
Abstract
White adipose tissue (WAT) is innervated by the sympathetic nervous system. A role for WAT sympathetic noradrenergic nerves in lipid mobilization has been suggested. To gain insight into the involvement of nerve activity in the delipidation process, WAT nerves were investigated in rat retroperitoneal and epididymal depots after prolonged fasting. A significant increase in tyrosine hydroxylase (TH) content was found in epididymal and, especially, retroperitoneal WAT by Western blotting. Accordingly, an increased immunoreactivity for TH was detected by immunohistochemistry in epididymal and, especially, retroperitoneal vascular and parenchymal noradrenergic nerves. Neuropeptide Y (NPY)-containing nerves were found around arteries and in the parenchyma. Double-staining experiments and confocal microscopy showed that most perivascular and some parenchymal noradrenergic nerves also contained NPY. Detection of protein gene product (PGP) 9.5, a general marker of peripheral nerves, by Western blotting and PGP 9.5-TH by double-staining experiments showed significantly increased noradrenergic nerve density in fasted retroperitoneal, but not epididymal depots, suggesting that formation of new nerves takes place in retroperitoneal WAT in fasting conditions. On the whole, these data confirm the important role of sympathetic noradrenergic nerves in WAT lipid mobilization during fasting but also raise questions about the physiological role of regional-dependent nerve adjustments and their functional significance in relation to white adipocyte secretory products.
M
Some experimental evidence suggests that lipid mobilization from WAT is under the control of the sympathetic nervous system. Indeed, electrical stimulation of adipose tissue nerves increases production of free fatty acids (Correll 1963). Unilateral denervation of bilateral fat depots results in a mass asymmetry suggestive of diminished fat mobilization from the denervated tissue (Cantu and Goodman 1967). Cold exposure (Garofalo et al. 1996), as well as fasting (Migliorini et al. 1997), increases noradrenaline turnover in WAT and, finally, the naturally occurring decrease in body fat observed in Siberian hamsters exposed to a short “winter-like” photoperiod is accompanied by an increased noradrenaline turnover in their visceral fat (Youngstrom and Bartness 1995). Thus, in conditions such as prolonged fasting, severe exercise, stress and cold exposure, direct and selective activation of the sympathetic nerves supplying WAT—rather than adrenal medullary catecholamines or catabolic hormones, as proposed in origin (Cahill 1970)—is believed to be the main mechanism triggering the increased rate of lipolysis in white adipocytes.
In small rodents, WAT is located in several subcutaneous and visceral depots (Cinti 1999). It is provided with sympathetic nerves distributed as perivascular and sparse parenchymal axons (Slavin and Ballard 1978; Bartness and Bamshad 1998; Cinti 1999) and with a smaller amount of peptidergic nerves containing neuropeptide Y (NPY), substance P (SP), and calcitonin gene-related peptide (CGRP) (Cui and Himms-Hagen 1992; Giordano et al. 1996,1998), the latter two neuropeptides usually being found in sensory nerves.
Although the WAT nerve supply and its distribution in normal conditions and in different species have been the subject of detailed morphological investigations (Bartness et al. 2001), systematic studies of nerve distribution in fasting animals have still not been performed. In particular, we reasoned that if fasting-dependent lipid mobilization is really due to WAT sympathetic activation, plastic nervous changes should occur in fasted WAT to allow the nerve supply to meet lipid mobilization requirements. To address these questions, rats were fasted for different periods of time (from 3 to 7 days) to obtain four groups of animals with body weight losses of ∼15%, 20%, 25%, and 30%. Because WAT is a multi-depot organ, with possible locally relevant metabolic and secretory functions (Rayner 2001), we examined the two largest visceral WAT depots in small rodents, the retroperitoneal and the epididymal depot. Their nerve supply was studied using biochemical (Western blotting) and morphological methods (immunohistochemistry on cryosections and confocal microscopy). Protein gene product (PGP) 9.5, which is considered a highly sensitive marker of neuronal elements including peripheral axonal projections (Thompson et al. 1983; Wilson et al. 1988), was employed as a pan-peripheral nerve marker, and tyrosine hydroxylase (TH), the rate-limiting enzyme in the biosynthesis of catecholamines, was evaluated to determine the amount of noradrenergic nerves and visualize their distribution. The presence and distribution of NPY-containing nerves were also analyzed.
Materials and Methods
Animals and Tissues
Male Sprague-Dawley rats 9 weeks of age (180–220 g) were obtained from Morini Laboratories (S. Polo d'Enza, Italy). They were caged singly and kept at 22 ± 2C with a 12:12-hr light-dark cycle. Control animals had free access to water and standard chow (65% carbohydrates, 11% fat, 24% protein, w/w), whereas fasted animals were given only water. All animals were weighed daily at 9:00
Fasted animals were sacrificed when they had lost 15 ± 1% (after 3 days of fasting), 20 ± 1% (after 4–5 days), 25 ± 1% (after 5–6 days), or 30 ± 1% (after 6–7 days) of body weight. For immunohistochemistry, rats were anesthetized with 100 mg/kg ketamine (Ketavet; Farm. Gellini, Aprilia, Italy) in combination with 19 mg/kg xylazine (Rompum; Bayer AG, Leverkusen, Germany) and transcardially perfused with 4% formaldehyde in 0.1 M phosphate buffer (PB), pH 7.4. WAT depots were carefully dissected, measured (weight and volume), and postfixed by overnight immersion in the same fixative at 4C. After a brief wash in PB, tissues were cryo-protected in a solution of 30% sucrose in PB for 24 hr at 4C. A glass beaker containing 2-methylbutane (isopentane) was then cooled in liquid nitrogen and the specimens were embedded in OCT medium, frozen, and stored at −80C. For Western blot analysis, rats were decapitated and WAT depots carefully dissected, rapidly frozen in liquid nitrogen, and stored at −80C.
