
Abstract
The main autocrine/paracrine role of the active metabolite of vitamin D3, 1α,25-dihydroxyvitamin D3 (1,25-D3), is inhibition of cell growth and induction of cell differentiation and/or apoptosis. Synthesis and degradation of the secosteroid occurs not only in the kidney but also in normal tissue or malignant extrarenal tissues such as the colon. Because 25-hydroxyvitamin D3 24-hydroxylase (CYP24A1) is considered to be the main enzyme determining the biological half-life of 1,25-D3, we have examined expression of the CYP24A1 mRNA (by real-time RT-PCR) and protein (by immunohistochemistry) in normal human colon mucosa, colorectal adenomas, and adenocarcinomas in 111 patients. Although 76% of the normal and benign colonic tissue was either completely devoid of or expressed very low levels of CYP24A1, in the majority of the adenocarcinomas (69%), the enzyme was present at high concentrations. A parallel increased expression of the proliferation marker Ki-67 in the same samples suggests that overexpression of CYP24A1 reduced local 1,25-D3 availability, decreasing its antiproliferative effect.
Keywords
C
A recent meta-analysis of 18 cohort and case-control studies showed that an elevation of serum 25-D3 concentration to levels ≥33 ng/ml led to a 50% lower incidence of CRC (Gorham et al. 2005). Cumulative epidemiological evidence suggests that there is a direct correlation between reduced CRC incidence and sunlight exposure, nutritional vitamin D intake, and high serum levels of 25-D3 (Giovannucci et al. 2006).
Vitamin D metabolism is a strictly regulated, multi-step process, beginning with the formation of previtamin D3 in the skin, mediated by ultraviolet radiation, or with absorption of vitamin D from dietary sources (Henry 1997; Sawada et al. 2000; Cheng et al. 2003). Vitamin D is hydroxylated by CYP27A1 to 25-hydroxyvitamin D3 in the liver. The last step of the activation is accomplished by the 25-hydroxyvitamin D3 1α hydroxylase (CYP27B1) in the kidney. The most active metabolite of vitamin D3, 1α,25-dyhydroxyvitamin D3 (1,25-D3, also known as calcitriol), has a crucial role, not only in the maintenance of calcium and phosphate homeostasis, but also in a wide range of other biological functions, such as the induction of cell differentiation, apoptosis and inhibition of cell growth, and angiogenesis (Ingraham et al. 2008). After 1,25-D3 binds to the vitamin D receptor (VDR) the 1,25-D3/VDR complex exerts transcriptional activation and/or repression of target genes (Kato 2000). The physiologically active 1,25-D3 is catabolized by the cytochrome P450 25-hydroxyvitamin D3 24-hydroxylase (encoded by the gene CYP24A1) (Sakaki et al. 2000).
Activation and degradation of 1,25-D3 are not confined to the kidney. The activation of 25-D3 by CYP27B1 can occur in a wide range of extrarenal tissues (including the colon) predicting a potential role of CYP27B1 as modulator of vitamin D functions (Cross et al. 1997; Zehnder et al. 2001). Similarly, CYP24A1 holds vitamin D effects under strict control by irreversible clearance of vitamin D metabolites in a tissue-specific manner. Studies on human colon cancer cell lines and tumor samples show tissue-specific expression and regulation of vitamin D3 hydroxylases in the colon (Kallay et al. 2005). In early-stage colorectal tumors, CYP27B1 levels are higher than in normal mucosa, suggesting an autocrine/ paracrine chemopreventive effect of the locally synthesized 1,25-D3. In poorly differentiated colon carcinomas, however, CYP27B1 is undetectable (Bises et al. 2004). Conversely, CYP24A1 mRNA expression is upregulated in highly malignant colon cells (Bareis et al. 2001). In addition to CRC, in several other malignancies also, CYP24A1 levels are increased when compared with matched normal tissues (Townsend et al. 2005; Anderson et al. 2006; Parise et al. 2006). Understanding the changes in the expression and effectiveness of the vitamin D system, beginning from the normal mucosa until development of highly malignant adenocarcinomas, would help to provide a rational basis for attempts to use vitamin D compounds for prevention or treatment of CRC.
In the present study, we examined expression of the vitamin D–catabolizing enzyme, CYP24A1, in human normal colon mucosa, colorectal adenomas, and adenocarcinomas, and analyzed its possible association with tumor cell proliferation.
