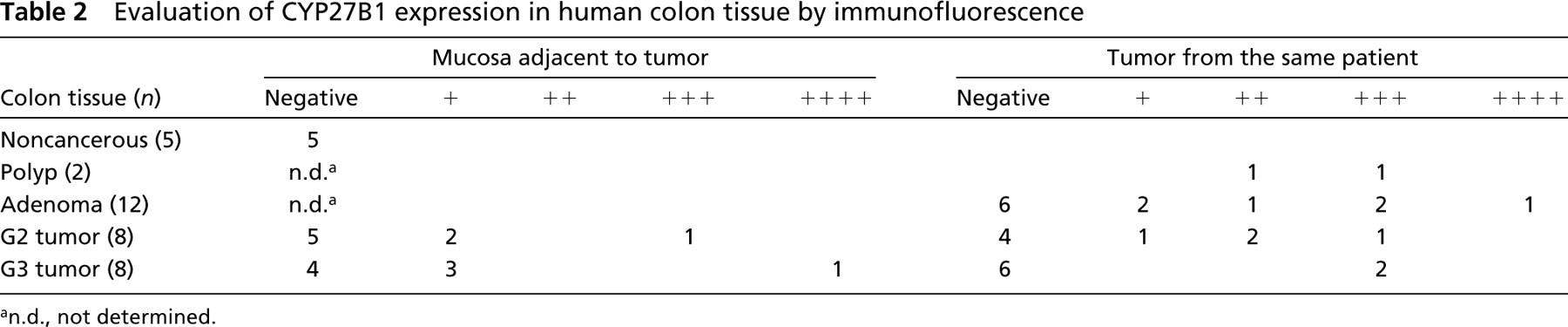
n.d., not determined.
In view of results provided by Evans et al. (1998), who demonstrated that patients with enhanced levels of VDR in their colon tumors have more benign outcome of the disease, we conclude that enhanced local synthesis of 1,25-D3 during early phases of colon tumorigenesis could conceivably stop or retard tumor progression.
Footnotes
Acknowledgements
Supported by the Austrian National Bank Nr. 8296 and by Boehringer Ingelheim Austria, Vienna, Austria.