Antibodies
The following antibodies were used for both immunohistochemistry and Western blotting: rabbit polyclonal serum against PGP 9.5 (Ultraclone; Isle of Wight, UK), sheep polyclonal serum against TH (Chemicon; Temecula, CA), and rabbit polyclonal serum against neuropeptide Y (NPY) (Sigma; St Louis, MO).
Western Blotting
Adipose tissue samples were taken from three rats per group. For PGP 9.5 and TH analysis, 100 μg and 80 μg, respectively, of total protein extracts were boiled, resolved by 12% SDS-PAGE under reducing conditions, and transferred to nitrocellulose filter paper. Incubation with the primary antibodies (PGP 9.5: 1:1700 v/v dilution; TH: 1:1000 v/v dilution) was performed for 2 hr at room temperature in a solution containing 20 mM Tris/HCl, pH 7.4 and 150 mM NaCl (TBS) with 5% milk. After multiple washes in TBS with 0.1% Tween, membranes were incubated with horseradish peroxidase-conjugated IgG secondary antibodies (1:40,000) for 1 hr at room temperature. The signal was detected on Biomax film (Kodak; Cinisello Balsamo, Milan, Italy) with Super Signal chemiluminescence Kits (Pierce Chemical Co.; Rockland, IL; femto and pico sensitivity for TH and PGP 9.5, respectively). Densitometric measurements were performed with Phoretix 1D, version 3.0 (Phoretix International Ltd.; King's Manor, Newcastle, UK).
Immunohistochemistry
Four-μm-thick cryosections obtained with a Leica CM1900 cryostat (Leica Microsystems; Vienna, Austria) were collected and air dried overnight at room temperature. Immunoreactivity was visualized using the avidin-biotin-peroxidase (ABC) method. After two 15-min washes in 0.05 M phosphate-buffered saline (PBS), pH 7.4, sections were incubated with 0.3% H2O2 in methanol for 30 min at room temperature to block endogenous peroxidase, washed twice in PBS (2 × 15 min), and incubated in 1:75 v/v normal goat serum (PGP 9.5 and NPY schedule; Vector Laboratories, Burlingame, CA) or in 1:75 v/v normal rabbit serum (TH schedule; Vector Laboratories) in PBS for 20 min at room temperature to block nonspecific sites. Sections were incubated overnight at 4C with the primary antibody at the following dilutions (v/v): PGP 9.5, 1:4000; TH, 1:500; NPY, 1:10,000; washed twice with PBS (2 × 15 min); incubated in 1:200 v/v biotinylated IgG anti-rabbit goat serum (PGP 9.5 and NPY schedule; Vector Laboratories) or 1:200 v/v biotinylated IgG anti-sheep rabbit serum (TH schedule; Vector Laboratories) in PBS for 30 min at room temperature; washed twice with PBS (2 × 15 min); incubated in ABC reagent (Vector Laboratories) in PBS for 1 hr at room temperature; washed twice with PBS (2 × 15 min); incubated in 0.02% H2O2 and 0.075% diaminobenzidine (Sigma) in 0.05 M Tris buffer (pH 7.6) and kept for 5 min in a dark room; rinsed in distilled water; and counterstained with hematoxylin. Sections were then dehydrated and mounted in Entellan. The ability of the antibodies to specifically detect the antigens was evaluated in sections (prepared as described above) of tissues known to contain the antigens (heart and intestine for PGP 9.5 and central nervous system and heart for TH- and NPY-containing nerves). Negative controls were obtained in each instance by omitting the primary antibody and using preimmune instead of primary antiserum. The images were stored as TIFF files. Brightness and contrast of the final images were adjusted using the Photoshop 6 software (Adobe Systems; Mountain View, CA).
Immunofluorescence and Confocal Microscopy
Immunofluorescence was performed on cryosections obtained according to the procedure described above. After two 15-min washes in PB, sections were incubated in 1:75 v/v normal donkey serum (Jackson ImmunoResearch; West Grove, PA) in PB for 20 min at room temperature to block nonspecific sites. Sections were incubated overnight at 4C with the mixture containing the primary antibodies (PGP 9.5 and TH at dilutions of 1:4000 and 1:300; TH and NPY at 1:300 and 1:8000, respectively); washed twice with PB (2 × 15 min); and incubated in 1:100 v/v FITC donkey anti-rabbit (Jackson) and TRITC donkey anti-sheep (Jackson) antibodies in PB for 30 min at room temperature. Sections were subsequently washed twice with PB, air dried, and coverslipped using Vectashield mounting medium (Vector Laboratories). Fluorescence was detected with a BioRad (Hercules, CA) microradiance confocal laser-scanning microscope equipped with an argon and He/Ne mixed gas laser. FITC and TRITC were excited with the 488- and 543-nm lines, respectively, imaged separately (using 515/30 and 570 nm filters, respectively). Sections were viewed in an Eclipse E600 Nikon microscope (Nikon; Sesto Fiorentino, Florence, Italy) with a X60 plan-apochromat objective and 1.4 numerical aperture. Images (512 × 512 pixels) were obtained sequentially from two channels using a confocal pinhole of 2–3 o.u. and stored as TIFF files. Brightness and contrast of the final images were adjusted using the Photoshop 6 software (Adobe Systems).
Results
Retroperitoneal Depot
The retroperitoneal lipid store was massively and rapidly mobilized during fasting; indeed, in the rats that had lost 15% of body weight it shrank in weight by ∼85% (Figure 1A) and in volume by ∼50% (Figure 1B). At light microscopy, the vast majority of retroperitoneal adipocytes in the rats that had lost 15% and 25% of body weight exhibited signs of delipidation (reduced cell diameter, thicker cytoplasmic rim, and small lipid droplets). As previously reported (Napolitano and Gagne 1963; Cinti 1999), the delipidation process was not uniform, as clusters of shrinking adipocytes were found among almost unchanged cells. Finally, in animals with a weight loss of 30% the vast majority of adipocytes contained no lipids and the remaining cells still exhibited small lipid droplets.