Materials and Methods
Patients
Tissue specimens of cancerous lesions and histologically normal mucosa outside the tumor border were available from 48 patients (26 men and 22 women) who underwent surgery for primary colon adenocarcinoma at the 1st Department of Surgery, Semmelweis University, Budapest. The median age was 69 years (range 44–85). In addition, benign lesions of the colon: 3 hyperplastic polyps; 23 tubular, 4 tubulovillous, and 18 villous adenomas, were collected from 48 (30 male and 18 female) patients (median age 58 years, range 24–85). Normal colon tissue biopsies were obtained from 7 male and 8 female patients (median age 76 years, range 46–85) by endoscopy at the 1st Department of Medicine, Semmelweis University, Budapest. Tissue samples collected from patients were cut in two. Half of the specimen was frozen immediately in liquid nitrogen for RNA extraction, and the other half was fixed in formaldehyde for paraffin embedding.
Malignant tumors (all of the adenocarcinoma type) were graded according to the World Health Organization classification (Jass 1989). Of the 48 adenocarcinoma samples, 8 were of low grade, i.e., well-differentiated (G1), 27 were of intermediate grade, i.e., moderately differentiated (G2), and 13 were of high grade, i.e., poorly differentiated or undifferentiated (G3). The tumors were localized as follows: 11 in the rectum, 12 in the sigma, 3 in the descending, 7 in the transversal, and 6 in the ascending colon, and 7 in the cecum. Data on the localization of two tumor samples could not be obtained. Permission from the Ethics Commission of the Semmelweis University Medical School was granted prior to initiation of the study (ethical approval number 179/2007).
RNA Extraction and Quantitative Reverse Transcriptase Polymerase Chain Reaction (qRT-PCR)
Total RNA from each sample (70–80 mg tissue) was extracted by a spin-column-based method, using the RNEasy Mini Kit (Qiagen; Hilden, Germany), and was quantified by measuring the absorption at 260/280 nm by NanoDrop spectrophotometer (Nanodrop Technologies; Montchanin, DE). The integrity of the RNAs was determined by ethidium bromide staining on agarose gel. Two μg total RNA from each sample was reverse transcribed using Moloney murine leukemia virus reverse transcriptase, according to the manufacturer's instructions (Promega; Madison, WI), in a total volume of 20 μll.
VDR, CYP27B1, and CYP24A1 mRNA levels were quantified by the comparative ΔΔC
List of primer pairs
Immunohistochemistry
Immunohistochemistry for CYP24A1 was performed using a two-step indirect immunoperoxidase technique. Sections (5 μm) of paraffin-embedded tissue on silane-coated slides were incubated for 25 min at 60C, deparaffinized, and rehydrated. After two washes in phosphate-buffered saline (PBS, pH 7.2), the last washing containing 0.05% Tween-20, sections were boiled (three times for 5 min) in citrate buffer (pH 6.0) in a microwave oven for antigen retrieval. After washes in PBS, sections were permeabilized in 0.2% Tween-20 for 10 min. Endogenous peroxidase was inactivated by incubation in 3% H2O2 in methanol for 10 min. Sections chosen for CYP24A1 and VDR double staining underwent an additional inactivation of the endogenous alkaline phosphatase with 0.2 N HCl at room temperature for 8 min after the slides were washed with PBS-Tween, 0.05%. Normal kidney sections were used as positive control.
Single immunostaining (for CYP24A1) was performed using the Ultravision LP Detecting System (LabVision Corporation; Fremont, CA) according to the manufacturer's instructions. Briefly, after washing with PBS/ 0.05% Tween-20 and blocking with the Ultra Block solution, sections were incubated with the polyclonal rabbit anti-CYP24A1 antibody (dilution 1:500) overnight at 4C. The anti-CYP24A1 antibody was kindly provided by Dr. Pamela Hershberger (University of Pittsburgh Cancer Institute). After washing in PBS/ 0.05% Tween-20, the enhancer solution was applied for 20 min, followed by horseradish peroxidase–labeled rabbit polymer for 30 min. CYP24A1 was visualized by incubation with diaminobenzidine tetrahydrochloride (DAB) chromogen (DakoCytomation; Carpinteria, CA). The sections were then washed with deionized water and counterstained with hematoxylin (DakoCytomation) for 2 min, dehydrated, and mounted in Eukitt (Kindler; Freiburg, Germany). As negative control, isotype-matched IgG was used instead of the primary antibody. Expression of CYP24A1 was evaluated semiquantitatively. Ten randomly selected areas of each slide were manually analyzed using the high-power field objective (x40), with 500 cells counted per field. Staining intensities were classified as no staining (neg), weak positive (+), moderate positive (+ +), and strong positive (+ + +).
For CYP24A1 and VDR double immunohistochemical staining, the sections were incubated with primary anti-CYP24A1 antibody for 60 min at room temperature and stained with the Ultravision system as described above. After visualization of CYP24A1, rabbit serum was used for blocking (30 min), and then the monoclonal rat anti-VDR antibody (Chemicon; Temecula, CA), dilution 1:200, was applied overnight at 4C. Biotinylated polyclonal rabbit anti-rat immunoglobulin (DakoCytomation) was added as secondary antibody for 1 hr, prior to washing in PBS/0.05% Tween-20 and visualization of VDR with streptavidin-alkaline phosphatase conjugate (30 min) and permanent red chromogene (3 min, both DakoCytomation). Finally, the sections were counterstained with hematoxylin and mounted in Glycergel Mounting Medium (both DakoCytomation) without prior dehydration.
Ki-67 staining was performed with the Ventana ES automatic immunostainer device. Briefly, after deparaffination, antibody retrieval was carried out in Tris (hydroxymethyl)-aminomethan-ethylenediaminetetra-acetic acid (TRIS-EDTA) solution (Target Retrieval Solution; DakoCytomation) in a microwave oven (30 min). Monoclonal mouse primary Ki-67 antibody (DakoCytomation) was used in a 1:120 dilution for 32 min at 42C after peroxidase and serum blocking. The expression was visualized with DAB chromogen substrate (DakoCytomation). Positive controls recommended by the manufacturer (DakoCytomation) were used to confirm correct immunohistochemical staining for Ki-67. As negative control, isotype-matched IgG was used instead of the primary antibody. Scoring for semiquantitative analysis: ten randomly selected areas of each slide were manually analyzed using a high-power field objective (x40), with 1000 cells counted per field. Immunoreaction was evaluated as the percentage of cells staining positively. The following values were given for semiquantitative evaluation: 0 (0–5% positivity), 1 (6–20% positivity), 2 (21–40% positivity), 3 (41–60% positivity), 4 (61–80% positivity), and 5 (81–100% positivity). Immunoreactions were scored as positive where nuclear staining was seen.
Statistics
The unpaired t-test was used to analyze the association between the expression levels estimated by real-time RT-PCR and the type of the sample (normal, adenoma, adenocarcinoma). The relations between characteristics variables were examined by χ2 test, or Fisher's exact test, as appropriate. Hockberg's procedure was used in cases of multiple comparisons. Significance was defined as p<0.05. To measure the correlation between CYP24A1 and Ki-67 protein expression, we have calculated Spearman's rank correlation coefficient. The data were analyzed using the SPSS statistics package (SPSS; Chicago, IL), version 15.0 for Windows.
Results
Detection of CYP24A1 mRNA by Real-time RT-PCR
We grouped the tissue samples from our patient cohort into three categories: normal mucosa, benign lesions (polyps and adenomas), and adenocarcinomas. We measured CYP24A1 mRNA expression levels using real-time qRT-PCR. In parallel we have determined the mRNA expression level of the 1,25-D3–synthesizing enzyme (CYP27B1) and that of the VDR, as well (Table 2). CYP24A1 levels were significantly higher both in the benign lesions and in adenocarcinomas, compared with the normal mucosa. In this patient cohort, CYP27B1 expression did not change significantly, although there was a trend toward lower expression in adenocarcinomas. VDR expression, however, was significantly decreased in polyps and adenomas and was even more decreased in adenocarcinomas.
Expression of CYP24A1 Protein in Human Colon Tissue
Immunohistochemistry showed marked differences in the level of CYP24A1 expression with increasing malignity (Figure 1). None to low expression of CYP24A1 protein was apparent in normal colonic mucosa and in the adjacent normal colonic epithelial part of sections with pathological features (Figure 1E), as well as in the majority of adenomas (Figure 1A). In contrast, expression of CYP24A1 was significantly higher in the adenocarcinomas (Figures 1C, 1E, and 1H), highlighted also in Table 3. For statistical analysis, polyps and adenomas were merged into a single category to avoid inaccuracies in statistical analysis due to low case numbers in the polyp group. CYP24A1 was mainly present in the epithelial cells, and stromal expression was very low, if any. Expression of CYP24A1 was confined to the cytoplasm in all studied materials (Figures 1A, 1C, and 1H, insets). The punctated pattern of the staining is consistent with the mitochondrial localization of the enzyme.