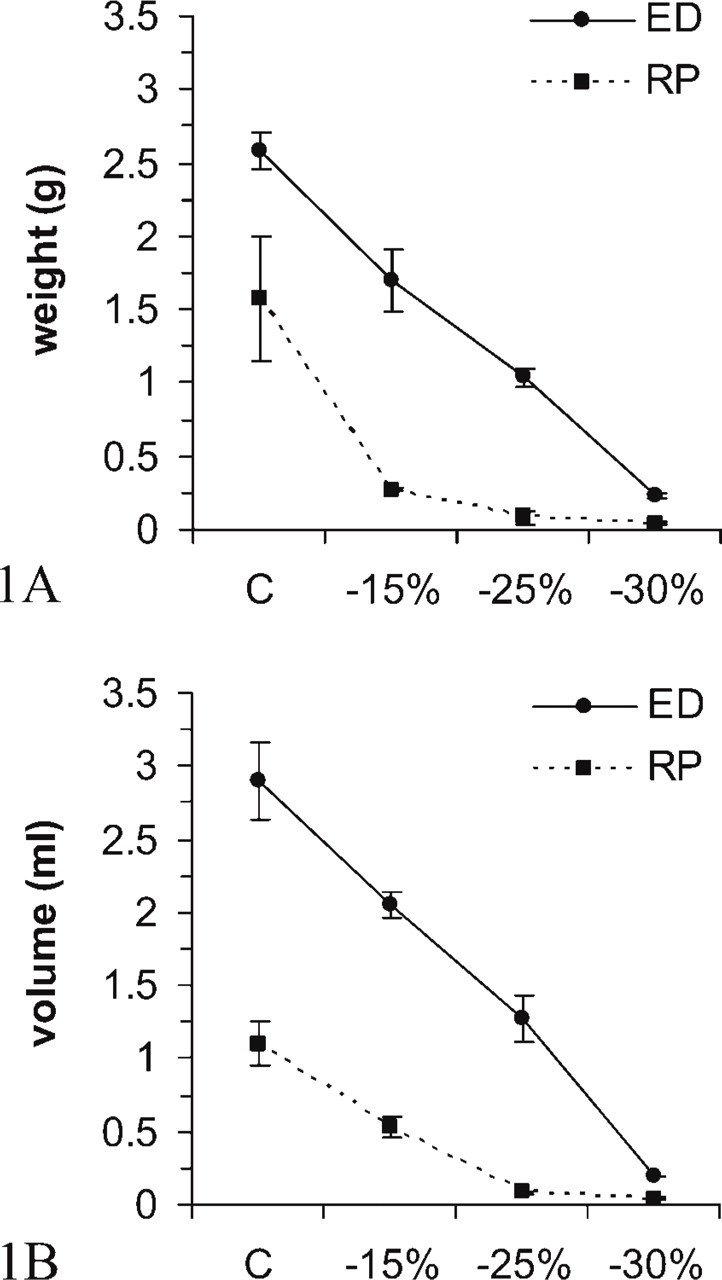
Weight (
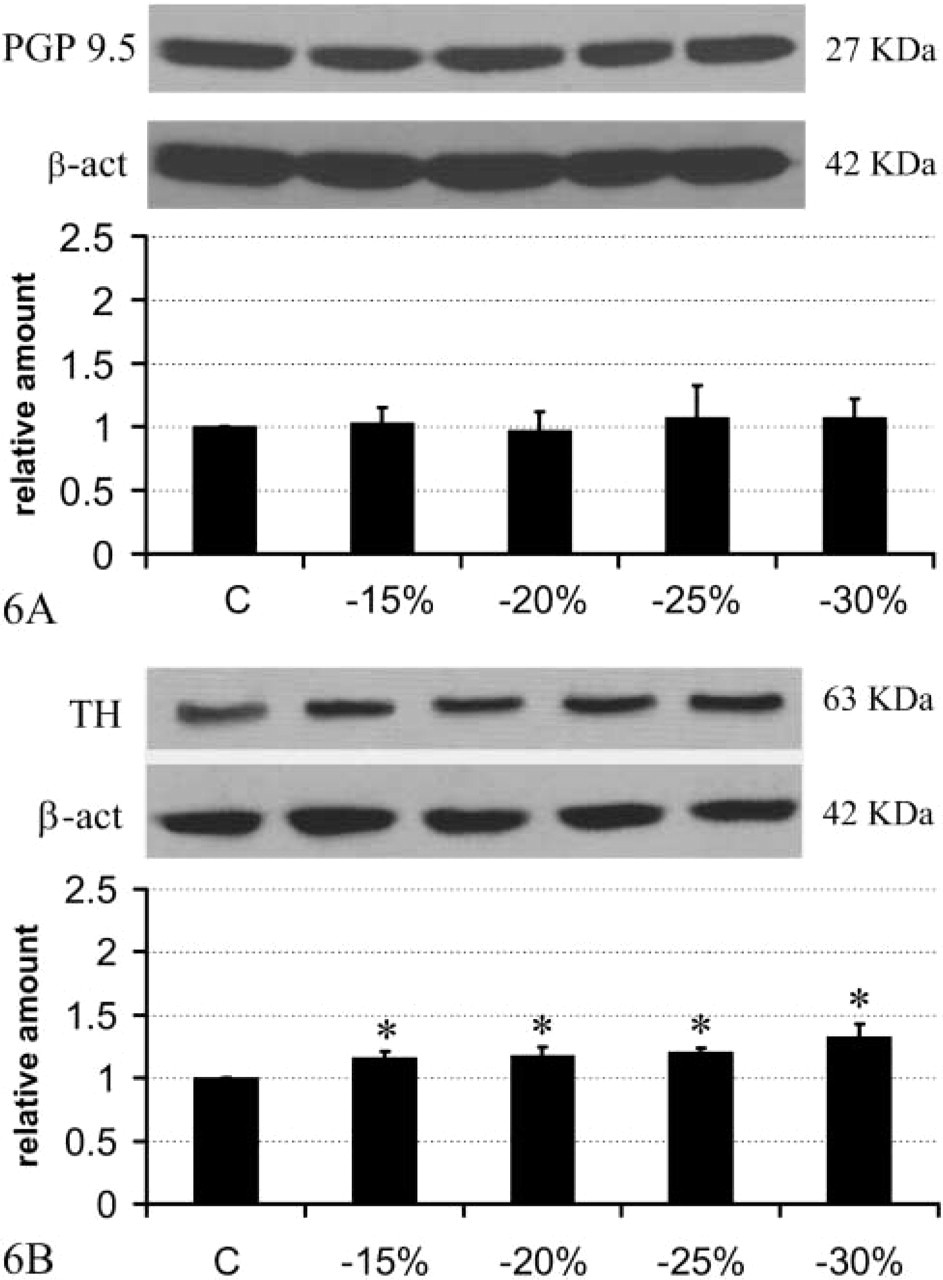
Brown adipose tissue (BAT) is among the most innervated tissues in the mammalian body, sharing the same degree of innervation with the heart (Barnard et al. 1980). Using PGP 9.5 as a general marker of peripheral nerves, retroperitoneal WAT was shown to be ∼3.5 times less innervated than interscapular BAT (Figure 2), in line with morphological and functional data demonstrating a lower degree of innervation in WAT than BAT (Bartness and Bamshad 1998; Cinti 1999). Previous data obtained with fluorescence histochemistry (Slavin and Ballard 1978) and immunohistochemistry on paraffin-embedded tissue (Giordano et al. 1996; Cinti 1999) have shown that WAT is mainly provided with noradrenergic nerves, distributed as perivascular fibers mainly in the adventitia of arteries and as sparse parenchymal fibers in close association with capillaries. To determine whether fasting-dependent delipidation was associated with a