Evaluation of relative VDR, CYP24A1, and CYP27B1 mRNA expression levels by qRT-PCR in normal colonic mucosa, benign lesions, and adenocarcinomas
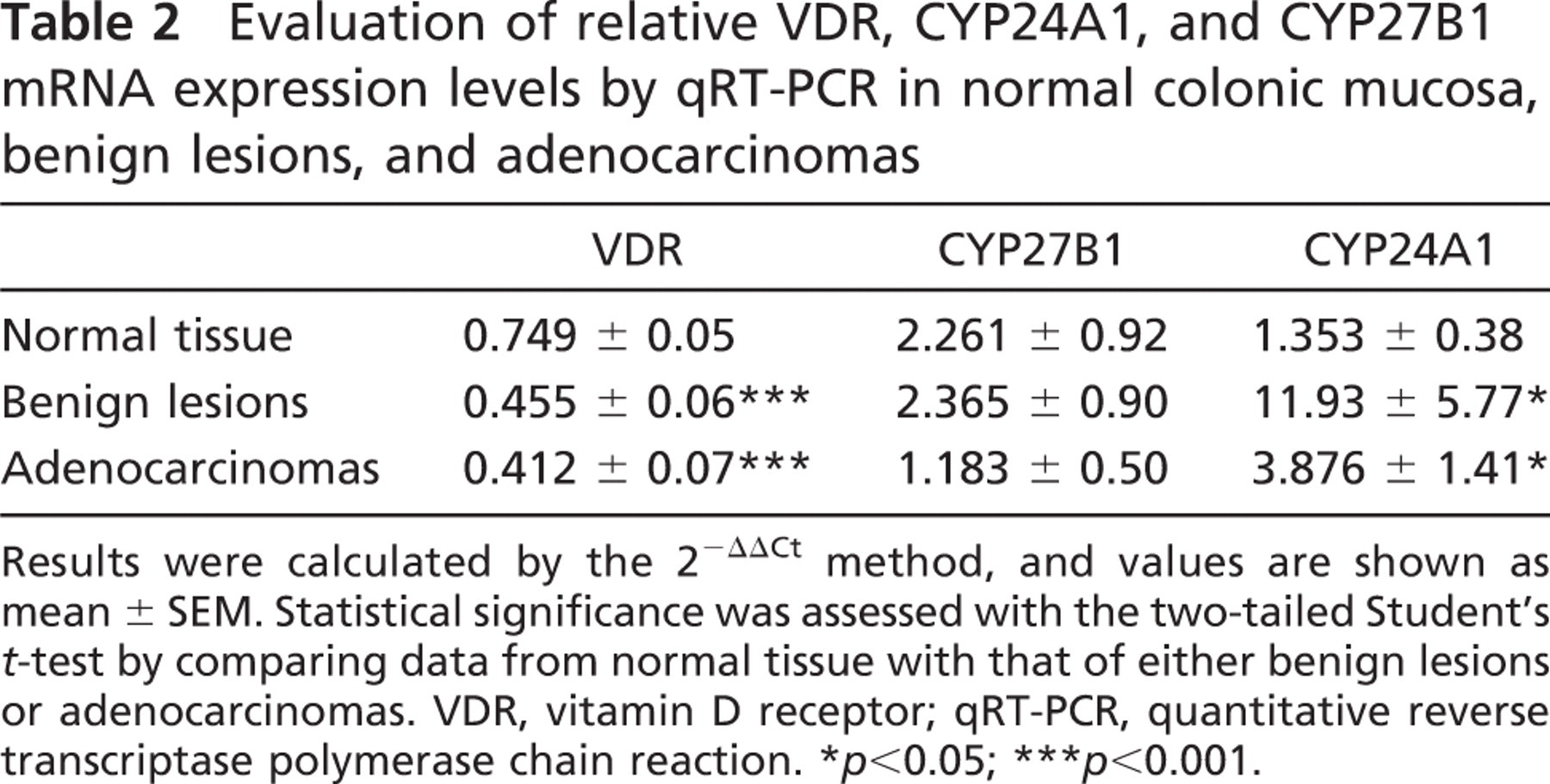
Results were calculated by the 2−ΔΔCt method, and values are shown as mean ± SEM. Statistical significance was assessed with the two-tailed Student's t-test by comparing data from normal tissue with that of either benign lesions or adenocarcinomas. VDR, vitamin D receptor; qRT-PCR, quantitative reverse transcriptase polymerase chain reaction. ∗ p<0.05; ∗∗∗ p<0.001.
We examined possible associations between CYP24A1 expression and clinical characteristics of the patients. No statistical correlation was found between CYP24A1 expression and patients’ gender or age at diagnosis in our patient cohort.
We also examined the relationship between the expression of CYP24A1 and the histological parameters of the tumors (tumor/lymph node/metastasis stage, grade, level of necrosis, tumor vascularization, and perifocal inflammation), but no correlation was found between these parameters and CYP24A1 expression (Table 4.).
To test our hypothesis that CYP24A1 overexpression would reduce local 1,25-D3 availability, decreasing its antiproliferative effect, we examined the association between the expression of CYP24A1 and that of Ki-67, a widely accepted proliferation marker (Figures 1B and 1D). Indeed, we found a strong correlation between CYP24A1 and Ki-67 expression; the overall correlation coefficient was r s = 0.376 (p<0.001) (Table 5). Intriguingly, the correlation was also strong when we considered only the normal and benign samples (r s = 0.341; p<0.001) but was lost when we considered only the carcinomas.
We also assessed expression of the VDR on serial sections in our patient cohort by double staining with CYP24A1. We have not seen any correlation between CYP24A1 and VDR expression (Figures 1H and 1I). In addition to sections highly positive both for VDR and CYP24A1, there were tissue samples expressing only one of the examined proteins. Figure 1H illustrates a section with high CYP24A1 expression. The inset reveals that strong CYP24A1 positivity can be found both in cells with high VDR expression, as well as in cells lacking VDR. Figure 1I is an example of a sample with high VDR levels that is almost completely devoid of CYP24A1.
Discussion
Accumulating epidemiological data suggest a protective role for vitamin D3 against various malignancies; therefore, extraskeletal antimitogenic effects of 1,25-D3 have been studied intensively (Deeb et al. 2007; Fleet 2008). The most-studied effect of 1,25-D3, in addition to calcium regulation and phosphate homeostasis, is growth inhibition of proliferating cells.
Our study is the first to compare expression of the vitamin D–catabolizing enzyme CYP24A1 in human normal and pathological colon tissues, on both the mRNA and the protein level, and to correlate it with proliferation.
CYP24A1 is one of the main target genes of 1,25-D3 bound to VDR. Constitutive (basic) levels are very low, except in the kidney, and induction occurs quickly, in some tissues achieving a several hundred-fold increase, exceeding the expression of most other vitamin D–responsive genes. Local CYP24A1 is induced by 1,25-D3 synthesized from its precursor, 25-D3, in cells expressing CYP27B1. In cells with a balanced vitamin D system, there may be a period of several hours while 1,25-D3 levels are above baseline before the newly transcribed CYP24A1 reduces local 1,25-D3 levels in a negative-feedback manner (Schuster et al. 2006).
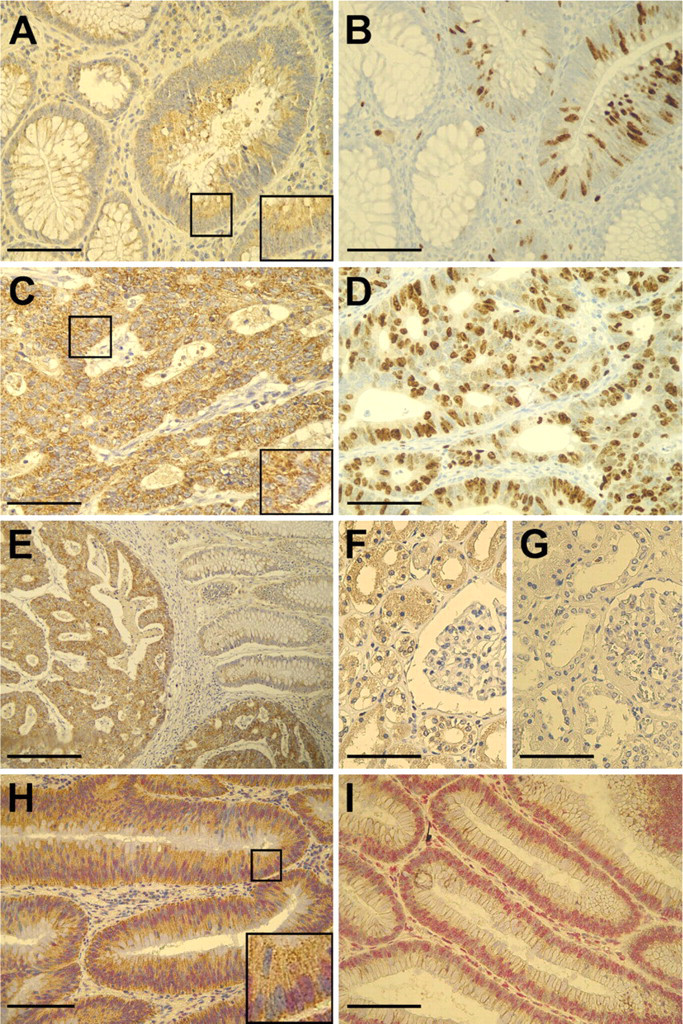
Immunhistochemical staining of CYP24A1
Evaluation of CYP24A1 immunoreactivity in normal colonic mucosa, benign lesions, and adenocarcinomas