rearrangement of the nerve supply in retroperitoneal WAT, we initially performed Western blotting analyses on control and fasted rats by detecting PGP 9.5. Results showed a significant and progressive increase in its expression (Figure 3A), suggesting that a progressive increase in nerve density takes place during prolonged fasting. This increase was matched by an increased expression of TH, the marker of noradrenergic nerves (Figure 3B). In particular, TH increased significantly at 15% of weight loss, remained unchanged at 20% and 25% of weight loss, and then increased again at 30% of weight loss. By contrast, PGP 9.5 increased progressively and almost linearly during fasting. These different patterns of the structural (PGP 9.5) and the functional (TH) nerve marker might be explained by a process whereby peripheral nerves first grow, prompted by adequate stimuli, and later acquire the neurotransmitter phenotype.
Indeed, by immunohistochemistry the retroperitoneal depot showed a high density of noradrenergic nerves. Rich plexuses of noradrenergic nerves were found around large arteries and arterioles (Figure 4A), and numerous TH-positive nerves usually associated with capillaries were found among white adipocytes (Figures 4C and 4E). The distribution of the parenchymal noradrenergic nerves was denser in some WAT areas and sparser in others. Nevertheless, light microscopic analysis did not permit the inference of a clear connection between density of innervation and degree of delipidation. Numerous NPY-containing nerves were seen around arteries and, to a lesser extent, in the parenchyma, where they were always associated with capillaries (Figure 5, bottom panel). Immunofluorescence and confocal microscopy showed that the vast majority of vascular, and some parenchymal, noradrenergic nerves also contained NPY (Figure 5). Some “pure” NPY-containing nerves were found around arteries and, rarely, in the parenchyma.
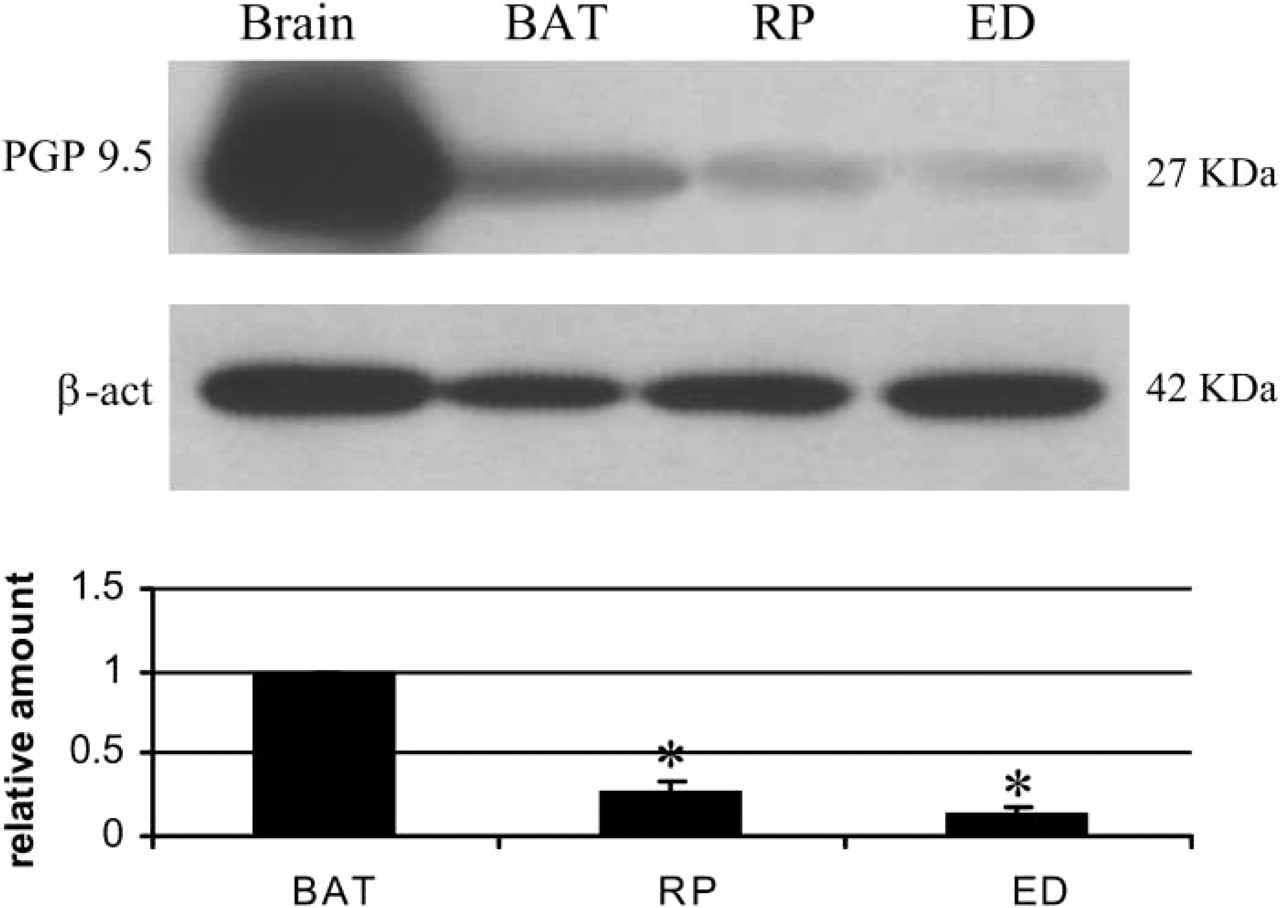