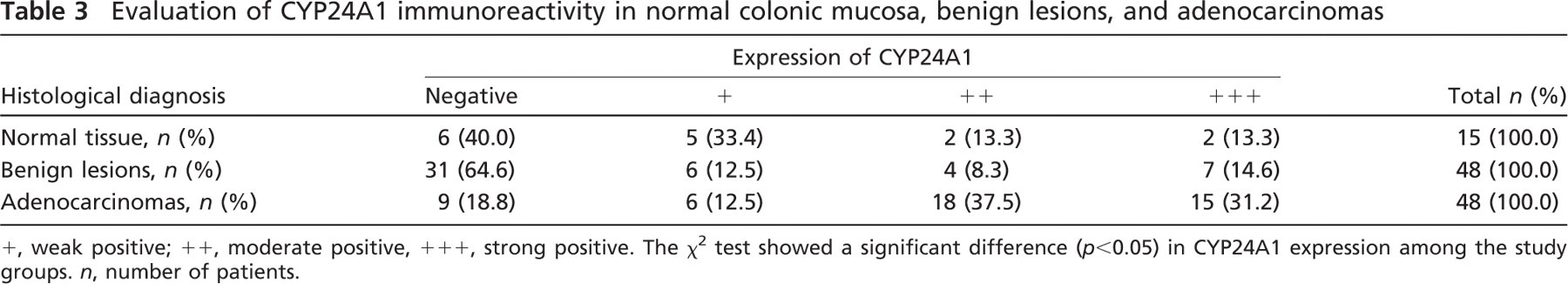
+, weak positive; + +, moderate positive, + + +, strong positive. The χ2 test showed a significant difference (p<0.05) in CYP24A1 expression among the study groups. n, number of patients.
In several cancers, an unbalanced overexpression of CYP24A1 was reported. The upregulation of CYP24A1 mRNA was observed not only in colon (Bareis et al. 2001; Cross and Kallay 2005; Anderson et al. 2006) but also in cervical (Friedrich et al. 2003), ovarian (Friedrich et al. 2003; Anderson et al. 2006), basal cell (Mitschele et al. 2004), cutaneous squamous cell (Reichrath et al. 2004), esophageal (Mimori et al. 2004), and lung (Parise et al. 2006) carcinomas, suggesting involvement of CYP24A1 in tumorigenesis. Whether this increase in mRNA translates into higher amounts of the degrading enzyme and ultimately in more degradation of the active compound needed to be addressed. We indeed have observed discrepancies between mRNA and protein expression levels (Tables 2 and 3). This might be a consequence of different factors, e.g., imbalance between CYP24A1 mRNA and protein stability and half-life, or differences in the method of analysis and data presentation.
Our study showed that in normal colon mucosa, colon polyps, and adenomas CYP24A1, protein expression was very low. In 37 out of 63 tissue samples (59%), CYP24A1 was undetectable, whereas 11 samples (out of 63) were moderately positive. In most of the adenocarcinomas (69%), the major 1,25-D3–degrading enzyme was overexpressed (Figure 1; Table 3). This indicates that 24-hydroxylase activity will blunt the increment of endogenously synthesized 1,25-D3 mainly in adenocarcinomas, but not during early tumorigenesis, underlying the importance of vitamin D3 mainly as a chemopreventive agent.
Correlation between expression of CYP24A1 and tumor parameters

p<0.05 is considered significant.
To determine whether the upregulation of CYP24A1 expression was a result of possible higher local 1,25-D3 concentrations and/or elevated VDR levels, we measured expression of CYP27B1 and VDR in our patient cohort. mRNA data showed that whereas CYP27B1 levels did not change significantly, although they had a tendency to decrease in adenocarcinomas, VDR levels were significantly lower in both benign and malignant tumors. We have also not detected any correlation between VDR and CYP24A1 protein levels. Therefore, it is unlikely that the high CYP24A1 levels in the malignant tumors were a result of the normal, physiological transcriptional induction by VDR-bound 1,25-D3.
One of the possible causes of elevated CYP24A1 expression might be gene amplification. Several studies have already identified gain of 20q13 (the gene locus of human CYP24A1) in a variety of malignancies (Zitzelsberger et al. 2001; Fridlyand et al. 2006; Lin et al. 2006), including colorectal carcinoma (Diep et al. 2006). Chromosome 20q gain was also suggested to be involved in the progression of colorectal adenoma to carcinoma, and was often associated with liver metastasis (Korn et al. 1999). Recently, results of two independent array-based comparative genomic hybridization studies confirmed, in a large series of sporadic colorectal carcinomas, amplification of a region on the long arm of chromosome 20, including the CYP24A1 gene (Nakao et al. 2004; Lassmann et al. 2007). However, unequivocal proof that amplification of the 20q13 locus results in high CYP24A1 expression is still missing. Whether other cell-type–selective regulators of CYP24A1 gene expression, such as parathyroid hormone, calcitonin, phorbol ester, γ-interferon, etc. (Omdahl 2005), were involved in the overexpression of CYP24A1 in colorectal tumors was not examined.
A recent study showed nuclear expression of both CYP27A1 protein and CPY24A1 protein mainly in normal colonic epithelium (Matusiak and Benya 2007). The authors suggested that translocation of these proteins from the nucleus to the cytoplasm is a function of malignant progression. However, both enzymes belong to the cytochrome P450 family and are mixed-function oxidoreductases. CYP24A1 (similarly to CYP27B1) is located in the inner mitochondrial membrane, where it is part of a mini–electron transport chain consisting of NADPH-ferredoxin reductase, ferredoxin, and the terminal cytochrome CYP24A1 (Gray et al. 1990; Omdahl 2005). Therefore, it is very unlikely that these enzymes would be functional outside the mitochondrion. For proper functioning, CYP24A1 needs a 150-aa N-terminal sequence containing the mitochondrial targeting domain (Ou et al. 1994). A recent study revealed that a splice variant of CYP24A1 that lacks this mitochondrial target sequence failed to produce any 24-hydroxylase activity (Ren et al. 2005), although it was still able to bind both 25-D3 and 1,25-D3. In our study, expression of CYP24A1 was confined to the cytoplasm of cells, and nuclear localization was detected neither in the colon specimens nor in the kidney (used as positive control) (Figures 1A, 1C, 1E, 1F, 1H, and 1I). The discrepancies with the data of Matusiak and Benya (2007) might be due to the quality of antibodies used. The primary antibody used for immunohistochemistry in our set of samples has been proven to be specific for CYP24A1 in immunoblot analysis, recognizing one single band at 59 kDa. Subcellular fractionation of this antibody demonstrated that the immunoreactive species localizes to the mitochondrial/membrane fraction (Parise et al. 2006). Thus, the staining that we observe in our tissues is consistent with the results obtained in subcellular fractionation studies.
The association of CYP24A1 and Ki-67 expression
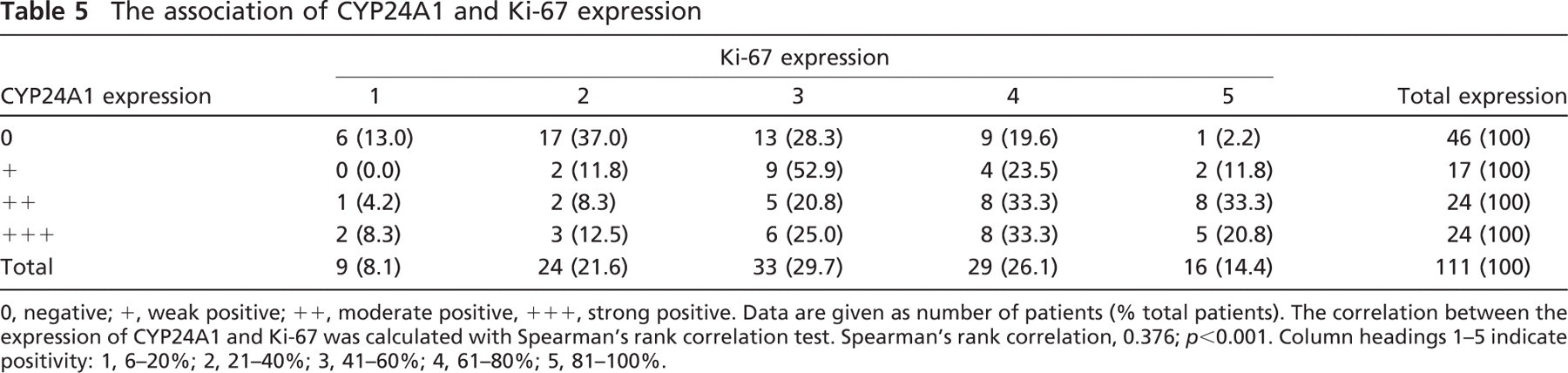
0, negative; +, weak positive; + +, moderate positive, + + +, strong positive. Data are given as number of patients (% total patients). The correlation between the expression of CYP24A1 and Ki-67 was calculated with Spearman's rank correlation test. Spearman's rank correlation, 0.376; p<0.001. Column headings 1–5 indicate positivity: 1, 6–20%; 2, 21–40%; 3, 41–60%; 4, 61–80%; 5, 81–100%.