Western blot protein gene product (PGP) 9.5 analysis of brain (positive control, 25 μg), interscapular brown adipose tissue (BAT) (60 μg), retroperitoneal (RP, 60 μg), and epididymal (ED, 60 μg) fat in normal adult rats. PGP 9.5 quantities are normalized with β-actin; RP and ED signals are referred to BAT taken as 1.0. Values are expressed as mean ± SE (
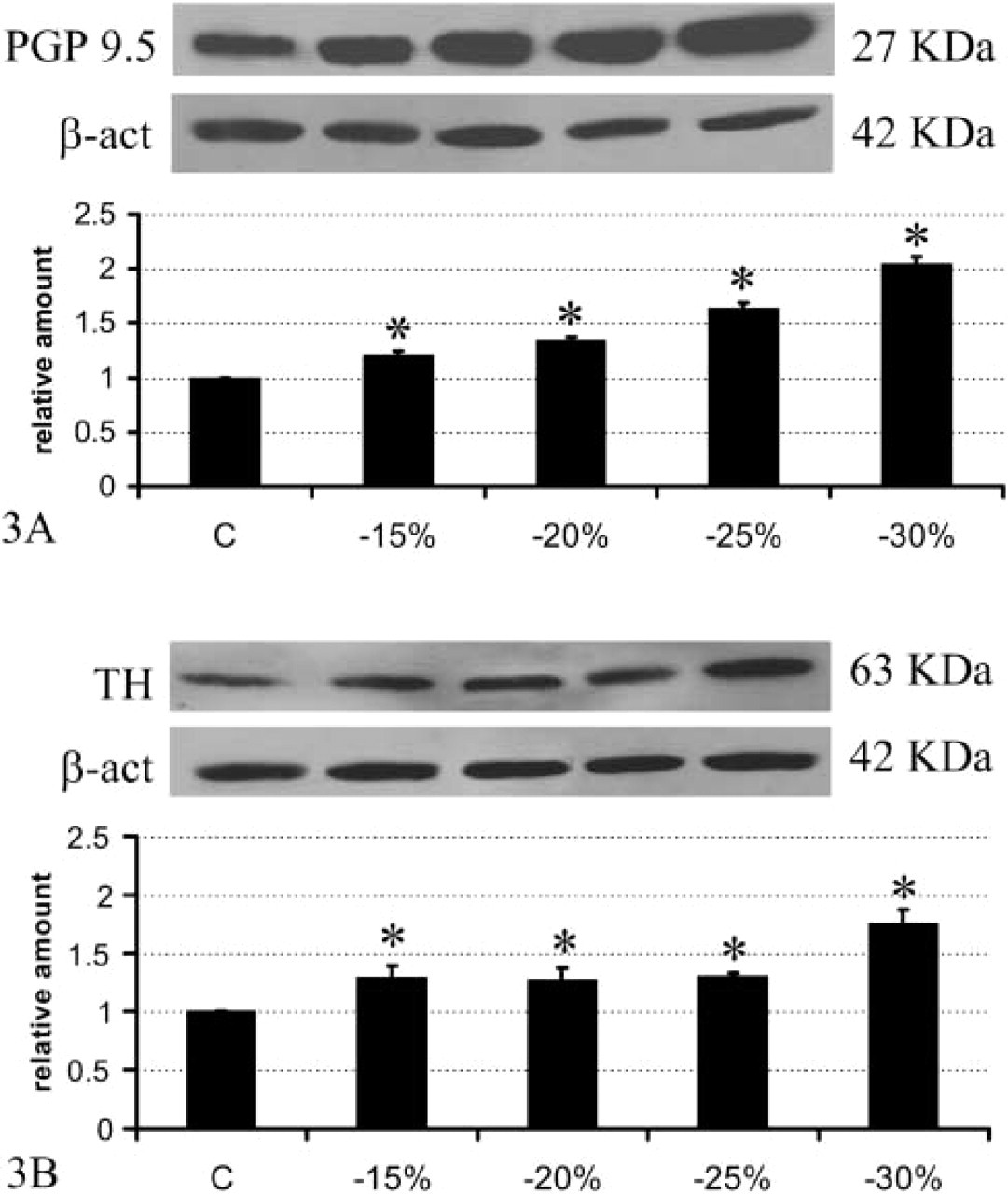
Western blot PGP 9.5 (
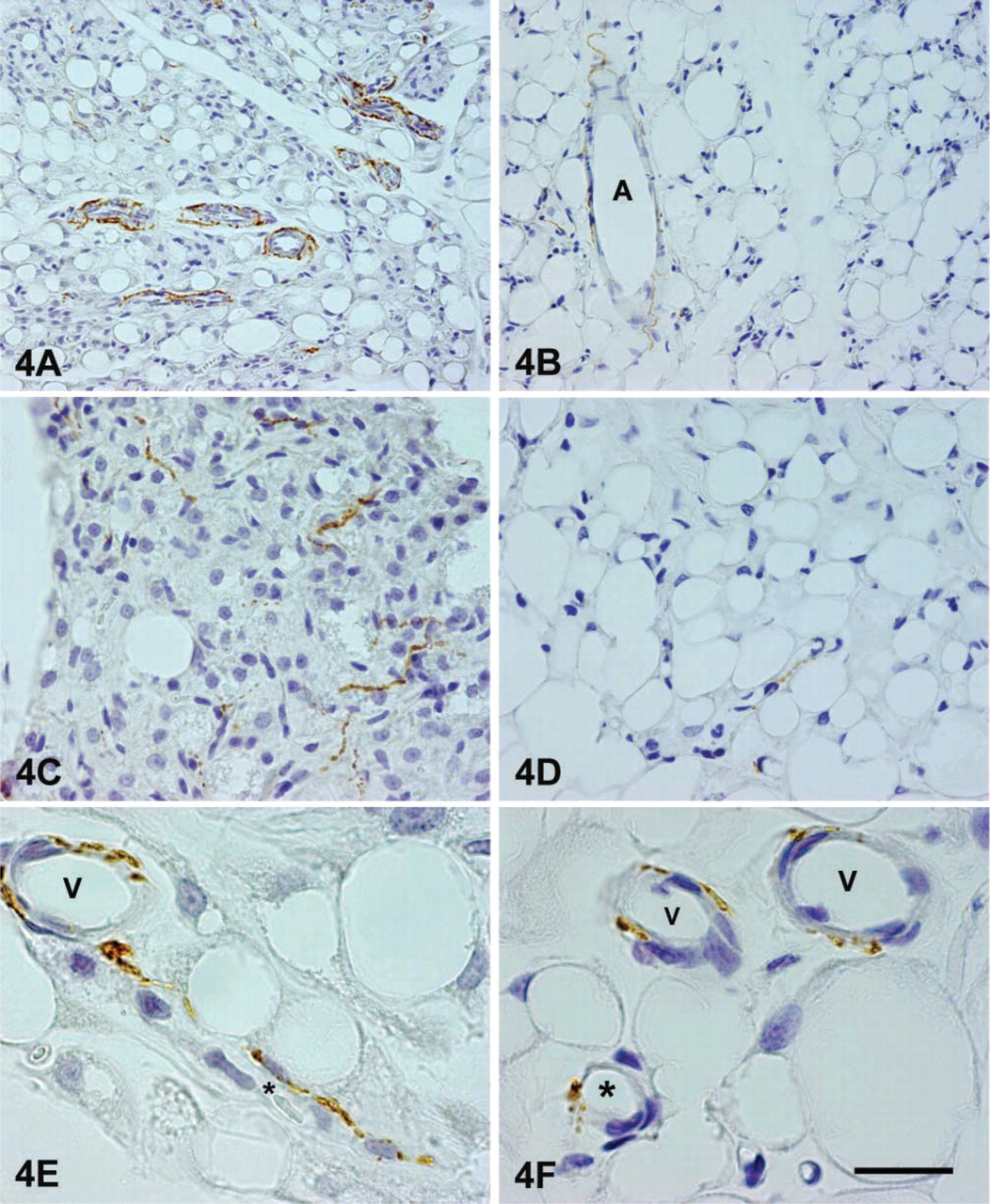
Sympathetic noradrenergic nerve distribution in retroperitoneal (
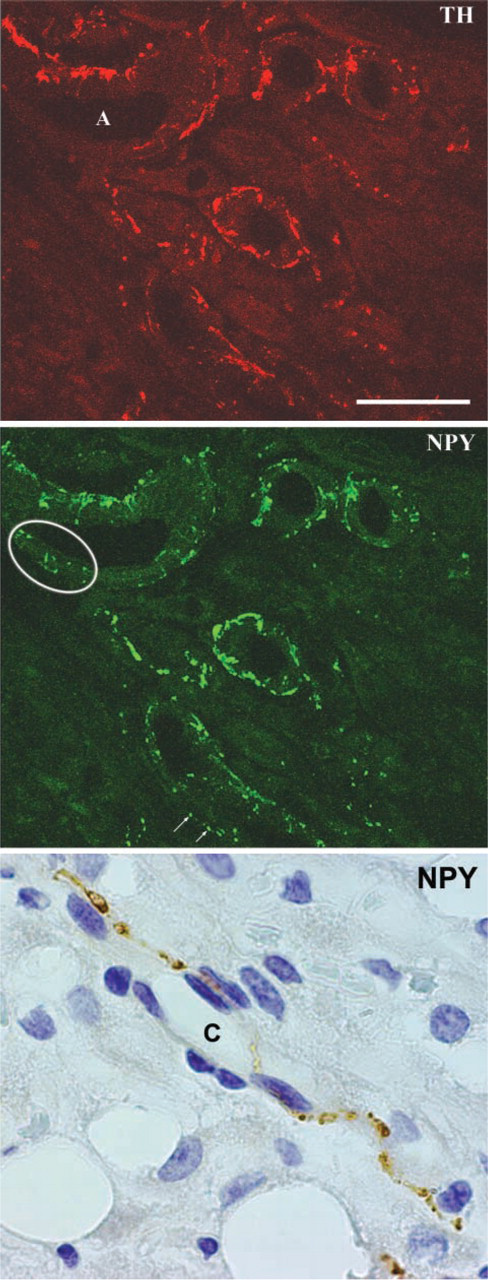
Neuropeptide Y (NPY)-containing nerves in the retroperitoneal depot of rats that have lost 25% of body weight. At confocal microscopy the vast majority of perivascular noradrenergic TH nerves (upper panel) are also seen to contain NPY (middle panel). Some “pure” perivascular peptidergic nerves are also visible (circled area and white arrows). Bottom panel: a parenchymal NPY-containing nerve is found in close association with a capillary (C). Bar: Upper and middle panels = 50 μm; bottom panel = 35 μm.