High proliferation is a well-known attribute of tumor cells. The causes are multifactorial. The growth-inhibiting role of vitamin D has been studied extensively (for review, see, e.g., Deeb et al. 2007; Fleet 2008). Our study is the first to examine the correlation between proliferation and CYP24A1 mRNA and protein expression during colorectal tumorigenesis. We have shown previously (Cross and Kallay 2005) that in colon adenocarcinoma cells expressing the 1,25-D3–synthesizing enzyme CYP27B1, the proliferation marker Ki-67 was barely present, suggesting a functioning, antiproliferative vitamin D system. In the present study, we found a significant correlation between CYP24A1 and Ki-67 expression, revealing that tissues with high CYP24A1 levels have an increased proliferation rate. Whether there is causality in this observation needs to be further investigated. Parise et al. (2009) have shown that in lung tumor xenografts, CYP24A1 overexpression resulted in elimination of local 1,25-D3. Therefore, it is likely that CYP24A1 overexpression would cause reduced local calcitroic levels, decreasing the antiproliferative, antimitotic activity of the vitamin D hormone and thus contributing (at least to some extent) to hyperproliferation in colon tumors also. The fact that the correlation was more obvious when restricted to the normal and benign lesions suggests a greater importance for the vitamin D system in prevention and early tumourigenesis. Consequently, inhibition of the vitamin D–catabolizing enzyme would be a rational option when using vitamin D3 for tumor prevention and treatment, because it might potentiate the antitumor effects of 1,25-D3 by prolonging its half-life. Several studies support the idea of using inhibitors of CYP24A1 in anti-cancer treatment. Lechner et al. (2007) have demonstrated that a 24-phenylsulfone analog of 1,25-D3, KRC-24SO2 Ph-1, rapidly and potently inhibited 24-hydroxylase activity, and not only increased the half-life of 1,25-D3, but also rescued its antiproliferative effect in human colon cancer cells. Ly et al. (1999) showed that liarazole, a selective CYP24A1 blocker, administered together with 1,25-D3, synergically inhibited the growth of human prostate cancer cells. In addition, the P450 inhibitor ketoconazole also reinforced the antiproliferative effect of calcitriol in vitro (Peehl et al. 2002) and in vivo (Kang et al. 1997). The chemotherapeutic agents daunorubicin hydrochloride, etoposide, and vincristin sulfate induced the degradation of CYP24A1 mRNA, and thus enhanced 1,25-D3 levels in kidney cells (Tan et al. 2007). Although the effect of two selective CYP24A1 inhibitors on human keratinocytes was intensively investigated in a preclinical study (Schuster et al. 2001), no clinical studies have as yet attempted to administer inhibitors of CYP24A1 in human malignancies. Our results further support the importance of selective CYP24A1 inhibition when employing vitamin D in the treatment of CRC.
In summary, this is the first study to correlate CYP24A1 protein levels with proliferation, suggesting that CYP24A1 overexpression counteracts the antiproliferative effect of 1,25-D3 during tumor progression. The autocrine/ paracrine growth regulation of colonocytes by the active vitamin D seems to be functional only in the early stages of tumorigenesis, and it becomes impaired during tumor progression, owing to CYP24A1 overexpression. 1,25-D3 potentiates the antitumor effect of several chemotherapeutic agents, such as taxanes and platinum analogs (Hershberger et al. 2001; Deeb et al. 2007); therefore, modulating expression of the vitamin D–catabolizing enzyme in colorectal tumorigenesis might offer new antitumor strategies.
Footnotes
Acknowledgements
This work was supported by a grant from the Austrian Academic Exchange Service (A11-2004; to HCH, GS, EK, PL), from Aktion Österreich-Ungarn (73öu3; to HCH, GS, EK), the TEVA-Biogal Research Award for Physicians (to HCH, JPK, GS), grants NKFP-1A/007/2004 and NKFP-1A/ 002/2004 from the National Research and Technological Office of Hungary (to JPK), research grant ETT 022/2006 from the Hungarian Ministry of Health (to JPK), and a grant from the European Union (MRTN-CT-2005-019496; to TN).