To establish whether the increased nerve density observed in retroperitoneal WAT depended solely on an increase of noradrenergic nerves or also of other nerve types, fasted retroperitoneal WAT was double stained with PGP 9.5 and TH. Observation of numerous sections in all fasted conditions revealed very few nerves to be not noradrenergic, allowing to conclude that, during fasting, nerve density in retroperitoneal WAT increases and that this increase is entirely due to sympathetic noradrenergic innervation.
Epididymal Depot
Compared with the retroperitoneal depot, the fasted epididymal WAT underwent a smaller and slower reduction in weight (Figure 1A) and volume (Figure 1B). At light microscopy, the delipidation process did not show as many evident clusters of shrinking adipocytes, and in animals that had lost 30% of body weight most adipocytes were devoid of lipid store and only few adipocytes still contained small lipid droplets.
The epididymal WAT was ∼6.8 times less innervated than BAT and, importantly, its content in PGP 9.5 was about half that observed in retroperitoneal WAT (Figure 2). The two depots thus exhibit different degrees of innervation, although the functional significance of such differences is still unclear. By Western blot analysis the epididymal WAT, unlike the retroperitoneal depot, did not exhibit an increase of PGP 9.5 (Figure 6A), whereas TH increased slightly (by ∼15%) but significantly in rats with a weight loss of 15%, 20%, and 25%, and rather more (by ∼30%) in those that had lost 30% of body weight. Accordingly, immunohistochemistry showed a moderate degree of noradrenergic innervation, mainly around parenchymal arteries (Figure 4B), and rarely in the parenchyma itself, that was always associated with capillaries (Figures 4D and 4F). As in the retroperitoneal depot, the vast majority of perivascular noradrenergic nerves also contained NPY, some “pure” NPY-containing nerves were found around arteries, and very few nerves supplying the epididymal depot at the vascular or the parenchymal level were not noradrenergic (Figure 7, bottom panel).
Western blot PGP 9.5 (
Discussion
Fasting is associated with reduced noradrenaline turnover in heart (Young and Landsberg, 1977), pancreas and liver (Young and Landsberg 1979), brown fat (Young et al. 1982), and kidney (Daly et al. 1992). In contrast to the widespread suppression of centrally mediated sympathetic activity, WAT noradrenaline turnover increases in the fasted state (Migliorini et al. 1997). Here we show that TH, the catecholamine-synthesizing enzyme whose expression is related to rate of noradrenaline synthesis and content in sympathetic nerves (Flatmark 2000), increased significantly in fasted retroperitoneal and epididymal WAT. These results lend further support to the hypothesis that the sympathetic drive to WAT selectively increases during fasting. As shown by immunohistochemical analysis, such an increase involves both parenchymal nerves, from which the spillover of noradrenaline likely causes adipocyte lipolysis via adrenergic receptors (Lafontan and Berlan 1993) and blood vessel nerves.
In fasted rats, WAT blood vessels show rich plexuses of noradrenergic nerves as well as NPY-containing nerves. Using confocal microscopy, we found that the vast majority of the latter nerves were also positive for TH. Thus, NPY is co-stored with noradrenaline in WAT vascular nerves as in BAT (Cannon et al. 1986). The increased content of both neurotransmitters at the vascular level is likely related to the increased rate of WAT perfusion observed during fasting (Crandall et al. 1997), which allows for removal of the free fatty acids secreted by the slimming adipocytes. Notably, the presence of parenchymal NPY-containing nerves in the retroperitoneal and epididymal fat lobules of fasted rats might possibly underpin an action of NPY on the slimming adipocytes.
Unexpectedly, during fasting the WAT nerves showed different behaviors in the two depots. Evaluation of PGP 9.5 on Western blots showed unchanged nerve density and increased noradrenaline content in epididymal WAT, whereas in the retroperitoneal depot the more pronounced increase in noradrenaline content was accompanied by a significant and progressive increase in nerve density. Thus, while in the epididymal depot the increased sympathetic activity is linked to over-release of neurotransmitter from existing nerves, in the retroperitoneal depot such an increase is also due to de novo nerve formation. The linkage between sympathetic drive and WAT lipid mobilization thus appears to be closer in the retroperitoneal than the epididymal depot, where other hormonal or paracrine factors may act and give rise to the slower, more gradual, and less focal delipidation process observed in this depot, at least in these conditions.
It is unclear why the retroperitoneal and epididymal nerves exhibit such different behaviors in fasted animals. Different rates of fasting-dependent noradrenaline turnover (Youngstrom and Bartness 1995) and lipolysis (Sztalryd and Kraemer 1994) in different adipose depots have already been reported. The molecular cues regulating nerve rearrangement and eventual neoformation in fasted WAT are at present unknown. Nonetheless, it should be noted that white adipocytes have recently been shown to produce and secrete nerve growth factor (Peeraully et al. 2004). Furthermore, expression of Sema3A, a neurorepellent factor, is reduced in smooth muscle cells of WAT arteries and white adipocytes of fasted rats (Giordano et al. 2003), suggesting that a rearrangement of vascular and parenchymal nerves occurs in this condition.
Double staining of slimmed WAT sections with TH and the pan-nerve marker PGP 9.5 evidenced very few non-noradrenergic nerves at the vascular and the parenchymal levels. Although we could not assess their phenotype, substance P- and calcitonin gene-related peptide-containing nerves have been described in WAT pads (Giordano et al. 1996,1998). Moreover, the presence of parasympathetic nerves in WAT has recently been hypothesized (Kreier et al. 2002). Our data showing rare non-adrenergic nerves in fasted WAT argue against the presence of a substantial nerve supply other than sympathetic innervation.
Finally, the concepts regarding the physiological role of adipose tissue have changed profoundly over the past decade, with the recognition that WAT is not only a passive store of lipids but also an active endocrine organ secreting a wide range of hormones under the control, in most cases, of the sympathetic tone (Rayner 2001). Thus the fine, regional-dependent nerve adjustment occurring in WAT during fasting might account not only for white adipocyte fatty acid mobilization but also for the modulation of hormone secretion required to adapt body metabolism, energy expenditure processes and animal behavior to a resource-poor environment.
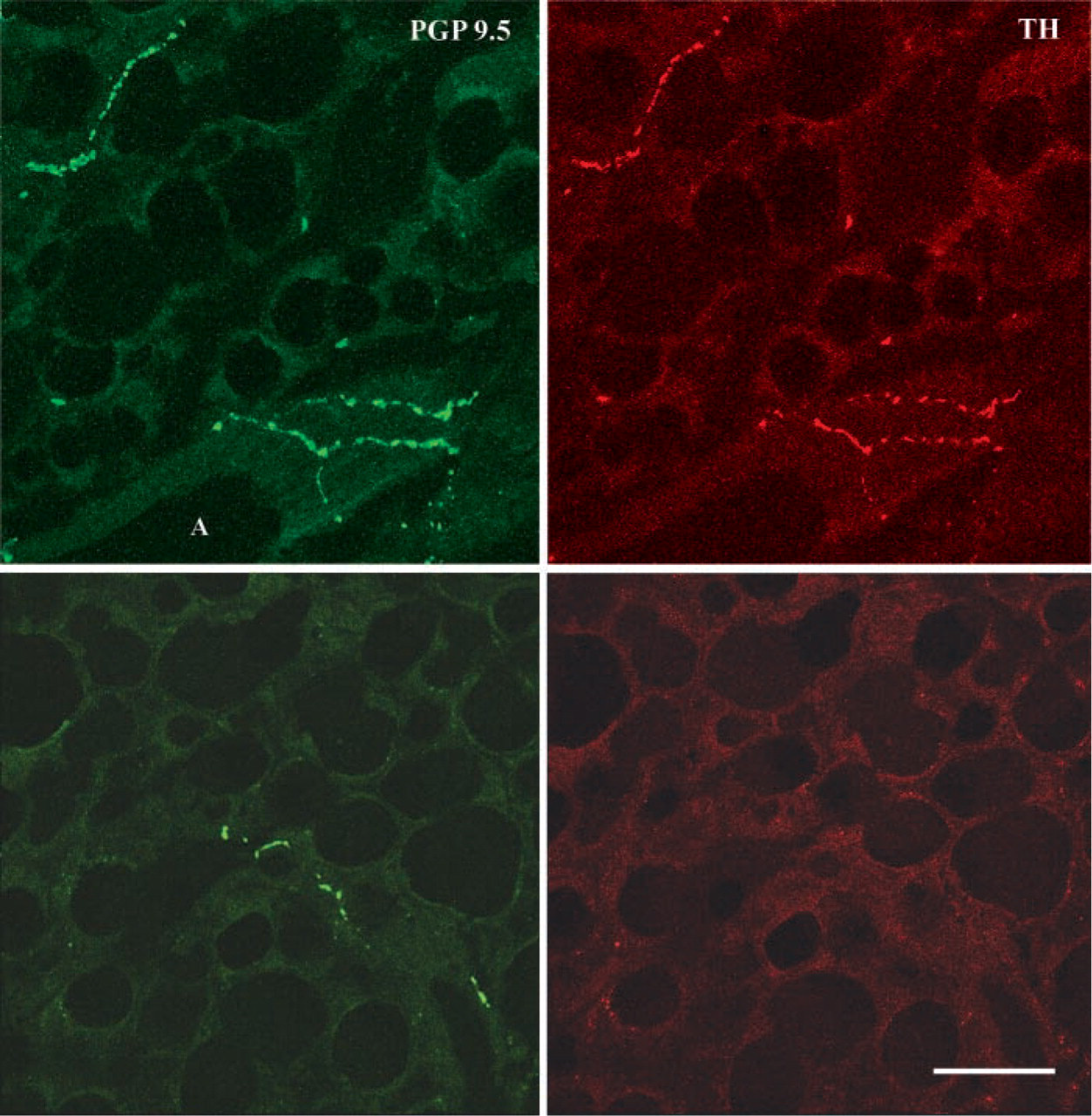
Double immunostaining for PGP 9.5 and TH and confocal microscopic analysis of epididymal depot after a weight loss of 25%. The vast majority of vascular and parenchymal nerves (structures positive for PGP 9.5, left upper panel) are also positive for TH (noradrenergic nerves, right upper panel). Bottom panels: a non-noradrenergic parenchymal nerve. A, artery. Bar = 40 μm.
Footnotes
Acknowledgements
This work was financed by grants from Polytechnic University of Marche (Contributi Ricerca Scientifica), by Ministry of University Cofinanziamento Bando 2002 and FIRB 2001 to S.C. and Cofinanziamento Bando 2003 to E.N., and by Ministry of Health grants to E.N. and M.O.C.